
谷子miR169家族及其靶基因的鉴定与功能分析
2022-05-05 05:24:16康庆芳赵孟瑶张凤洁武强强马芳芳王宇珅韩渊怀王兴春李雪垠
作物杂志 2022年2期
卢 平 康庆芳 赵孟瑶 张凤洁 武强强马芳芳 王宇珅 韩渊怀 王兴春 李雪垠
(1山西农业大学生命科学学院,030801,山西晋中;2山西农业大学农学院,030801,山西晋中)
谷子(Setaria italica)属单子叶禾本科二倍体植物(2n=2x=18),是我国重要的杂粮作物之一。谷子起源于中国,有悠久的栽培历史,具有耐旱和耐贫瘠的特性,是发掘抗旱基因和研究作物抗旱分子机制的优异种质资源[1]。在拟南芥和水稻中都没有发现 C4光合作用等重要过程,因此需要新的模式物种[2]。谷子是C4植物,具有生长能力强、CO2利用率高和耐旱等优点,并且具有C4途径。谷子具有二倍体基因组小、自花授粉和种子产量多等优点,同时谷子突变体xiaomi的发现也解决了谷子生长周期较长和植株较大等不适合作为模式生物研究的问题[3]。并且随着谷子基因组测序的完成,谷子也逐渐成为一种模式植物[4-5]。
miRNA是一类小RNA(sRNA),是由内源基因编码、长度约为22个核苷酸的非编码单链RNA分子。miRNA广泛参与动植物基因转录后的基因表达调控,通过与靶 mRNA的靶位点结合抑制基因表达。在植物中,miRNA参与器官建成、生长发育、开花与育性转换、物质运输和抗逆响应等多个生理生化过程[6-7]。虽然植物miRNA生物发生、降解、功能及调控机制已得到了较清晰的阐述,但目前关于植物miRNA作用机制的认知仍处于初级阶段[8]。谷子抗旱性突出,是研究作物耐旱性的理想模型系统,在干旱胁迫下,14个miRNA被上调,4个miRNA被下调。此外,已鉴定出72个潜在的新miRNA,其中3个在干旱条件下差异表达[9]。miR169是植物中较大的miRNA家族,miRNA数据库miRBase(http://www.mirbase.org/)已收录拟南芥、水稻、玉米、大豆、番茄、油菜、大豆、玉米和山核桃等35个物种的400多个miR169基因,这些基因在植物的生长发育及非生物胁迫应答的调控过程中起重要作用。拟南芥miR169家族在胁迫诱导植物开花中起关键作用,miR169d在拟南芥中过表达导致提前开花[10];水稻miR169基因与盐胁迫应答和干旱诱导相关,还负调控水稻对稻瘟病菌的免疫[11-12];玉米zma-miR169/ZmNFYA14参与根发育和盐胁迫应答[13];大豆miR169基因参与盐碱和干旱等非生物胁迫应答反应[14-15];番茄miR169基因能响应致病疫霉菌的侵染[16];油菜miR169d基因可能通过调控靶基因的表达而参与油菜的早花发育[17];杨树miR169基因家族的靶基因包括转录因子、蛋白激酶、抗病蛋白和泛素连接酶基因等,说明其广泛参与杨树多种生命活动的调控[18];葡萄miR169的靶基因主要是 NF-YA转录因子基因[19],是葡萄生长发育过程中抗逆境胁迫响应的主要调控因子;山核桃miR169通过作用其下游基因调节花的发育。由于对miR169尚缺乏系统研究,至今仅有部分植物miR169家族成员的功能被解析。
