
性状相关的分子标记在甘蔗分子育种中的应用研究
2022-05-05 05:24:12罗含敏熊发前丘立杭段维兴高轶静覃夏燕吴建明李杨瑞刘俊仙
作物杂志 2022年2期
罗含敏 熊发前 丘立杭 刘 菁 段维兴高轶静 覃夏燕 吴建明 李杨瑞 刘俊仙
(1广西壮族自治区农业科学院甘蔗研究所,530007,广西南宁;2农业农村部广西甘蔗生物技术与遗传改良重点实验室/广西甘蔗遗传改良重点实验室,530007,广西南宁;3广西壮族自治区农业科学院经济作物研究所,530007,广西南宁)
甘蔗(Saccharum officinarumssp.)被广泛种植于热带和亚热带地区,不仅是重要的禾本科糖料作物,也是重要的生物质与能源作物和经济作物[1-2]。广西是我国最大的甘蔗生产基地,自2000年以来,种植面积占我国糖料作物总面积的 70%以上,另外,甘蔗糖占我国食糖总量的90%以上,因此,甘蔗产业对保障国家蔗糖供给安全有着举足轻重的作用。
甘蔗栽培由新植和宿根组成,国内的一般可收获3年(1年新植和2年宿根),而国外多数产蔗国多于3年。其单产为单位面积有效蔗茎的总重量,由株高、分蘖、茎径、锤度和单茎重等多基因调控的重要数量性状组成[3-4],因此,确保甘蔗种植稳产增产的重要途径之一是培育优良新品种,高产、高糖、宿根性好和抗性强等优良性状需综合兼顾,不同时代甘蔗品种的更迭对提高甘蔗产量意义非凡[5]。然而,甘蔗新品种选育和改良主要采用传统的有性杂交程序(图1),该程序存在选育周期长、经验性居多和效率低等问题,育成1个甘蔗新品种需要8~10年,甚至更长的时间。此外,甘蔗栽培种的基因组高度杂合,染色体呈非整倍性[6],而当前甘蔗高贵化育种的遗传基础狭窄[7-8],这些加大了甘蔗品种选育和遗传改良的难度[9]。虽然可通过近缘属和远缘属杂交引入更多的优异基因资源以拓宽甘蔗遗传基础[10-11],但重要数量性状在甘蔗杂交后代中易受环境因子影响而表现不稳定[12-13],这也加剧了甘蔗品种选育难度。由此可知,甘蔗常规品种选育的瓶颈显而易见。目前,随着分子生物学及基因组学技术和手段的快速发展,甘蔗栽培种R570和细茎野生种割手密(Saccharum spontaneumL. AP85-441)基因组的公布[6,14],且越来越多甘蔗栽培品种(系)RNA-Seq数据的发表[15-18],给基于大数据挖掘与开发甘蔗性状相关的分子标记在甘蔗育种上的利用带来了新的希望和机遇。虽然基于基因组的分子标记极大地促进了重要作物农艺性状的遗传选育[19],然而,甘蔗分子标记辅助育种仍面临着许多挑战。本文通过分析甘蔗分子标记辅助育种的现状及其瓶颈,总结应用于甘蔗的分子标记种类及其存在的问题,阐明分子标记在甘蔗遗传连锁图谱构建中的作用及其研究状况,进而从甘蔗产量与蔗糖分性状相关 QTL的定位、甘蔗主要抗病基因的定位及其在抗病分子育种中的应用概况、关联分析方法在甘蔗重要农艺性状研究中的应用等方面对性状相关的分子标记进行综述,为分子标记技术助力甘蔗新品种选育和提高育种效率提供理论参考。
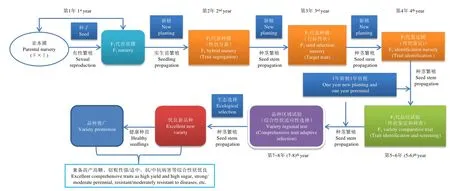
图1 甘蔗新品种的常规杂交选育流程Fig.1 The process of conventional cross breeding for new sugarcane varieties
1 甘蔗分子标记辅助育种现状及瓶颈
