
改进的布谷鸟搜索算法对支持向量机参数优化
2022-05-11 01:32顾佳鑫贺兴时
西安工程大学学报 2022年2期
顾佳鑫,贺兴时, 刘 青
(西安工程大学 理学院, 陕西 西安 710048)
0 引 言
1995年,CORTES等学者基于VC维理论和结构风险最小原则(SRM),提出了支持向量机理论,将需要处理的分类问题转化为凸二次规划问题[1]。目前,已被广泛应用到智能交通[2]、金融预测[3]、生物医学和图像处理[4]等领域。
虽然SVM具有训练速度快和精确度高的优点,但SVM的性能受模型参数惩罚因子C和核函数的参数σ的影响[5]。因此,如何选择这2个参数是SVM研究的热点和难点。传统的SVM参数优化方法为网格搜索法、实验法、交叉验证法[6]和梯度下降法[7]。网格法和实验法耗时长;交叉验证法计算量大,难以实现2个以上的参数选取;梯度下降法对初始值选择敏感,实验结果误差大。因此,设计高效的参数寻优方法是目前的研究重点。
群智能算法在参数优化方面具有较强的并行处理能力,同时具有全局寻优的优点,故常用于SVM的参数优化[8]。具有代表性的为粒子群算法(PSO)[9]、人工鱼群算法(AFSA)[10]、人工蜂群算法(ABC)[11]、萤火虫算法(FA)[12]、蝙蝠算法(BA)[13]、花授粉算法(FPA)[14]和布谷鸟搜索算法(CS)[15]等。
针对原始布谷鸟搜索算法易陷入局部最优解、收敛速度慢、求解精度低的问题,宋庆庆等将混沌序列引入到布谷鸟搜索算法中来初始化鸟窝位置,增加布谷鸟种群多样性[16];陈程等提出了动态权重改变自适应概率的双重搜索布谷鸟算法(DECS),通过在寻优过程中引入的新型步长因子以及记忆策略,实现算法双重搜索模式的能力[17];张珍珍等将多阶段动态扰动引入到布谷鸟搜索算法中,并提出融合正弦余弦和种群初始化策略的布谷鸟算法[18-19]。该算法提高了求解精度,具有更好的优化性能。但上述文献都没有提到改进后布谷鸟搜索算法的应用问题。
