
小麦品种周麦22抗叶锈病的QTL定位
2022-05-05 05:24闫晓翠段振盈杨华丽姚占军李在峰
作物杂志 2022年2期
闫晓翠 段振盈 杨华丽 姚占军 李在峰
(1河北农业大学植物保护学院/河北省农作物病虫害生物防治工程技术研究中心,071001,河北保定;2河北农业大学农学院/华北作物种质资源研究与利用教育部重点实验室,071001,河北保定)
小麦叶锈病是由叶锈病菌(Puccinia tirticina)侵染引起的真菌病害,致病菌属于活体寄生菌,主要侵染小麦叶片,但在发病严重或高度感病的小麦品种中,也可侵染叶鞘和颖壳[1-2],该病害是影响世界小麦生产安全的主要病害之一。几乎每年小麦主产区均有叶锈病的发生,20世纪中后期墨西哥西北部发生叶锈病害大流行,产量损失高达70%[3],在中国华北冬麦区发生过6次(1969、1973、1975、1979、2012和2015年)叶锈病害流行,其中仅2015年产量损失就高达19.1万t[4-5]。近年来,小麦叶锈病发生危害有明显加重趋势。虽然采用化学药剂能短暂防治叶锈病,但培育广谱抗病品种是行之有效的最佳方法。
小麦对叶锈病抗性分为小种专化抗性(race specific resistance)和非小种专化抗性(non-race specific resistance)[6]。小种专化抗性通常是对特定菌种表现出抗性,常发生在小麦苗期,又称为苗期抗性,常因病原菌毒性的变异而丧失[7-8],通过基因聚合和基因布局可以延长此类抗性基因的有效性。非小种专化抗性一般对1种病原菌的多个不同生理小种或者多种病原菌有效,一般在成株期表现,通常具有慢锈抗病性(slow rusting resistance)和持久抗病性(durable resistance)的特点[9],多个抗性基因的聚合可能表现出对此类病菌完全抗病。
至今,已正式命名的抗叶锈病基因(Lr)有79个[10],其中大部分为苗期抗病基因,仅有16个抗叶锈病基因表现为成株抗性(Lr12、Lr13、Lr22a、Lr22b、Lr34、Lr35、Lr37、Lr46、Lr48、Lr49、Lr67、Lr68、Lr74、Lr75、Lr77和Lr78)。其中,Lr34/Yr18/Pm38/Sr57[11]、Lr46/Yr29/Pm38/Sr58[12]和Lr67/Yr46/Pm46/Sr55[13]等基因兼抗多种病害,具有一因多效及抗性持久稳定的优点。SSR标记具有多态性高、稳定性强、共显性高及成本低等优点,常被广泛应用于抗病基因的初步定位[14]。2016年,师令智等[15]利用SSR标记在CIMMYT品系19HRWSN-76中定位出1个苗期叶锈抗病基因LrHR76;2015年,王佳真等[16]在潍麦8号中定位到1个位于2AS染色体上的成株QTL位点QLr.hbau-2AS。