分子标记辅助育种技术可提高作物育种效率,也能准确地进行目标农艺性状的定向选择。虽然分子标记在甘蔗及其种质遗传多样性研究中已有许多报道,但目前只有抗褐锈病基因标记能够应用于甘蔗分子标记辅助育种中[20]。究其原因主要有 4点,一是甘蔗遗传背景复杂且基因组高度杂合与庞大,二是甘蔗高贵化代表性品种全基因组序列尚未测序并组装完成,三是缺乏简单实用高效的 DNA分子标记,四是遗传连锁图谱构建密度极度不饱和。这些是导致甘蔗种质资源的分子精准鉴定和评价、高密度遗传连锁图谱的构建、重要性状QTL精细定位、关键基因的图位克隆以及与之紧密连锁的分子标记开发等都存在困难的原因,因此,很难真正达到甘蔗分子标记辅助育种和分子聚合育种的目的。
2 甘蔗的分子标记种类及其存在问题
目前,虽然有越来越多与甘蔗相关的转录组测序(RNA-Seq)报道,但甘蔗基因组的de novo测序及其组装尚未完成,而已公布的相关甘蔗基因组在育种研究工作中的应用有待挖掘。因此,为了加快甘蔗分子育种的进程,针对目标性状开发简单、实用和高效的分子标记十分必要。目前应用在甘蔗种质资源分子鉴定和评价上的分子标记技术有RAPD[21]、ISSR[22]、AFLP[23]、SRAP[24]和 SCoT[25],虽然这些分子标记技术在甘蔗DNA多态性检测上发挥着重要作用,也可用于甘蔗种质资源的遗传多样性分析和构建其DNA身份证数据库,但RAPD技术的重复性较差[26],AFLP技术的操作流程较复杂且对科研人员及实验仪器设备要求较高[27],ISSR[22]、SRAP[28]和 SCoT[29]虽然操作简单但多态性不够丰富,且都是显性分子标记技术,扩增条带数较多,PCR扩增结果易受环境影响,其重复性和稳定性不够好。许多研究表明,SSR标记在甘蔗上有丰富的多态性,已被广泛应用于甘蔗遗传多样性分析[30]、分子遗传连锁图谱构建[31-32]和QTL定位[33],虽然国内外已开发出了数量较多的甘蔗SSR标记引物对,但开发效率相对较低。现今,随着高通量测序技术的快速发展和测序成本的大幅下降,转录组测序或者简化基因组测序在规模化开发甘蔗SSR标记上有巨大潜力,但甘蔗基因组庞大且复杂(约10Gb),目前开发出来的SSR标记引物对数量还远远不够,且在任意2份甘蔗材料之间具有多态性的SSR标记引物数量更是有限。此外,扩增出来的片段大小差异小,需要通过较为复杂的后期技术和手段(如聚丙烯酰胺凝胶电泳或毛细管电泳等)分离。这些缺点严重制约了分子标记在甘蔗种质资源的精准鉴定及评价、高密度遗传连锁图谱的构建、重要性状的 QTL定位、基因的图位克隆以及分子标记辅助育种中的应用。
3 分子标记在甘蔗遗传连锁图谱构建中的作用
高密度和高分辨率的遗传连锁图谱是进行数量性状基因精细定位(quantitative trait loci,QTL)、分子标记辅助选择育种、重要基因克隆及比较基因组研究的基础[34]。虽然分子标记技术的开发与应用促进了甘蔗属作物遗传连锁图谱的构建,但由于甘蔗属及其近缘属一般为复杂的多倍体或高度杂合的非整倍性多倍体作物,染色体数多达100~150条,基因组庞大且高度杂合,因此,其遗传连锁图谱的构建研究滞后于其他作物[31,35-37]。据保守估计,由于甘蔗的基因组遗传距离为17 000~18 000cM[37-38],理论上构建1个平均距离为10cM的饱和遗传连锁图谱,大概需要1700~1800个单双剂量标记。目前,甘蔗遗传连锁图谱的构建主要利用RFLP、RAPD、SSR和AFLP分子标记技术。
