
生物有机肥对马铃薯产量与土壤氮循环作用机制研究
2022-05-12 09:31赵健宇王凤新孟潮彪
农业机械学报 2022年4期
赵健宇 王凤新 孟潮彪,2 金 松 彭 敏
(1.中国农业大学水利与土木工程学院,北京 100083;2.浙江大学土水资源与环境研究所,杭州 310058;3.怀俄明大学工业与民用建筑工程系,拉勒米 WY 82071;4.元泰丰(包头)生物科技有限公司,包头 014199)
0 引言
土壤氮循环是土壤生态系统中重要的循环之一,与其密切相关的氨氧化细菌(Ammonia-oxidizing bacteria,AOB)、氨氧化古菌(Ammonia-oxidizing archaea,AOA)、反硝化细菌和固氮菌是参与硝化、反硝化和固氮作用的重要功能微生物,其群落丰度和结构等因素直接调节氮素在土壤中的转化,影响作物对氮素的吸收利用[1]。
研究发现,施肥会导致土壤微生物的丰度和结构发生变化。有机和无机肥料配合施用已被证明是提高土壤肥力和维持农作物产量的有效施肥方式[2]。施用粪肥、作物秸秆作为有机肥料还田能显著增加固氮菌、反硝化菌和AOB丰度[3-6]。然而,也有研究发现施用粪肥对反硝化菌丰度无影响,无机肥与粪肥配施减少固氮菌丰度,而无机肥与秸秆还田配合对固氮菌丰度无明显影响[4,7-9]。有机肥料对氮循环相关微生物的丰度产生不同程度的影响,与土壤理化特性的变化有关。有机肥料的施入直接为反硝化菌等异养微生物生长提供碳源,促进土壤微生物生长[10]。不同有机肥和无机肥配施量对氮素形态(铵态氮和硝态氮)的影响直接关系到硝化作用和反硝化作用底物的浓度,进而对相关微生物的丰度和功能产生影响[11-12]。当土壤中可利用的氮能充分满足固氮微生物的需求时,固氮微生物的活性将会降低,生物固氮作用受到抑制[13]。此外,施肥造成的土壤pH值变化也是影响微生物变化的主要因素之一[3,14-15]。因此,氮循环过程相互关联,受到土壤环境因子驱动,不同的肥料类型及施用量会对氮循环相关微生物的活性和丰度产生重要影响,从而影响土壤养分循环。
褐煤具有多孔性和较大的比表面积,具有较高的有机质和腐殖酸含量,对各种土壤化合物的吸附作用较强[16],有助于增加土壤有机碳含量、碳氮比和阳离子交换能力,是有机肥料的基础[17]。褐煤中的腐殖酸可以提高马铃薯对含氮化合物的吸收,促进马铃薯生长和块茎形成。与粪肥等有机肥料相比,褐煤稳定性较强,有机质矿化速率有限,短期内可能对土壤微生物群落丰度和结构影响较小[18]。土壤养分的供应离不开微生物的参与。随着生物技术的发展,出现以褐煤为原料进行微生物发酵生产的煤基生物有机肥[19-20]。煤基生物有机肥与无机肥料混施,能否促进肥料本身和土壤养分的缓慢释放,从而提高肥料利用效率;煤基生物有机肥能否改善土壤的理化特性和调节土壤氮循环功能微生物,进而促进马铃薯对氮素的吸收利用,实现产量提升,值得研究。
目前的研究很少关注生物有机肥联合化肥施用对作物生长及土壤氮循环微生物的影响。本研究拟通过大田试验,利用分子微生物学方法标记相关功能基因,研究增施煤基生物有机肥对滴灌马铃薯产量和土壤理化特性以及氮循环相关功能微生物丰度的影响,揭示煤基生物有机肥对作物产量和土壤氮循环的作用机制,为土壤科学管理和提高氮素利用率及制定合理施肥制度提供理论依据。
1 材料与方法
1.1 试验区概况
试验于2019年5月11日—9月10日在内蒙古自治区包头市固阳县旧城村(41°1′N,110°3′E,海拔1 356 m),该地区属于温带大陆性气候,年平均气温为4℃,年平均降雨量为300 mm,多年平均20 cm蒸发皿蒸发量为2 270 mm左右,年日照总时数为3 130 h,无霜期为95~110 d。该试验地土壤质地为砂壤土,0~80 cm处土层平均干容重为1.35 g/cm3,平均田间持水率约为21.67%(体积含水率)。前茬作物为马铃薯,耕作层(0~40 cm)土壤pH值8.10,有机碳质量比3.71 g/kg,全氮质量比0.39 g/kg,全磷质量比0.61 g/kg,硝态氮质量比12.0 mg/kg,速效钾质量比141.0 mg/kg。