为了提高原始布谷鸟搜索算法的寻优能力,本文提出改进的布谷鸟搜索算法(GFCS):用动态发现概率P代替固定发现概率Pa,自适应地调整布谷鸟莱维飞行的步长控制因子α,在布谷鸟随机游走更新公式中改进动态惯性权重w*;利用GFCS算法优化SVM的惩罚因子C和核函数参数σ解决SVM参数选择盲目的问题,并和传统的SVM、粒子群算法优化SVM、萤火虫算法优化SVM和布谷鸟搜索算法优化SVM进行比较。
1 支持向量机
支持向量机的原理是建立一个分类超平面作为决策面,使得正类和负类样本间隔最大化。将大小为l的训练样本集{(xi,yi),i=1,2,…,l}分为2类:如果xi∈Rn属于第1类,则标记(yi=1)为正;如果xi∈Rn属于第2类,则标记(yi=-1)为负。其中Rn为样本空间的维数[1]。可求得最优分类超平面:
(1)
式中:w为超平面法向量;b为阈值;C为惩罚因子;ξi为松弛变量;φ(xi)为映射函数。引入Lagrange函数,得到式(1)的对偶问题:
(2)
求解式(2)得到最优分类函数:
(3)
式中:ai(i=1,2,…,l)T为Lagrange乘子,K(xi,xj)=φ(xi)Tφ(xj)为核函数。本文采用径向基核函数作为SVM模型的核函数,表达式为
K(xi,xj)=exp(-‖xi-xj‖2/2σ2)
其中σ为核函数参数。
2 布谷鸟搜索算法及改进
2.1 原始布谷鸟搜索算法
布谷鸟搜索算法起源于布谷鸟孕育雏鸟的行为。一些布谷鸟在生产时会将自己的蛋放入百灵鸟、黄莺等宿主鸟的巢中,让它们代替自己喂养雏鸟。当宿主鸟发现鸟蛋来源不明,就会抛弃这些外来的鸟蛋或者重新筑巢。为了模拟布谷鸟的孕育雏鸟行为,YANG等提出了以下3条理想规则[20]:
1) 布谷鸟一次只产下一个蛋,并随机选择鸟窝来孵化;
2) 最好的鸟窝将被保留到下一代;