割手密最早的遗传图谱是通过RFLP分子标记技术构建的[39],该图谱包含216个单剂量标记(single dose marker,即只在父本或母本一方存在的标记)和44个连锁群。利用RAPD分子标记技术构建的割手密遗传连锁图谱包含279个单剂量标记和42个连锁群[40]。然而,通过整合这2种分子标记技术构建的割手密遗传连锁图谱则包含442个单剂量标记和64个连锁群[41],其密度和分辨率明显优于单一的分子标记技术。以此估算割手密无性系“SES208”(2n=80)的基因组长度为3300cM,而通过单一RAPD分子标记技术推断的仅为 2550cM[40],其基因组大小显然与甘蔗属预计相差甚远。由此可见,利用单一的分子标记技术构建的甘蔗遗传连锁图谱的基因组覆盖率小,无法精细定位。
在栽培甘蔗分子遗传连锁图谱方面,分子标记SSR和AFLP整合用于构建甘蔗杂交父母本和甘蔗栽培品种杂交后代[37,42],以及甘蔗商业品种Co419与野生种割手密Y75-1-2的杂交F1和回交BC1群体的遗传连锁图谱的构建[31],将新型分子标记技术TRAP(target region amplified polymorphism)融入这 2种技术整合体系后,以美国路易斯安那州主栽品种LCP85-384作为遗传材料构建的遗传连锁图谱包含1111个多态性标记,其中有773个为单剂量标记,分布在108个连锁群上,总遗传距离为12 313cM[32],这间接证实甘蔗基因组庞大的事实[6]。另外,新型分子标记EST-SSR与AFLP联合构建的甘蔗栽培品种(IAC66-6×TUC71-7)后代遗传连锁图谱的总遗传距离也有4843.19cM[43]。可见,新型分子标记技术的研发及多种分子标记技术的联合应用对构建甘蔗高密度遗传连锁图谱意义重大。
综上可知,单一分子标记技术的应用对甘蔗属遗传连锁图谱构建的贡献有限,而不同分子标记技术的整合应用效果更为明显。此外,不同甘蔗种属间及其杂交后代的遗传距离差异很大,这暗示甘蔗基因组极其复杂多变,对构建高密度和高分辨率遗传连锁图谱造成极大挑战,重要农艺性状功能基因精细定位的难度较大。
4 甘蔗产量与糖分性状相关QTL的定位
产量和糖分相关性状一直是甘蔗育种研究的热点,实现高产、高糖也是甘蔗育种和栽培技术的瓶颈[44-45]。虽然分子标记技术的快速发展给作物遗传连锁图谱的构建提供了可靠的技术支撑,且利用分子标记开展性状相关QTL定位的报道并不鲜见,也在甘蔗属作物上得到了应用[46],但目前国内外关于甘蔗产量和糖分相关性状 QTL定位的报道相对较少。
Ming等[47]利用甘蔗栽培种与割手密杂交后代的分离群体构建了包含735个RFLP标记的遗传连锁图谱,结合2年表型数据的统计分析和遗传标记的连锁分析,定位到了甘蔗产量、糖分、茎重、有效茎数、纤维含量和灰分含量等性状的QTL。Pinto等[48]利用优良种质SP80-180(父本)和SP80-4966(母本)的杂交后代群体为材料,也构建了2年生育期的甘蔗RFLP遗传连锁图谱,并定位了120个与单位面积含糖量和产糖量、纤维含量以及蔗茎产量相关的QTL,其中有26个QTL在2年中均有定位到,有32个QTL仅在宿根蔗中定位到,此外,还鉴定了50个与这4个性状具有双基因上位互作的标记,进一步分析发现,1个与蔗糖合成相关的QTL与蔗茎产量呈负相关,但与单位面积含糖量和产糖量呈正相关。由于数量性状在甘蔗杂交后代中易受环境影响,表现不稳定[12-13],故检测中不能忽略QTL、收获年限和种植地点之间交互作用的影响。针对这一问题,Pastina等[49]提出了一种联合区间作图法与混合模型分析法对甘蔗多重–多年收获–多点试验数据的QTL检测策略,通过该策略发现甘蔗分离群体后代在2个试验点下连续3年收获的46个QTL,其中13个为蔗茎产量,14个为糖产量,11个为纤维分,8个为蔗糖分。