1.2 试验设计
试验共计4个处理:单施化肥(CF)、化肥增施1 500 kg/hm2煤基生物有机肥(BF1)、化肥增施3 000 kg/hm2煤基生物有机肥(BF2)、化肥增施4 500 kg/hm2煤基生物有机肥(BF3),每个处理重复3次,共计12个小区,随机布置。所用化肥为尿素(含N 46%)、磷酸二铵(含N 16%,P2O544%)、硫酸钾(含K2O 50%),施用量为210、150、180 kg/hm2。煤基生物有机肥由元泰丰(包头)生物科技有限公司生产,主要原料为褐煤,其pH值为8.48,有机质质量比为976 g/kg,全氮质量比为30.9 g/kg,全碳质量比为366 g/kg,碳氮比为11.9。
1.3 农艺措施
试验所用马铃薯品种为克新一号,每个小区种植7垄,垄长6.00 m,垄底宽0.80 m,垄高0.30 m,一垄单行,株距约为0.25 m,播种深度为0.15 m,种植密度为52 500 株/hm2,所有处理小区均采用黑色地膜(宽约0.70 m,厚约0.008 mm)覆盖。马铃薯于2019年5月11日播种,9月10日收获。采用滴灌灌溉,垄的中央、滴头的正下方埋设负压计,陶土头埋深为20 cm,土壤水势低于-25 kPa时开始灌水,每次灌水量取5 mm[21]。马铃薯全生育期共灌水56次,灌水总量为280 mm,各生育期(除A外)灌水量如图1(图中A~E分别表示芽条生长期、幼苗期、块茎形成期、块茎膨大期、成熟期)所示。有机肥和全部磷肥、30%氮肥和30%钾肥在旋耕起垄前人工撒施,剩余氮肥和钾肥平均分成2份分别在块茎形成期(6月24日)和块茎膨大期(7月20日)随水追施。

图1 全生育期累计灌水量Fig.1 Accumulated irrigation amount in growth period
1.4 测定项目及方法
1.4.1作物干物质量及块茎产量
干物质量:块茎成熟期从每个小区随机选取10株马铃薯,将植株茎、叶、根和块茎分离并称鲜质量,植株的茎、叶、根和块茎置于105℃干燥箱中杀青30 min,然后80℃恒温干燥至质量恒定,再冷却至室温(20℃),分别称量得茎、叶、根和块茎的干物质量。
块茎产量及分级:收获时每个小区随机取10株马铃薯,用电子天平(精度为0.01 g)称量单株块茎质量和商品薯质量(≥75 g);每个小区选取中间3垄称取鲜质量,计算单位面积上块茎总产量。
1.4.2土壤样品采集
块茎成熟收获前,各小区选取3个代表样点采集0~20 cm深度土样,混合均匀后分成2份装入无菌袋低温保存带回实验室。一份鲜土4℃保存,用于测定土壤理化性质;另一份鲜土-20℃保存,用于提取土壤DNA。
1.4.3土壤理化性质测定
土壤理化性质的测定参照《土壤农业化学分析方法》[22]中的相关方法进行。土壤pH值用土壤pH计测定,水土质量比为5∶1;土壤含水率采用干燥法测定;有机碳含量采用重铬酸钾容量法测定;全氮、全磷和全钾含量分别采用凯氏定氮法、氢氧化钠熔融-钼锑抗比色法和氢氧化钠熔融-火焰光度法测定;2 mol/L氯化钾溶液浸提土壤后用连续流动分析仪测定硝态氮和铵态氮含量。
1.4.4土壤微生物DNA提取和功能基因荧光定量分析
从土样中取0.5 g,通过PowerSoil DNA提取试剂盒(MoBio Laboratories,Carlsbad,CA,美国)按操作步骤提取土壤总DNA。提取的DNA保存在-20℃下备用。通过琼脂糖凝胶电泳((Bio-Rad,Hercules,CA,美国)和ND-2000型分光光度计(Thermo Fisher Scientific,Waltham,MA,美国)检测所提DNA片段大小和质量。PCR反应体系为20 μL,其中包含4 μL的5×FastPfu缓冲液,2 μL dNTPs(2.5 mmol/L),上下游引物(5 μmol/L)各0.8 μL,0.4 μL的FastPfu聚合酶(Promega,Fitchburg,WI,美国)和10 ng的DNA模板,最后用双蒸水补齐至20 μL。16S rRNA、ITS、nirS、AOB-amoA和nifH基因荧光实时定量PCR分析所用的引物及反应条件见表 1。质粒及标准曲线的制备参照文献[23]。