小麦品种周麦22(国审麦2007007)是河南省周口市农业科学院选育出的优良品种,该品种为半冬性品种,具有产量高、品质优、农艺性状优良及抗病性好等优点,在我国黄淮冬麦区南部得到了大面积推广和应用。至2019年周麦22累计种植面积达670万hm2,高产稳产的原因之一是对小麦叶锈病[17]和条锈病[18]均表现出良好的抗性。Wang等[17]对周麦22进行了苗期抗叶锈病研究,定位到1个苗期抗叶锈病基因LrZH22。为了明确其所携带的成株抗叶锈基因,本研究利用周麦22/铭贤169及其255个F2:3家系,对其进行成株期抗叶锈病QTL分析,并结合已有的研究结果进行比较,挖掘周麦22中抗小麦叶锈病的新QTL位点,并找到与QTL紧密连锁的分子标记以应用于辅助选择育种。
1 材料与方法
1.1 供试材料
抗病亲本为周麦22,系谱:周麦12(周8425A/SW73295)/温 6//周麦 13(周 8425B/周麦 9 号),感病亲本铭贤 169及其杂交、自交获得的 255个F2:3家系群体;感病对照品种为郑州5389,也作为叶锈菌的扩繁材料。于小麦成株期接种3个叶锈菌生理小种FHRT、THTT和THJT的混合菌种。小种的命名采用Long等[19]的四字母命名法。所有供试材料和叶锈菌种均由河北农业大学植物保护学院小麦锈病实验室提供。
1.2 成株期接种和侵染型调查
于2014-2015和2015-2016年度将抗病亲本周麦22、感病亲本铭贤169及其255个F2:3代群体分别播种于河北农业大学(河北保定)试验地。田间种植方式为行距25cm,行长1.5m,每行25粒,每9行加种1份感病品种郑州5389作为发病对照。适当浇水、施肥和田间除草。
在小麦拔节期采用喷雾法进行田间接种,接种日期通常为4月下旬的16:00,确保土壤有一定湿度。若土壤太旱,需在接菌前2d进行灌溉,避免接菌时湿度不够而影响接种质量。当感病对照郑州5389病斑占叶片的表面积达100%时进行田间成株表型鉴定,即最终病害严重度(final disease severity,FDS)。其接菌流程与田间病害表型鉴定参考Li等[20]的方法。
1.3 DNA提取及分子标记筛选
利用CTAB法[21]提取抗病亲本周麦22、感病亲本铭贤169及其F2:3代家系小麦叶片全基因组DNA,其中每份材料分别选取3~5株进行DNA混合提取,并分别用 1×TE稀释成 40~50ng/µL工作液,作为大群体的筛选。此外,参照Hao等[22]提出的优选小群体(PSG)策略构建抗感小群体,结合田间FDS从中选取5个抗病(FDS 5%以下)和5个感病(FDS尽量选最大,接近 100%)的家系植株,分别提取叶片全基因组DNA作为抗感小群体。利用分布在所有小麦染色体组的1005对SSR标记对周麦22、铭贤169及抗感小群体进行多态性分子标记筛选。所需的PCR扩增体系、反应程序和PCR检测参考王佳真等[23]的方法。