另外,这些QTL受收获互作、地理位置互作及二者均互作的影响均达显著水平。在甘蔗的3个生育周期中,Singh等[50]对7个不同地理环境下甘蔗产量相关性状开展了更加全面细致的定位,连锁分析发现杂交后代分离群体中有8个群体与母本Co86011连锁,16个群体与父本CoH70连锁,基于SSR分子标记构建了总长为2606.77cM的遗传连锁图谱,还精细定位到了31个与父母本在遗传影响上一致的QTL,分别是2个苗期锤度 QTL、7个田间锤度 QTL、6个蔗糖分QTL、4个有效茎数QTL、1个茎长QTL、3个茎粗QTLs、6个节间数QTL和2个绿叶片数QTL,其中与糖含量性状相关 QTL连锁的 SSR标记在UGSM31548和 UGSM31649之间,这暗示甘蔗农艺性状标记的遗传距离比较大。
然而,Hoarau等[51]以甘蔗栽培种R570的自交系群体为标记对象,利用1000个AFLP标记构建了覆盖R570一半基因组的遗传连锁图谱,并定位到了4个甘蔗重要农艺性状的QTL,包括株高、茎数、茎径和锤度相关,这些 QTL的个体差异解析了整个表型变异的3%~7%,且可能受40个假定的数量性状等位基因(quantitative trait allele,QTA)调控。为了更加精准地挖掘甘蔗产量相关 QTL的连锁基因,Balsalobre等[52]构建了基于223个连锁群体测序基因分型(genotyping-by-sequencing,GBS)数据的QTL遗传连锁图谱,通过GBS测序分析开发了3433到15906个高质量的标记,利用其中的 993个单剂量标记累积绘制的图长为3682.04cM,平均标记密度为3.70cM,定位到了7个产量构成性状相关的QTL,并关联到可溶性固体含量QTL和纤维含量QTL的候选基因。
总之,杂交后代分离群体密度及其个体间遗传差异大小与遗传连锁图谱的总遗传距离和平均标记密度密切关联,而甘蔗杂交后代分离群体个体差异大致使其分子标记间隔距离较大,这给控制甘蔗重要性状 QTL的主效基因定位和挖掘带来了许多困难。
5 甘蔗主要抗病基因的定位及其在抗病分子育种中的应用
目前,随着气候的变化,全球蔗区甘蔗病害严重频发,对产量和蔗糖分造成了不可逆转的损失[53-56]。因此,甘蔗抗病育种迫在眉睫,然而抗病品种的选育进展缓慢。褐锈病是甘蔗最严重的叶部病害之一,自Daugrois等[57]报道,Bru1为甘蔗抗褐锈病的主效基因后,Asnaghi等[58]采用AFLP分子标记/BSA测序的联合方法获得了Bru1基因目标区域的分子标记,在该基因两侧共获得8个AFLP遗传标记,而其中与Bru1基因最近的2个遗传标记的遗传距离分别为1.9和2.2cM。McIntyre等[59]利用甘蔗抗性基因类似物的序列对遗传图谱进行标记验证,共检测到31个标记,其中3个标记与甘蔗抗褐锈病显著相关,还定位到解释甘蔗抗褐锈病 7%~18%表型变异的 QTL[60]。Raboin等[61]利用抗病品种 MQ76-53和感病品种 R570杂交后代的分离群体作为材料,综合运用AFLP、SSR和RFLP技术构建了由 424个单剂量标记组成的 MQ76-53遗传连锁图谱,定位到了1个新的抗褐锈病的主效QTL和1个微效QTL,但与主效QTL最近的AFLP遗传标记(acgcta16)的距离有23cM。可见,这些与Bru1基因相关的遗传标记的分辨率不高,相关的QTL所解析的表型变异也大,在抗锈病育种材料筛选的难度较大。随着生物技术的发展,Asnaghi等[62]和Le Cunff等[63]均运用比较基因组学策略来加密甘蔗目的基因的遗传图谱,最终获得Bru1基因区域内高分辨率遗传图谱,并将Bru1基因定位于甘蔗第7条染色体0.42cM区域内,发现该区域与高粱第4号染色体上的约225kb区域和水稻第2号染色体短臂上的600kb区域具有高度同线性,这是目前与Bru1基因共分离最近的遗传标记距离,分别达到0.28和0.14cM的高分辨率。说明基于同源功能基因序列靶向开发目标标记是可行的,这对于缺乏参考基因组的作物具有借鉴意义。