表1 PCR扩增引物及反应条件Tab.1 Primers and reaction conditions of PCR analysis
1.5 数据分析
采用Microsoft Excel 2010对试验数据进行计算,Origin 2017进行绘图,SPSS 20统计软件进行统计分析,采用单因素方差分析和LSD法分析处理间显著性差异(P<0.05),相关性分析采用Pearson法。
2 结果与分析
2.1 煤基生物有机肥对马铃薯成熟期干物质量和块茎产量的影响
由图2(图中不同小写字母表示处理间差异显著(P<0.05),下同)可知,BF3处理马铃薯植株干物质积累总量最大,比CF处理多27.11%。在块茎成熟期时,干物质积累量主要集中于块茎,占干物质积累总量的62.21%~71.65%;其次是茎,占干物质积累总量的17.56%~25.76%;根和叶干物质积累量分别占总量的2.31%~2.98%、8.30%~9.73%。煤基生物有机肥对块茎干物质积累量具有极显著影响(P<0.01),BF3和BF2处理块茎干物质量分别比CF处理多10.37%、9.32%,BF1较CF处理多6.05%;BF2与BF3间无显著性差异(P>0.05)。

图2 马铃薯成熟期干物质积累量Fig.2 Dry matter accumulation at potato maturation stage
由图3可知,增施煤基生物有机肥处理的产量均高于单施化肥处理且差异显著(P<0.05),随着煤基生物有机肥量的增加,单株薯质量、商品薯质量和块茎总产量均有不同程度增加。与CF处理相比,BF3处理显著提高了单株薯质量、商品薯质量和块茎总产量,分别增加了22.62%、17.73%和9.49%。就块茎总产量来说,增施1 500~4 500 kg/hm2的煤基生物有机肥,较单施化肥增产幅度为5.30%~9.49%。
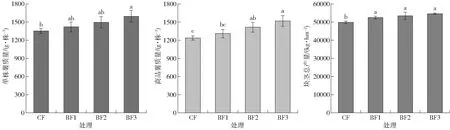
图3 不同处理对马铃薯单株薯质量、商品薯质量和块茎总产量的影响Fig.3 Tuber weight per plant,commodity potato weight and yield in responding to different treatments
2.2 煤基生物有机肥对土壤基本理化性质的影响
不同煤基生物有机肥增施量对0~20 cm土壤pH值和有机碳、全氮、全磷、全钾、硝态氮、铵态氮含量的影响,如表2所示。与CF处理相比,BF1、BF2和BF3显著降低了土壤pH值,显著增加了有机碳含量。增施煤基生物有机肥对土壤全氮、全钾、硝态氮和铵态氮含量变化没有产生显著差异,但随着增施量的增加,呈现出土壤全氮、硝态氮和铵态氮递增趋势。

表2 不同处理的土壤基本理化性质Tab.2 Physical and chemical properties of soils under different treatments
2.3 煤基生物有机肥对土壤微生物丰度的影响