3) 可选择的鸟窝数量N是固定的,宿主鸟发现外来鸟蛋的概率Pa∈[0,1]。
基于以上3个理想状态,YANG等采用式(4)更新下一代鸟窝位置[20]:
i=1,2,…,N
(4)
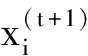
(5)
式中:α0为常数,一般取0.01;xb为当前最优解。
L(λ)~μ=t-λ(1<λ≤3)
(6)
由式(6)可知,布谷鸟在寻窝过程中,其飞行路径变化为带有重尾的概率分布,使得布谷鸟飞行路径表现出了莱维飞行的本质,即在寻优路径中频繁的短步长偶尔会出现长步长。采用式(7)计算莱维随机数[21]:
(7)
式中:μ和v服从标准正态分布;β=1.5,
结合式(4)~(7),CS算法通过莱维飞行采用式(8)更新鸟窝位置:
(8)
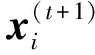
(9)
2.2 改进的布谷鸟搜索算法(GFCS)
2.2.1 动态调整发现概率
在原始布谷鸟算法中,用固定的发现概率Pa=0.25控制全局搜索和偏好随机游走,不利于全局搜索和局部搜索之间的平衡。本文提出动态发现概率P代替固定发现概率Pa提高算法寻优性能
(10)
式中:t为当前迭代的次数;T是最大迭代次数。P随t的变化如图1所示。
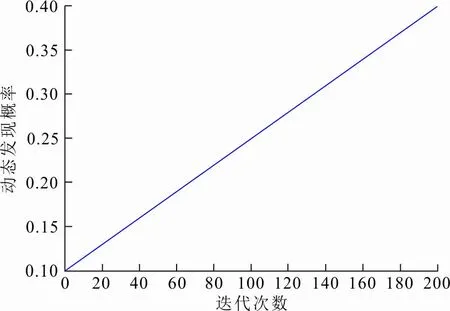
图 1 动态发现概率变化Fig.1 Change of dynamic discovery probability
由图1可以看出,动态发现概率P在[0.1,0.4]之间随迭代次数不断增大。迭代前期以较小的P进行全局搜索;随着迭代过程的进行,P不断增大,直至迭代后期;当P相对较大时,帮助算法在靠近全局最优解时进行局部精细搜索,提高算法的运算精度和搜索效率。
2.2.2 自适应调整步长