迄今,随着甘蔗抗褐锈病Bru1基因被广泛地深入研究,这一抗性标记的开发也逐渐被成功应用于甘蔗分子标记辅助育种中,并陆续被法国[64]、美国[65-66]、阿根廷[67]、中国[68-72]、巴西[73]等国家的科研工作者应用于分子育种研究,这意味着在甘蔗抗锈病育种中迈出了坚实的一步,并给其他农艺性状的分子标记辅助育种提供了参考。
黄点病是仅次于褐锈病的叶部严重病害,Aljanabi等[33]利用抗病品种“M134/75”和感病品种“R570”杂交的227个后代个体作为作图群体,利用AFLP和SSR标记构建了包含557个单剂量标记的甘蔗栽培种M134/75的遗传连锁图谱,并在连锁群LG87上定位到了一个抗甘蔗黄点病主效基因CIR12284,该基因位于标记aagctt3和actctc10之间,而与距离最近的actctc10标记的遗传距离也有14cM。而Parmessur等[74]通过融入EST-SSR分子标记对 227个作图群体进行了基因分型,再经过RAD-seq测序分析得到含528个单剂量标记和2777个RAD-seq标记的甘蔗M134/75的遗传连锁图谱,经Software Joinmap 4.1分析后产生总长8277cM(2046标记)由286个连锁群组成的图谱,随后,基于EST-SSR分子标记在M134/75的LG28上鉴定到1个抗黄点病的主效QTL,其最近的遗传标记距离为5cM,分辨率提高超过2.5倍,且这个QTL解析了30%的表型变异。可见,采用不同的标记技术,靶向基因遗传标记距离的差距较大,这可能与物种染色体的倍性及复杂度有关。
据不完全统计[75],黑穗病是我国甘蔗生产上最主要的病害,发生严重年份造成甘蔗产量损失达20%~50%,蔗糖分降低 0.3%~0.5%(绝对值),宿根蔗受害尤为严重。抗黑穗病甘蔗新品种的选育是减轻黑穗病对甘蔗产业危害的根本途径,但由于甘蔗遗传背景复杂,迄今为止,关于甘蔗抗黑穗病基因的定位、克隆及其分子标记辅助育种的研究进展十分缓慢,相关的研究报道甚少[76]。许莉萍等[77]结合RAPD技术和BSA法,获得了1个与甘蔗抗黑穗病生理小种2基因连锁的分子标记,并将其转化为SCAR标记。高轶静等[78]则通过SSR技术与BSA法的整合,获得了1个与甘蔗抗黑穗病基因连锁的SSR标记。
6 关联分析在甘蔗重要农艺性状研究中的应用
根据孟德尔和摩尔根提出的遗传定律,调控甘蔗重要性状的遗传基因既存在分离与自由组合,也会发生连锁与互换,而甘蔗基因组庞大且高度杂合,染色体存在非整倍性现象,控制重要性状的基因发生连锁遗传的概率较高。虽然已有不少研究者针对甘蔗属重要性状进行了遗传连锁图谱构建和QTL定位研究,但农艺性状一般受多个数量性状位点或基因共同调控[79],理论上发生连锁基因的不平衡遗传在甘蔗上的体现更为突出[80-81]。然而,基于连锁不平衡(linkage disequilibrium,LD)的性状关联分析在甘蔗上的研究报道较少。