与单施化肥相比,增施煤基生物有机肥可以使0~20 cm深度土壤细菌和真菌总量增加26%~97%,其中细菌丰度增加33%~104%,真菌丰度增加11%~80%(图4),但无显著差异(P>0.05)。而随着煤基生物有机肥增施量的增加,土壤细菌和真菌丰度均呈现减少趋势,但均高于CF处理。各处理中土壤细菌/真菌丰度比为2.4~3.3,随生物有机肥增施量的增加呈现出先降后升的趋势;BF3处理较BF2处理真菌丰度减少26%,细菌丰度几乎不变,致使BF3处理土壤细菌/真菌丰度比提高;而BF2处理比BF1处理真菌丰度减少17%,细菌丰度减少31%,造成BF2处理土壤细菌/真菌丰度比大为降低,甚至低于CF处理。说明适量增施煤基生物有机肥可以增加土壤细菌和真菌的丰度,进一步增施会导致其丰度减少;由于微生物丰度减少的比例不同,细菌/真菌丰度比发生变化。
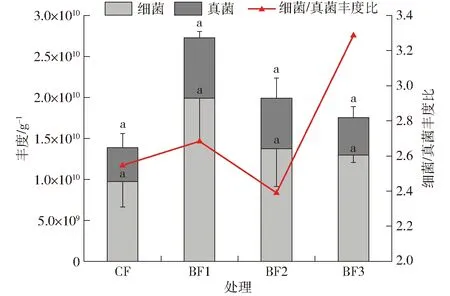
图4 不同处理土壤中细菌和真菌丰度及细菌/真菌丰度比Fig.4 Abundance and ratio of bacteria and fungi in soil under different treatments
2.4 煤基生物有机肥对土壤氮循环相关微生物功能基因丰度的影响
由亚硝酸盐还原酶基因(nirS)、氨氧化细菌(AOB)的氨单加氧酶基因(amoA)和固氮酶基因(nifH)丰度测定表明(图5),试验土壤中nifH基因和AOB-amoA基因拷贝数高出nirS基因拷贝数1个数量级,与CF处理相比,增施煤基生物有机肥的0~20 cm深度土壤中上述3种与氮循环相关的功能基因丰度均有所提高,但nifH基因和AOB-amoA基因在统计学上无显著差异(P>0.05)。与CF处理相比,对于nifH基因和AOB-amoA基因丰度,以BF3处理提高幅度最大,分别达74%和77%,BF2处理提高幅度最小,为29%和40%;对于nirS基因丰度,以BF2处理提高幅度最大,为90%,BF3处理提高幅度最小,为33%。随着煤基生物有机肥增施量的增加,nifH基因和AOB-amoA基因丰度呈现先减后增,nirS基因丰度呈现先增后减的趋势。

图5 不同处理土壤中nirS、AOB-amoA和nifH基因拷贝数Fig.5 Copies of nirS,AOB-amoA and nifH genes in soil under different treatments
2.5 土壤理化性质与氮循环相关微生物功能基因的相关分析
土壤微生物丰度与土壤基本理化性质的相关分析(表3)表明,细菌16S rRNA、真菌ITS和nifH基因丰度与土壤含水率、pH值、有机碳含量、全氮含量、硝态氮含量、铵态氮含量均无显著相关性。除nirS基因丰度与有机碳含量呈显著正相关,AOB-amoA基因丰度与全氮含量呈显著正相关外,nirS、AOB-amoA基因丰度与其他土壤理化性质无显著相关性。

表3 土壤微生物丰度与土壤基本理化性质的相关系数Tab.3 Correlation coefficients between soil microbial abundances and physical and chemical properties of soil
3 讨论
3.1 煤基生物有机肥对马铃薯干物质积累和产量的影响

3.2 煤基生物有机肥对土壤微生物丰度的影响
土壤微生物在一定程度上能反映土壤肥力水平,土壤细菌丰度高,表明土壤性质和水肥条件好,可以作为评价土壤健康的指标[35]。倪雷[36]研究发现,作物生育时期是影响土壤细菌丰度变化的首要因素,施加有机肥后增加了细菌丰度,而自小麦拔节期后丰度递减且无显著差异。本研究结果与之相似,增施煤基生物有机肥较单施化肥不同程度增加了土壤细菌和真菌丰度,但无显著差异。一方面由于施加有机肥带入了有机质和碳源,促进了土壤团粒结构形成,进而促进微生物生命活动和生长繁殖,细菌和真菌丰度增加[37];另一方面可能是作物生育期内气候变化和产生的根系分泌物导致成熟期各处理土壤细菌和真菌丰度无显著差异。随着煤基生物有机肥施用量的增加,土壤中细菌和真菌丰度均呈现减少趋势,这可能是由于煤基生物有机肥本身含有大量腐殖酸、大中小分子系列有机物和有益活性菌,这些组分均可加剧微生物活动,造成微生物间相互争夺养分,优势群体逐渐占主导地位,抑制其他种群繁殖,并充当关键物种(Keystone species),起到稳定微生物生态平衡的作用[38]。其次,褐煤在碱性土壤中可能释放出一些化合物,如酚类、邻苯二甲酸盐、多芳香烃、苯类和脂肪族化合物等,其中一些对细胞代谢有毒害作用[18]。这也是随着煤基生物有机肥施用量增加,微生物生长受到抑制的原因之一。