针对原始布谷鸟搜索算法寻优精度低的缺点,通过自适应地调整布谷鸟莱维飞行的步长因子α,使得莱维飞行步长随迭代的进行不断减小。在寻优初期拥有较大的步长因子,从而扩大算法前期的搜索空间,提高全局寻优能力;在寻优过程中,步长减小,提高算法局部搜索性能。基于上述分析,将式(7)的α0=0.01改成:
(11)
α0的变化如图2所示。由图2可以看出,α0的值随着迭代次数非线性递减。迭代前期α0的值较大,衰减速度较快,有利于提高算法全局寻优能力,保证算法前期收敛速度快;迭代后期α0的值逐渐减小,衰减速度放缓,提高算法的局部搜索精度。从而将式(8)更新为:
(12)
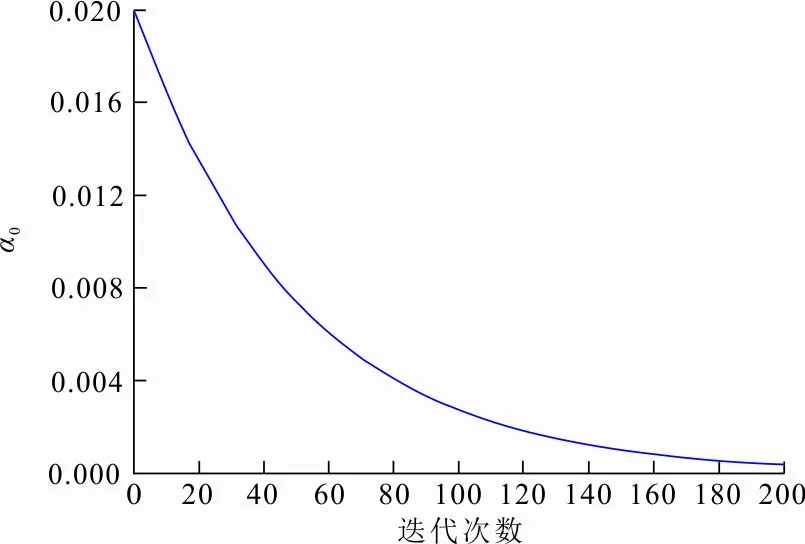
图 2 α0变化曲线Fig.2 Change of α0
2.2.3 动态惯性权重的偏好随机游动
在原始布谷鸟算法的偏好随机游动环节中,采用固定上一代鸟窝位置的更新方式,见式(9),容易造成算法在迭代后期陷入局部最优值。本文在上一代鸟窝位置处引入动态惯性权重w*,灵活地将上一代鸟窝位置与迭代过程动态联系起来。
(13)
式中:t为当前迭代的次数;T是最大迭代次数。w*的变化如图3所示。
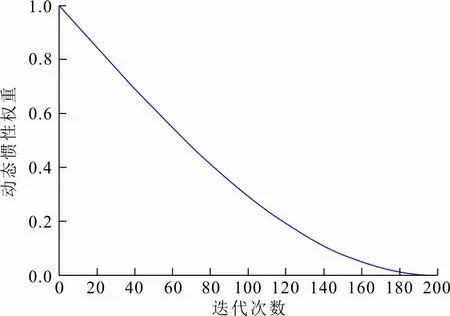
图 3 动态惯性权重w*变化Fig.3 Change of dynamic inertia weight w*
通过图3可以看出,w*在(0,1)之间随迭代次数非线性递减,即随着迭代次数增加,w*逐步减小。在迭代前期,给上一代鸟窝位置赋予相对大的w*,使上一代鸟窝具有更大的作用能力与范围,帮助布谷鸟算法在前期扩大搜索空间,广泛寻找全局最优解;到迭代后期,算法接近最优解时,对上一代鸟窝位置采用相对较小的w*,有效地削弱了上一代鸟窝的保留信息,帮助布谷鸟算法有效跳出局部最优,使布谷鸟个体具有更好的局部寻优能力。将式(9)更新为
(14)