起初,Jannoo等[82]对59份甘蔗商业品种材料进行LD分析,通过38个RFLP探针揭示出这些材料间DNA的多态性,发现33个基因座与42例双位点事件有关联,且它们间的遗传距离大多不超过10cM,相对应的等位基因在起源品种中至少有1个被发现,这暗示它们存在血缘关系,即当基因座紧密相连时,由此产生的优异等位基因组合在随后的杂交中得以维持,也证实了基于LD在分子标记与甘蔗农艺性状之间进行关联分析的可行性。随后,Raboin等[83]对基于LD的关联分析在绘制甘蔗基因组图谱上的价值进行了评估,以品种“R570”为对照,使用42对AFLP标记引物对72个甘蔗现代栽培种进行了亲缘关系分析,基于1537个多态性标记的关联分析结果显示,许多连锁标记因遗传多倍性而无法被识别。然而,就共分离群体而言,同一个被分配的标记,这些甘蔗现代栽培种样本的LD关联分析结果和对照种后代的遗传连锁分析表现出一样的效果,说明甘蔗现代栽培种性状的LD关联分析不受遗传多倍性的影响。另外,Wei等[84]运用1068个AFLP标记和141个SSR标记对154份甘蔗品系进行关联分析,通过回归分析获得了与澳大利亚4种最重要甘蔗病害关联的分子标记,其中,4个标记解释了甘蔗白叶病和根腐病32%的表型变异,5个标记解释了甘蔗斐济病26.2%的表型变异,11个标记解释了甘蔗抗黑穗病 58.6%的表型变异(其中4个效应较大的标记解释了53.7%的表型变异),同时还提出种群结构在标记–性状关联分析中应该被考虑。
近年来,Banerjee等[85]利用基因组的123条ESTSSR引物对印度亚热带蔗区的102个甘蔗品种进行群体结构、亲缘关系和标记–性状 LD关联分析等研究,发掘了989个SSR标记位点。随后,基于混合线性模型分析,分别鉴定了4个和7个与茎径、株高及有效茎数显著相关的标记,另外,有 15个标记连续3年稳定地解释了57%的有效茎数、34%的茎粗、27%的茎长、20%的蔗糖分含量和19%的节数等甘蔗产量和蔗糖性状变异,进一步指出甘蔗家系分组的群体结构对利用基于基因数据的关联分析进行品种选育至关重要,这一观点与Wei等[84]一致。Singh等[86]也利用LD关联作图法获得4个可解释甘蔗抗赤腐病10%~16%表型变异的SSR标记,并通过EST测序与高粱基因组进行共线性比对以鉴定其候选基因,发现标记位点对应的基因编码重要的植物防御相关蛋白,认为它们可能是潜在的抗性候选基因。
Fickett等[87]利用基于全基因组关联分析(genome wide association study,GWAS)开发的SNP和InDel标记对甘蔗的11个产量性状和蔗糖分性状进行研究,发现56个标记与蔗糖分性状显著相关,且在多个性状上也表现出一致的显著性,随后,这些标记在更加多样化的群体中得到了验证,可用于甘蔗品系的分子标记辅助选育,Racedo等[80]和Yang等[81]的研究结果可作为最新的佐证。You等[88]也基于 SNP标记技术开发出了甘蔗高通量100K SNP矩阵试剂盒,用于构建3个甘蔗亲本的遗传连锁图谱,并对来自这3个亲本杂交及自交的469份后代材料进行关联分析,鉴定出了27个抗病基因,其中18个与SCYLV(Sugarcane yellow leaf virus)抗性相关的QTL关联,此外,这469份材料基因分型的结果显示,样本 SNP多态性的比例高达77.04%。可见,SNP阵列可作为高度多倍体甘蔗高通量基因分型的有效遗传标记工具,将在重要性状的关联分析及其调控基因挖掘上发挥巨大作用[89]。
7 展望