NF-YA转录因子基因是miR169的主要靶基因。NF-Y是真核生物中一类广泛存在的转录因子,能够特异性结合顺式作用元件CCAAT-box,进而对其下游靶基因进行正向或负向调控[20]。NF-Y作为保守的转录因子在植物的生长发育和对逆境胁迫响应中扮演着重要的角色[21]。NF-Y是1个三聚体转录因子,包括NF-YA、NF-YB和NF-YC 3个亚基。NF-YA作为三聚体最为重要的组成因子,在N端富含色氨酸或苏氨酸,有2个保守域NF-YB/NF-YC结合域和DNA结合域[22]。研究[12]发现,在一些NF-YA基因的3′UTR存在miR169的切割靶位点,miR169可以调控NF-YA基因。拟南芥ath-miR169/NF-YA5调控模块可以响应干旱,miR169表达受到抑制,NFYA5基因也就脱离了miR169的控制而上调表达,进而使植物抗旱性增强[23]。
miR169家族在植物中主要参与非生物胁迫应答调控,还能诱导植物提前开花等。目前关于谷子miR169(sit-miR169)的相关研究较少。鉴定sitmiR169家族是解析miR169参与谷子生长发育和胁迫响应机制的重要基础。本文利用生物信息学方法对sit-miR169家族的染色体分布、系统进化、碱基保守性、二级结构和sit-miR169及其靶基因表达情况进行分析,为进一步研究谷子miR169家族的进化和功能奠定理论基础。
1 材料与方法
1.1 谷子miR169家族的染色体定位分析
从 PmiREN[24]数据库(http://www.pmiren.com/)下载谷子sit-miR169家族成员的前体序列、成熟序列及染色体定位信息。利用TBtools[25]软件绘制sitmiR169家族基因的染色体定位图。
1.2 谷子miR169家族的系统进化分析
利用ClustalX 2.1对谷子sit-miR169前体序列进行多序列比对;从PmiREN数据库下载水稻和拟南芥miR169的前体序列,对谷子、水稻和拟南芥miR169的前体序列进行多序列比对。利用MEGA X和邻接法(Neighbor-Joining)构建系统发育树,Bootstrap值设为1000。
1.3 谷子miR169序列碱基保守性分析
利用WebLogo 3[26](http://weblogo. threeplusone.com/create.cgi)分析sit-miR169前体序列和成熟序列的碱基保守性。
1.4 谷子miR169前体二级结构预测
利用 RNAfold(http://rna. tbi. univie.ac.at./cgibin/RNAfold.cgi)在线预测sit-miR169前体二级茎环结构,用折叠算法和基本选项选择最小自由能(MFE)和配分函数。
1.5 谷子miR169组织特异性表达分析
从PmiREN数据库下载谷子sit-miR169家族成员在根、叶和花中的表达量数据,利用TBtools将sit-miR169的表达数据绘制成热图。
1.6 谷子miR169靶基因预测