3 GFCS算法对SVM参数优化
GFCS算法在优化SVM参数时,将待优化的参数组合(C,σ)模拟为鸟窝,具体步骤如下:
1) 原始数据集为训练集和测试集,并进行数据归一化。数据归一化公式为
(15)
式中:x、x*分别为归一化前后的样本值;xmax、xmin分别为样本数据中的最大、最小值。
2) 初始化参数。包括布谷鸟种群规模、布谷鸟鸟窝的位置、最大迭代次数、步长因子,被发现概率Pa、参数取值上下界及自变量(惩罚因子C和核函数的参数σ)等。
3) 随机分布鸟窝(C,σ)。采用式(16)作为布谷鸟鸟窝位置的适应度函数,并计算鸟窝的适应度值
(16)
式中:t为当前迭代的次数;a(t)为第t次迭代分类准确率。
4) 比较适应度值,保存当前最小适应度值及对应的鸟窝位置xi。
5) 根据改进后布谷鸟搜索式(12)、(14)更新鸟窝位置xi,并计算适应度值。
6) 产生一组随机数,与被发现概率Pa进行比较:若随机数小于被发现概率,则随机改变鸟窝位置;否则,保持原鸟窝位置不变。更新后的鸟窝位置有可能超出解空间范围,需要将其限制在解空间内:
(17)
式中:xmin和xmax分别为解空间的最大值和最小值i=1,2,…,n。
7) 比较适应度值,找到当前最优解并保留原最优鸟窝位置。
8) 若达到最大迭代次数或满足停止迭代条件,则转至9);否则转至2)继续迭代。
9) 输出最小适应度值并保留最优鸟窝,即得到最优参数组合(C,σ)。
10) 将输出的最优参数C,σ进行SVM模型的训练和测试。
4 实验结果及分析
实验在Matlab环境下实现,硬件配置为Windows 10操作系统,4 GiB内存,1 TiB硬盘,i7-7500和3.1 GHz主频CPU的计算机。
4.1 GFCS算法性能