基因组是作物分子育种研究的关键,也是突破传统育种的重要基础。甘蔗分子标记辅助育种进展十分缓慢,其真正原因在于甘蔗遗传背景复杂且基因组庞大与高度杂合,甘蔗高贵化代表性品种全基因组尚未测序并组装完毕,缺乏高质量甘蔗参考基因组序列,进而导致本身或借鉴其他作物基于基因组的许多分子标记的开发和应用难度大,标记的分辨率和精确性普遍较低。当前在甘蔗遗传研究上应用最多的是SSR分子标记,但多数集中在遗传多样性分析[90]和真假杂种鉴定上[91],关于其他传统分子标记技术的应用研究也有不少报道,然而,这些分子标记技术普遍存在通量低、分辨率和多态性不高等问题,严重制约其应用于甘蔗分子育种,尤其是性状关联优异基因的精细定位。最近,Yang等[92]开始尝试利用 GBS测序技术筛选出甘蔗抗性相关基因并进行了SNP标记开发,将这些标记在F1群体中进行验证,关联得到2个与抗橙锈病QTL显著相关的SNP标记。
随着甘蔗属相关全基因组序列图谱的陆续公布,如甘蔗栽培种 R570的单倍体基因组序列[6]和甘蔗细茎野生种单倍体割手密AP85-441的全基因组序列[14],使得高通量开发SSR和SNP标记成为可能。但我们初步研究发现,当将这2个基因组序列作为甘蔗参考基因组时,基于样本与参考基因组的比对结果开发的 SNP标记质量并不高,应用效果也不好。只有在获得了高质量甘蔗参考基因组序列的前提下,通过重测序和简化基因组测序(如SLAF、RAD和GBS)等高通量基因分型手段和生物信息学对甘蔗遗传群体及其自然群体的重要性状进行基因定位和关联作图,才能准确并高通量地挖掘出其中的变异组成,进而高效地开发相应的遗传标记(如SNP、InDel、SV、PAV和CNV),再结合群体结构聚类分析和表型组学分析的精准鉴定,采用连锁分析或者关联分析筛选出与甘蔗重要农艺性状紧密连锁的分子标记。
高密度的遗传连锁图谱和高分辨率的遗传标记是甘蔗分子标记辅助育种研究取得突破的关键。关联分析则是重要性状相关标记精准鉴定的重要手段,然而,重要性状的基因解析需要高质量且精细的全基因组序列图谱,即精准挖掘与性状关联标记的候选基因是甘蔗分子设计育种或者分子标记辅助育种策略的突破。目前,在甘蔗上,只能通过BSA法、蛋白质组、RNA-seq和代谢组测序等多种组学方法,鉴定出可能的候选基因,然后对其进行多重qRT-PCR表达验证来筛选关键基因,并对关键基因进行转基因功能验证或基因编辑研究。利用高通量测序技术对甘蔗重要性状进行连锁和关联分析,就目前拥有的高质量甘蔗参考基因组而言[6,14],仍存在测序成本偏高的问题,由于甘蔗基因组庞大(约10Gb),约为水稻(400Mb)的25倍,目前水稻完全可以通过高通量测序技术完成各种性状的基因定位研究,但甘蔗的定位难度很大。现今,高通量测序技术的迅速发展和成本不断降低以及表型组学的兴起,使得甘蔗重要性状基因的挖掘及其分子标记开发成为可能。这些技术将促进与甘蔗重要性状紧密连锁的分子标记和关键基因的编辑研究应用于甘蔗分子标记辅助育种和遗传改良中,从而提高甘蔗育种效率和准确性。
猜你喜欢
儿童故事画报·自然探秘(2022年12期)2022-11-24 10:10:12
基层中医药(2021年3期)2021-11-22 08:08:04
现代装饰(2020年7期)2020-07-27 01:27:44
特别健康(2018年3期)2018-07-04 00:40:08
NBA特刊(2018年7期)2018-06-08 05:48:32
现代装饰(2018年4期)2018-05-22 02:57:23
中国糖料(2016年1期)2016-12-01 06:48:57
浙江柑橘(2016年3期)2016-03-11 20:12:48
汽车维护与修理(2015年6期)2015-02-28 12:16:55
温州医科大学学报(2014年7期)2014-07-18 02:43:24