1.4 遗传图谱构建及QTL定位
以多态性SSR分子标记筛选抗感亲本及其255个F2:3家系,将田间成株叶锈病FDS与筛选的基因型数据进行整理。利用 Excel软件分析田间成株期叶锈病表型的分布,采用QTL Cartographer软件进行遗传定位分析,其LOD值设为2.5,分析有效的QTL位点贡献率和加性效应等遗传信息。
2 结果与分析
2.1 周麦 22/铭贤 169 F2:3群体成株期叶锈病抗性表型分
依据 FDS分析可知,感叶锈病对照品种郑州5389在2个年度的FDS分别为90%和100%,表明在每个环境中抗病亲本周麦 22/感病亲本铭贤 169 F2:3群体的叶锈病发病良好。其抗病亲本周麦22的FDS在5%左右,感病亲本铭贤169的FDS在80%以上,255个家系的叶片病斑面积呈1%~100%的连续性分布,说明该群体呈现数量性状遗传特性(图1)。
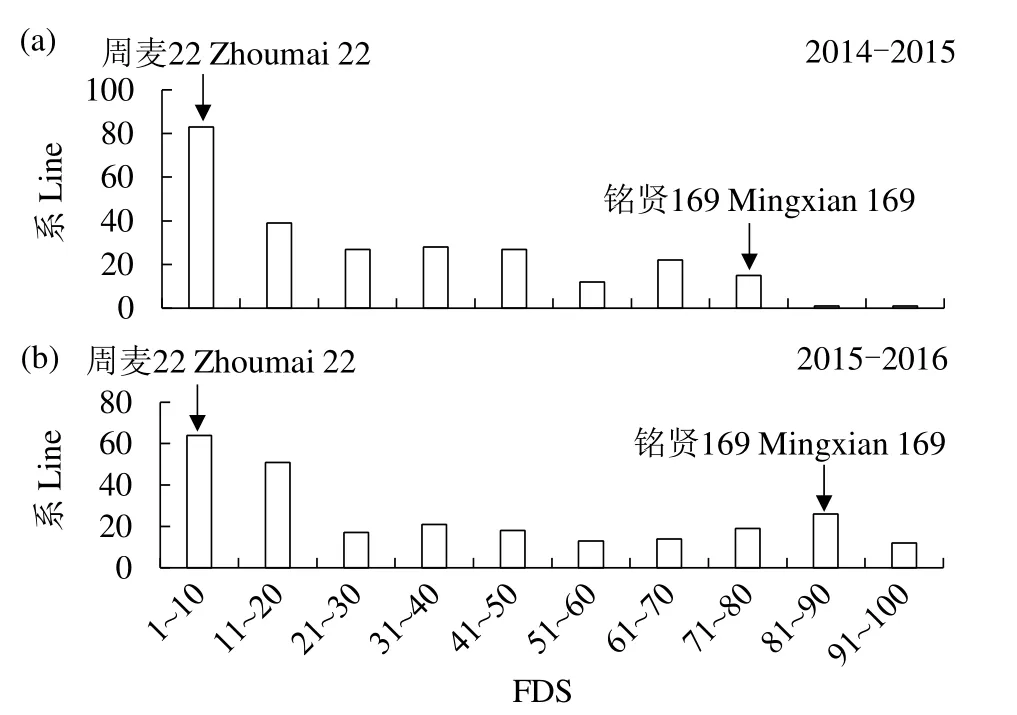
图1 周麦22/铭贤169 F2:3叶锈病最终严重度的频率分布Fig.1 Frequency distributions of the FDS of Zhoumai 22×Mingxian 169 F2:3 lines for leaf rust
此外,由表1可知,在2个年度中抗病亲本周麦22和感病亲本铭贤169的平均FDS分别为5%和85%,且在其群体中的平均FDS为30.5%~30.9%。同时,对2个年度中叶锈病FDS的相关系数进行计算,其呈现出显著相关(P<0.001),叶锈病FDS的相关系数(r)为0.95。

表1 周麦22/铭贤169 F2:3 255个群体在2个年度中的FDSTable 1 FDS for leaf rust of 255 F2:3 from Zhoumai 22×Mingxian 169 in two years
2.2 叶锈病成株期抗性QTL分析
利用分布于小麦染色体上的现存 1005对 SSR引物进行亲本多态性筛选,获得304对多态性SSR引物。将这些多态性标记应用于抗感小群体的筛选,获得交换值低于30%的55对多态性SSR标记(表2),并用其对周麦22/铭贤169的255个F2:3家系进行多态性筛选获得基因型。再结合2个年度的FDS结果,在群体中检测到2个成株抗性QTL,暂将其命名为QLr.hbau-1BL和QLr.hbau-2BS(表3和图2)。2个成株抗性QTL均来自抗性亲本周麦22,且这2个QTL均在2个年度中稳定被检测到(图2)。