为了分析GFCS算法的优化性能,选择6个常用的基准测试函数进行测试。其中Ackley、Rastrigin、Griewank为具有多个局部最小值的多峰函数,用于测试算法跳出局部极值,避免陷入局部最优的能力;Schwefel's2.2、Sphere、Sum square为单峰函数,用于测试算法的优化精度和收敛速度。所选择的6种函数设置见表1。
本次实验将提出的GFCS算法与PSO算法[22]、FA算法[23]、CS算法[20]进行比较。参数设置如表2,实验结果如图4所示。

表 1 基准测试函数Tab.1 Benchmark function

表 2 算法参数设置Tab.2 Algorithm parameter settings
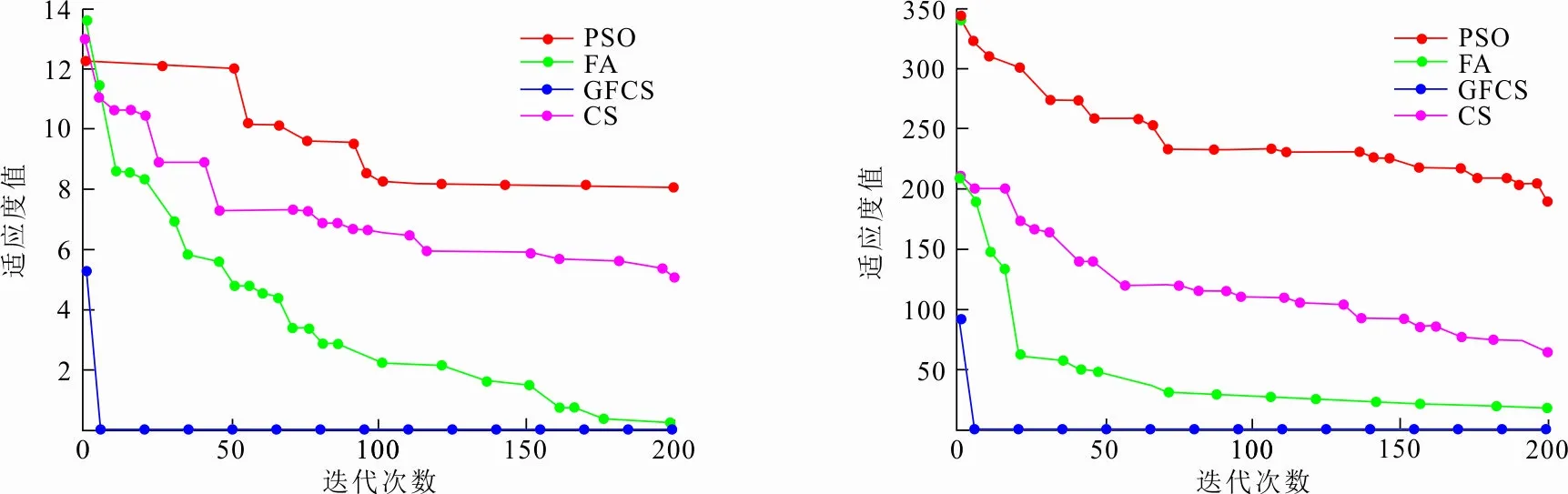
(a) Ackley函数收敛曲线 (b) Rastrigin函数收敛曲线
(c) Griewank函数收敛曲线 (d) Schwefel′s2.2函数收敛曲线

(e) Sum square函数收敛曲线 (f) Sphere函数收敛曲线图 4 不同算法在6种测试函数上收敛曲线Fig.4 Convergence curves of different algorithms on six test functions
4.2 GFCS算法优化SVM参数实验
为了验证GFCS算法对SVM的参数优化的有效性,选用径向基核函数作为实验核函数,优化对象为惩罚因子C和核函数的参数σ。采用UCI数据库中的标准数据集测试模型性能,实验数据集描述如表3所示。
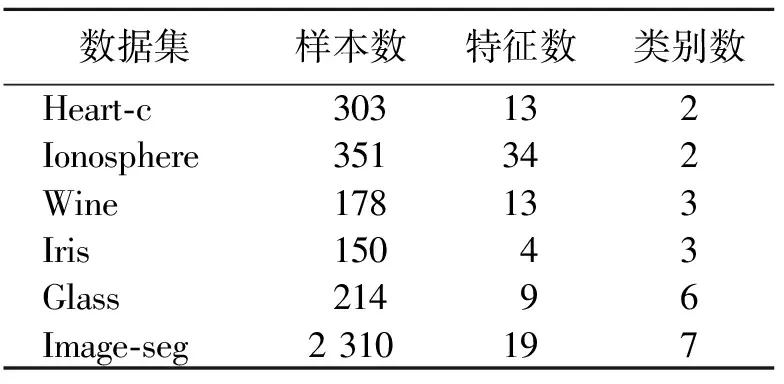
表 3 实验数据集描述Tab.3 Description of experimental data sets
从图4可以看出:GFCS算法收敛速度、稳定性和寻优能力明显高于PSO、FA、CS等算法,并且GFCS算法适应度值最低。表明GFCS算法全局寻优能力较强,不易陷入局部最优解。