本研究中,增施4 500 kg/hm2和1 500 kg/hm2煤基生物有机肥土壤细菌/真菌丰度比高于单施化肥,而增施3 000 kg/hm2煤基生物有机肥土壤细菌/真菌丰度比低于单施化肥,说明土壤微生物区系可以通过施用生物有机肥进行调控,适宜的有机肥施用量对细菌/真菌丰度比有增加趋势。然而增施该种煤基生物有机肥对土壤微生物区系的调控规律仍需进一步研究。
3.3 煤基生物有机肥对土壤理化性质和氮循环功能基因的影响
增施煤基生物有机肥对土壤pH值和有机碳含量影响显著。增施煤基生物有机肥后土壤有机碳含量增加了4.8%~18.4%,这与肥料本身碳含量丰富密切相关;土壤pH值降低了0.10~0.17,这可能是由于在有机肥分解过程中有机酸和CO2释放到土壤中所致[39]。
本研究利用PCR技术定量检测了nirS、AOB-amoA和nifH3个关键氮循环功能基因,以研究增施煤基生物有机肥对参与反硝化作用、硝化作用和固氮作用微生物的影响。土壤pH值、有机碳含量、氮含量等理化性质是影响氮循环的重要环境因子。先前的研究表明pH值是决定土壤微生物丰度与群落的最主要因素[3,14,40],而本研究中土壤pH值与氮循环相关微生物丰度无显著相关关系,说明增施煤基生物有机肥虽然显著降低了土壤pH值,但未能改变土壤酸碱度对微生物丰度变化产生影响。
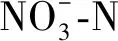
作为氨氧化作用的主要承担者,AOB在土壤氮循环中起到重要作用。先前有研究表明,肥料类型显著影响AOB丰度[45],而本研究结果表明,增施煤基生物有机肥对AOB-amoA基因拷贝数增加不显著。AOB是一类化能自养型微生物,一般不以有机碳为能源生长。因此,本研究中土壤有机碳含量增加没有导致AOB丰度显著提高。据报道,氨氧化作用的底物——氨的可利用性是氨氧化微生物生长的重要原因,无机氮作为能源物质,AOB的生长易受到其影响[12,40]。本研究中,AOB-amoA基因拷贝数与土壤全氮含量呈显著正相关,则进一步说明了氮素对AOB生长的重要性。
3.4 土壤氮循环特征与马铃薯增产机制

4 结论
(1)增施煤基生物有机肥可以调节土壤氮循环过程,增加土壤有机质和有效养分,显著提高马铃薯成熟期干物质积累量,显著提高单株薯质量、商品薯质量和块茎总产量,以增施4 500 kg/hm2煤基生物有机肥最佳。
(2)增施煤基生物有机肥较单施化肥不同程度增加了土壤细菌和真菌丰度,而随增施量增加,土壤细菌和真菌丰度均呈现出减少趋势,但无显著差异;适宜的生物有机肥增施量对细菌/真菌丰度比有增加趋势。
(3)增施煤基生物有机肥能显著降低土壤pH值,提高有机碳含量和nirS型反硝化菌丰度,而对AOB和固氮菌丰度影响不显著。相关性分析表明,nirS基因拷贝数与土壤有机碳含量显著正相关,AOB-amoA基因拷贝数与土壤全氮含量显著正相关,土壤有机碳是驱动nirS型反硝化菌丰度的重要环境因子。
猜你喜欢
安徽农业大学学报(2022年2期)2022-11-09
中国种业(2022年7期)2022-07-21
农家科技(2022年6期)2022-07-09
体育科技文献通报(2022年5期)2022-06-05
今日农业(2021年15期)2021-11-26
农民致富之友(2020年8期)2020-05-11
新农业(2019年4期)2019-05-29
大自然探索(2019年1期)2019-01-24
知识就是力量(2017年12期)2018-01-15
大自然探索(2017年3期)2017-04-07