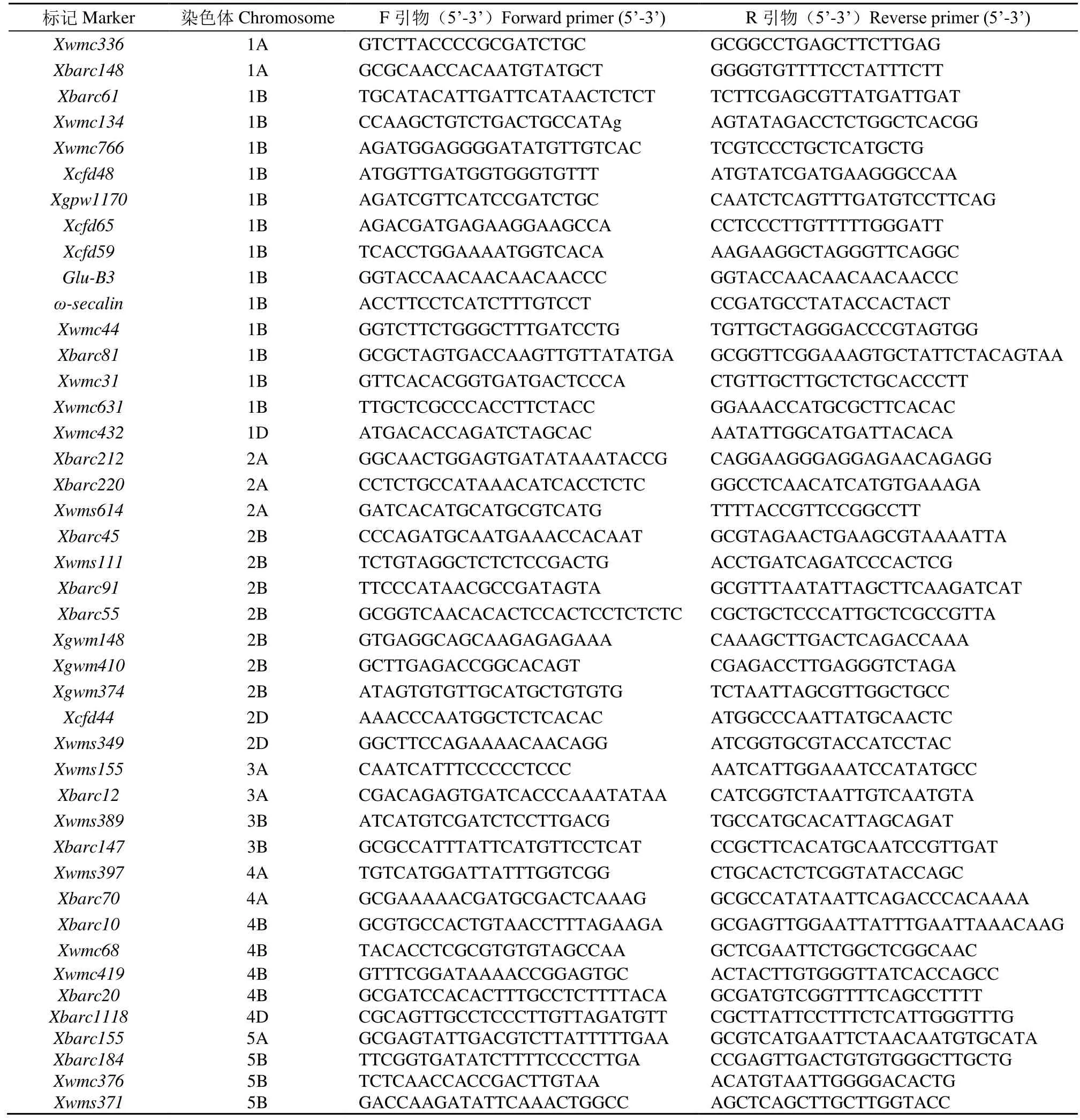
表2 55对SSR标记信息Table 2 Information of 55 SSR markers

续表2 Table 2 (continued)
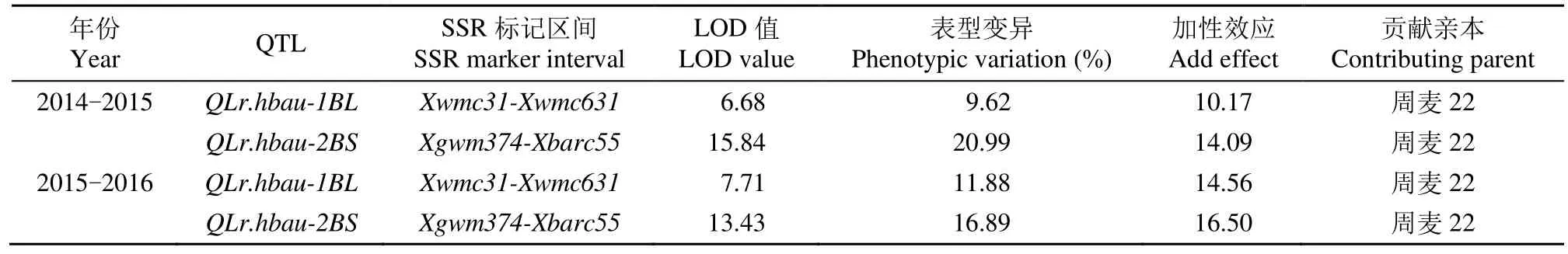
表3 周麦22/铭贤169 255个F2:3家系的叶锈病FDS的QTLTable 3 QTL for FDS to leaf rust in 255 F2:3 lines from Zhoumai 22/Mingxian 169

图2 位于1BL和2BS染色体上连续2年成株抗叶锈病Fig.2 Leaf rust adult-plant resistance QTL on chromosomes 1BL and 2BS in two years
由表3可知,第1个QTLQLr.hbau-1BL的侧翼标记为Xwmc31和Xwmc631,分别解释了2014-2015和 2015-2016年度表型变异为 9.62%和11.88%,加性效应分别为10.17%和14.56%。第2个QTLQLr.hbau-2BS侧翼标记Xgwm374和Xbarc55位于2BS染色体上,在2014-2015和2015-2016年度中均能检测到,分别解释了20.99%和16.89%的表型变异,其加性效应分别为14.09%和16.50%。结果表明,抗病品种周麦22中至少存在2个稳定有效的QTL,且表现出较高的抗性。因此,鉴定的QTL可作为抗叶锈病品种的亲本。同时,这些位点和它们紧密连锁的分子标记对于基因精细定位和分子标记辅助选择有重要作用。
3 讨论
本研究共检测到2个抗叶锈病QTL分别是QLr.hbau-1BL和QLr.hbau-2BS。QTL解释的总表型变异在叶锈病环境中为 9.62%~20.99%,证明它们对减轻病害严重程度的显著作用。目前,位于1B染色体上的基因或QTL有Lr46/Yr29[24]、QLr.caas-1BL[25]、QLr.pser-1BL[26]、QLr.hbu-1BL.2[27]及本研究中的QLr.hbau-1BL,根据它们的连锁或侧翼标记csLv46g22、Xwmc59-Xbarc213、Xwmc631-Xgwm268、Xbarc80-Xwmc728和Xwmc31-Xwmc631,确定其物理位置依次为670.2、667.2、637.2、685.1-686.7和610.35Mb。可知本研究中的QLr.hbau-1BL位点可能为新的成株抗叶锈位点。但由于标记区间较宽,还需要对图谱进行加密和进一步检验。