实验中,选取未优化参数的SVM、基于粒子群算法的SVM(PSO-SVM) 、基于萤火虫算法的SVM(FA-SVM)和基于布谷鸟搜索算法的SVM(CS-SVM)作为对比实验,与GFCS-SVM相比较。
参数设置如下:SVM的惩罚因子C和核函数的参数σ上界为100,下界为0.01;PSO算法速度上界为5,下界为-5,学习因子γ1=γ2=1.5;FA算法步长因子k=0.5,光强吸收系数r=1,最大吸引度h=0.2;CS算法被发现概率Pa=0.25,步长因子a0=0.01;GFCS算法动态发现概率P∈[0.1,0.4],自适应步长因子a0∈[0,0.02],惯性权重w*∈[0,1]。PSO、FA、CS和GFCS算法的种群规模N=30,最大迭代次数T=200。
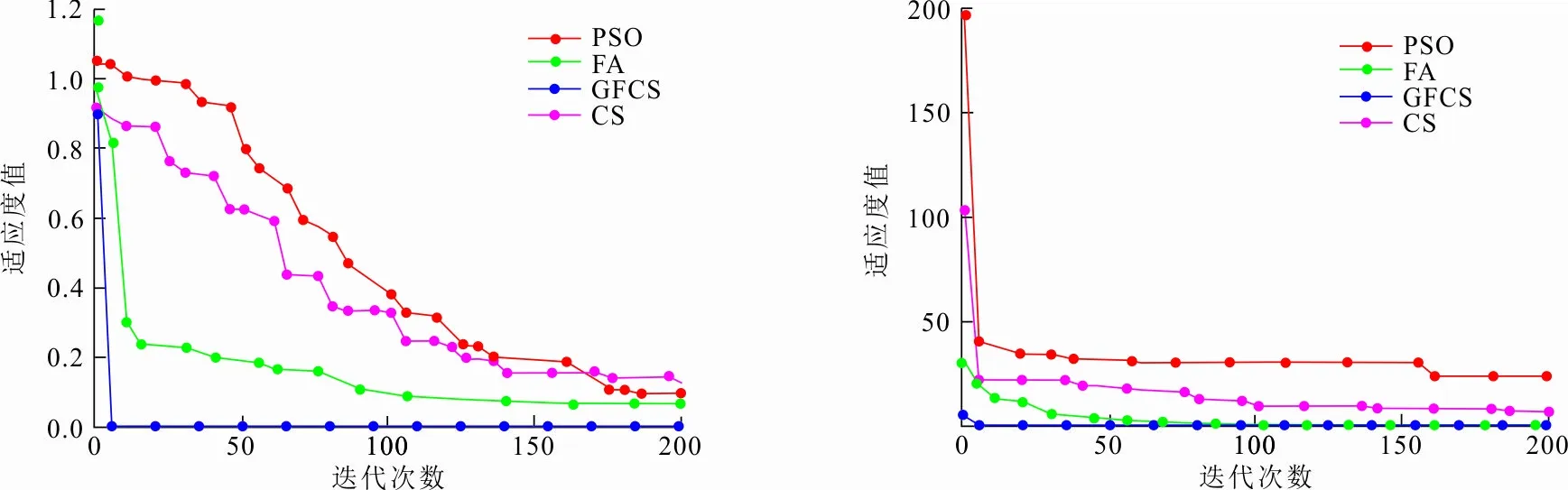
通过各个算法在6个UCI数据集上的实验仿真,分析GFCS-SVM分类性能。图5给出了GFCS算法优化SVM参数后在不同数据集分类效果,表4给出各算法在不同数据集上寻找的最优惩罚因子C和核函数的参数σ,表5给出各算法在不同数据集的分类准确率。结合图5和表5可以看出,GFCS-SVM在6个UCI数据集上都得到了最佳分类准确率。因此,和传统的SVM、原始群智能算法优化SVM相比,GFCS算法优化后SVM具有更好的分类效果。这是因为GFCS-SVM全局寻优和局部搜索最优解能力增强,可以更精确地锁定惩罚因子C和核函数的参数σ(见表4),使得SVM分类准确率提高。
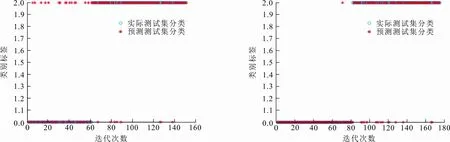
(a) Heart-c数据集 (b) Ionosphere数据集
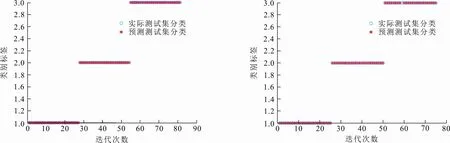
(c) Wine数据集 (d) Iris数据集
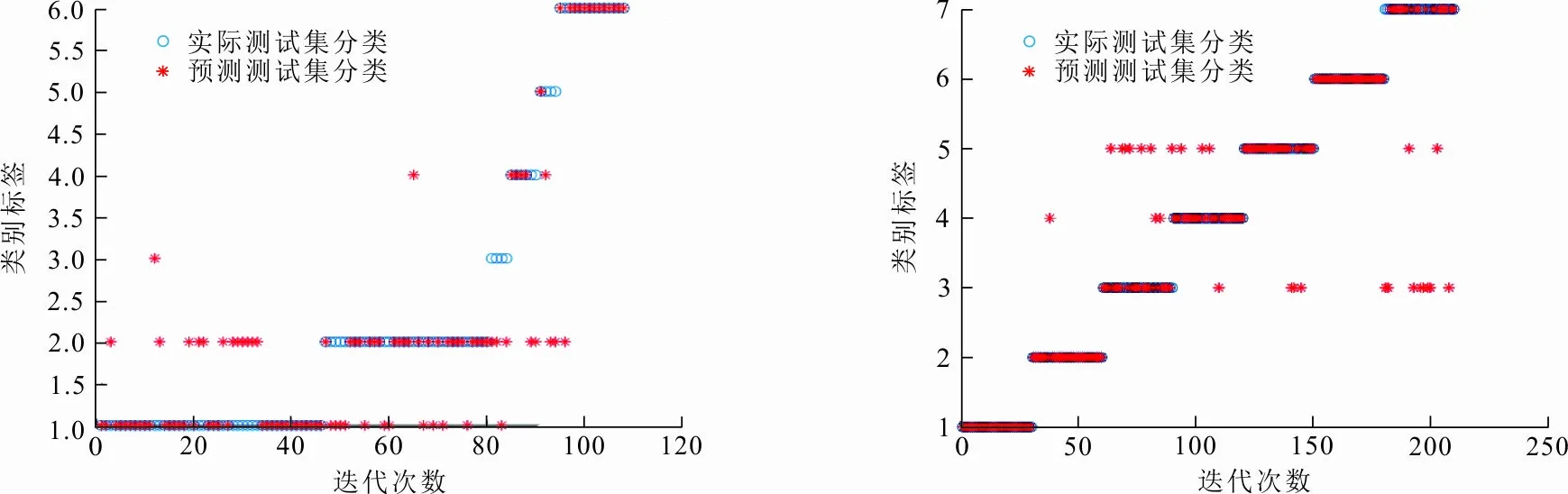
(e) Glass数据集 (f) Image-seg数据集图 5 GFCS-SVM在不同数据集的分类结果Fig.5 Classification result of GFCS-SVM on different data sets
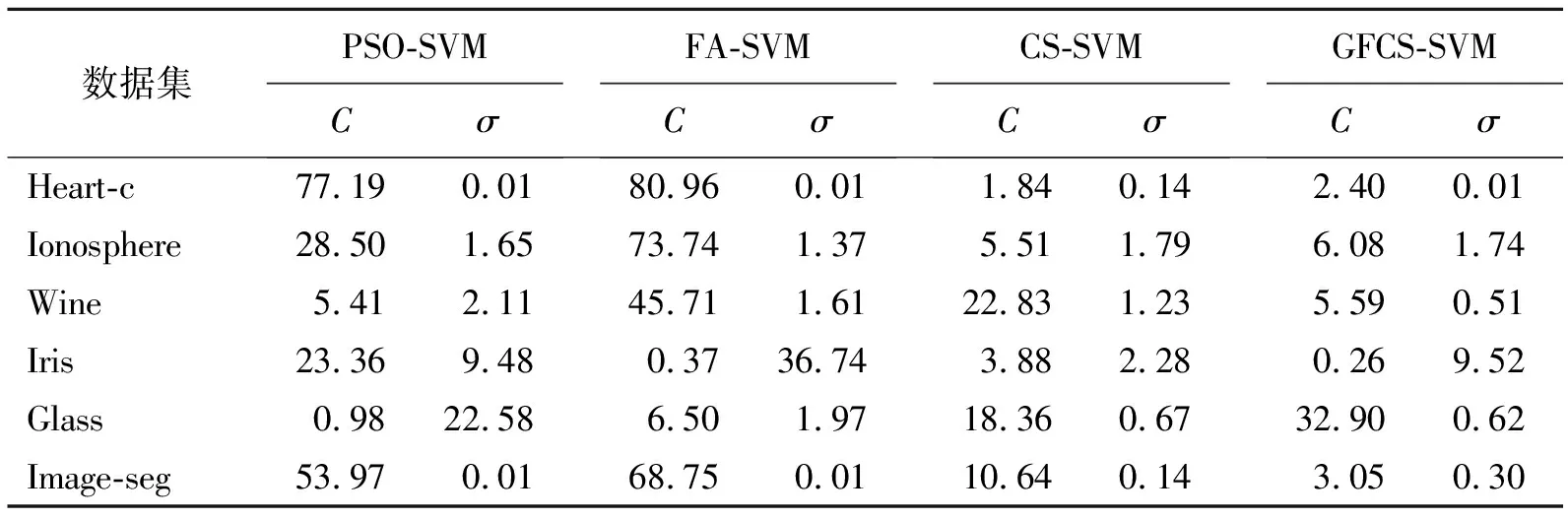
表 4 各算法在不同数据集上寻找的最优参数
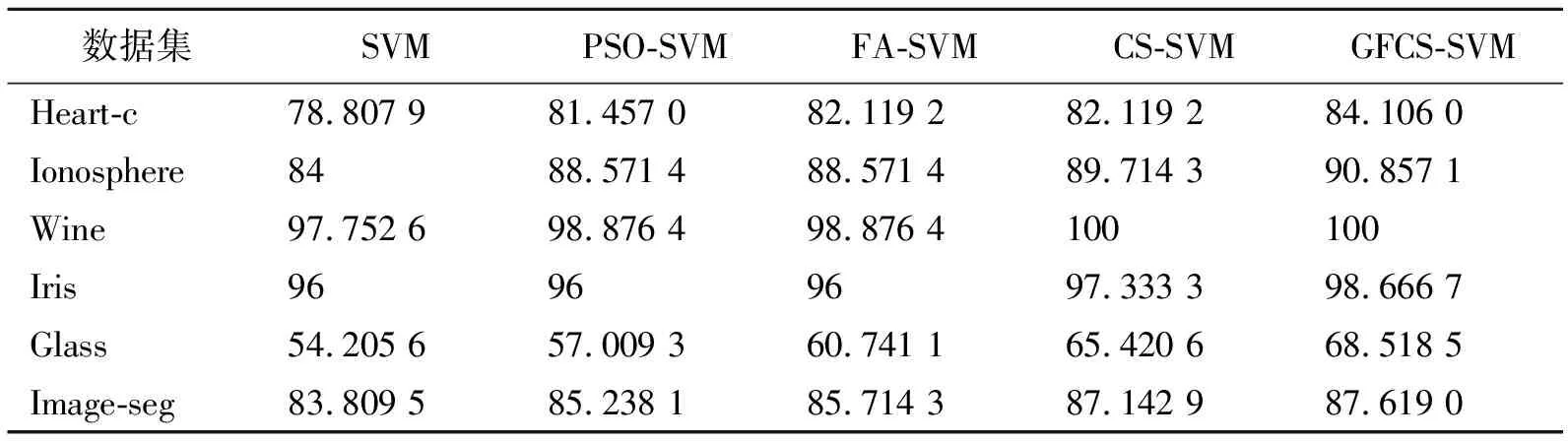
表 5 各算法在不同数据集上的分类准确率
5 结 语
针对原始布谷鸟搜索算法存在的易陷入局部最优、求解精度低,以及收敛速度慢等问题,从发现概率、步长因子和惯性权重等3个方面改进原始算法,提出了GFCS算法。通过6个基准测试函数的仿真,结果表明GFCS算法收敛速度快、有很强的稳定性和寻优能力。将GFCS算法应用于SVM惩罚因子C和核函数的参数σ寻优,避免了SVM参数选择盲目性;通过在6个UCI数据集进行测试,与未优化参数的SVM、PSO-SVM、FA-SVM、CS-SVM相比,GFCS-SVM分类效果最好。因此,GFCS算法是有效的SVM参数优化算法。未来可以将GFCS算法应用于其他SVM模型,如最小二乘支持向量机、孪生支持向量机和非平行超平面支持向量机等,解决大规模数据分类问题。
猜你喜欢
舰船科学技术(2022年11期)2022-07-15
红蜻蜓(2021年12期)2021-12-19
科学大观园(2019年10期)2019-09-10
证券市场周刊(2019年18期)2019-08-17
证券市场周刊(2019年19期)2019-05-27
中国经济周刊(2019年9期)2019-05-24
智能计算机与应用(2018年3期)2018-09-05
电脑知识与技术(2016年30期)2017-03-06
剑南文学(2016年14期)2016-08-22