对本研究中的第2个位于2B染色体上的抗病QTLQLr.hbau-2BS进行分析,有5个已知的抗叶锈病基因定位于小麦 2BS染色体上,分别为Lr13[28]、Lr16[29]、Lr23[30]、Lr73[31]和LrZH22[17]。Wang 等[17]在周麦 22中定位了苗期抗叶锈病基因LrZH22,与本研究中QLr.hbau-2BS位置一致,均位于紧密连锁的SSR标记Xbarc55和Xgwm374之间,抗性由已知抗叶锈病基因LrZH22提供,其抗病QTL均来自同一亲本周麦22。
周 8425B作为河南省骨干亲本广泛应用于我国小麦育种中。Zhang等[32]研究表明,LrZH22来源于周8425B的姊妹系周8425A,且具有全生育期抗性。Zhang等[33]对部分周麦系列品种进行了抗性鉴定和标记分析,发现携带抗病基因LrZH22的品种在田间均表现很好的抗叶锈性,目前已知携带LrZH22的周麦系列品种有周8425B、周麦11、周麦18、周麦22、周麦28和周麦30等。因此目前我国有效抗叶锈病基因为抗叶锈病基因LrZH22,抗性稳定,在不同抗性群体中的贡献率均很显著,有利于小麦抗叶锈病基因育种的研究。小麦品种中国春完整基因组序列公示将会极大促进小麦抗叶锈病基因的定位和克隆。同时根据中国春参考序列,可开发出紧密连锁或共分离分子标记,从而对定位的抗病基因进行精细定位,并用于分子标记辅助选择育种。
本研究中在周麦22中定位了2个抗叶锈病位点,可能还含有其他抗病位点,有待下一步研究。同时,本研究中找到的与抗病基因紧密连锁的分子标记可用于对抗病基因品种的筛选。
4 结论
在周麦22/铭贤169的255个F2:3家系群体中定位出2个抗病QTL,为QLr.hebau-1BL和QLr.hebau-2BS,分别解释9.62%~11.88%和16.89%~20.99%的表型变异。其中QLr.hebau-1BL可能为1个新的成株抗病位点,而QLr.hebau-2BS可能为全生育期抗病基因LrZH22。这2个抗病位点在2个年度中均表现出良好的抗性,为今后进一步基因挖掘及分子标记辅助选择育种提供良好基础。
猜你喜欢
中国蔬菜(2022年5期)2022-05-30
三农资讯半月报(2020年13期)2020-07-31
中国瓜菜(2019年8期)2019-09-19
发明与创新·大科技(2019年5期)2019-07-31
农民致富之友(2019年20期)2019-07-27
农民致富之友(2017年10期)2017-06-15
江苏农业科学(2016年6期)2016-07-25
江苏农业科学(2016年4期)2016-06-14
绿色科技(2016年7期)2016-05-14
中学生物学(2008年3期)2008-06-03