利用psRNATarget[27](http://plantgrn.noble.org/psRNATarget/)预测靶基因。以自带 cDNA文库Setaria italica(foxtail millet)、transcript、JGl genomic project、Phytozome 13、312 v 2.2为搜索数据源预测sit-miR169靶基因,得分模式使用Schema V1(2011 release),Expectation≤3,UPE≤25。将预测得到的靶基因进行整理,利用NCBI(https://www.ncbi.nlm.nih.gov/)对候选靶基因进行功能注释。
1.7 谷子miR169靶基因的组织特异性和环境诱导表达分析
从Phytozome数据库(https://phytozome.jgi.doe.gov/pz/portal.html)下载上述候选靶基因在不同组织和条件下的表达量数据,并绘制热图。
2 结果与分析
2.1 谷子miR169家族基因基本信息及染色体分布
根据PmiREN数据库,谷子基因组共有15个sitmiR169家族成员(表1)。sit-miR169k和sit-miR169o成熟序列碱基数为20,其余13个sit-miR169成熟序列碱基数均为21。15个sit-MIR169分布在除5和9号染色体外的7条染色体上(图1),其中位于6号染色体上的sit-miR169最多(5个)。

表1 sit-miR169家族成员的基本信息Table 1 Basic information of sit-miR169 family members
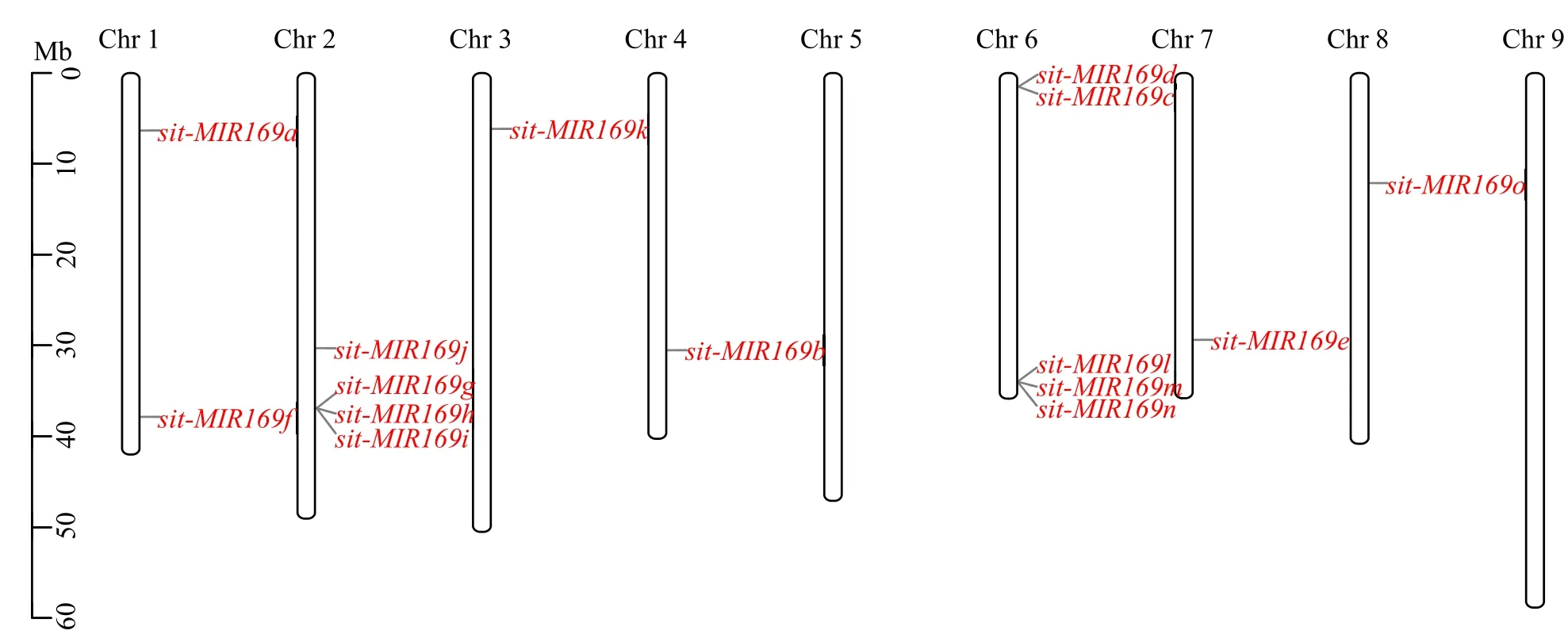
图1 sit-MIR169基因的染色体定位Fig.1 Chromosome location of sit-MIR169 genes
2.2 谷子miR169家族的系统进化分析
sit-miR169前体序列(sit-MIR169)系统进化分析(图2)显示,sit-miR169家族大致可以分为 2簇,sit-MIR169a、sit-MIR169e、sit-MIR169f和sit-MIR169j聚为一簇,其余11个sit-MIR169聚为另一簇。亲缘关系较近的sit-MIR169c和sit-MIR169d位于6号染色体的相邻位置,sit-MIR169h和sit-MIR169i位于2号染色体的相邻位置,因此它们可能产生于串联复制。然而亲缘关系较近的sit-MIR169a和sit-MIR169e基因并没有位于同一染色体,说明它们不是通过串联复制产生。
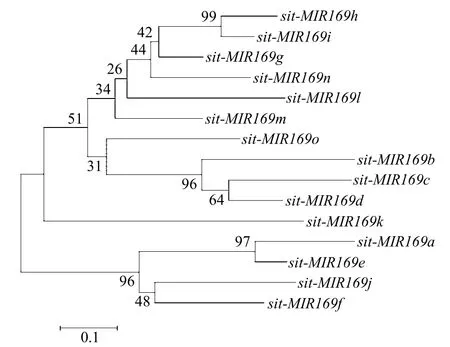
图2 sit-miR169家族的系统进化分析Fig.2 Phylogenetic analysis of sit-miR169 family
对谷子、水稻和拟南芥的miR169家族成员的前体序列进行系统进化分析,结果(图3)显示,谷子、水稻和拟南芥的miR169亲缘关系较近,尤其是谷子和水稻的部分miR169亲缘关系很近,例如sit-MIR169b/osa-MIR169b和sit-MIR169e/osa-MIR169c等。
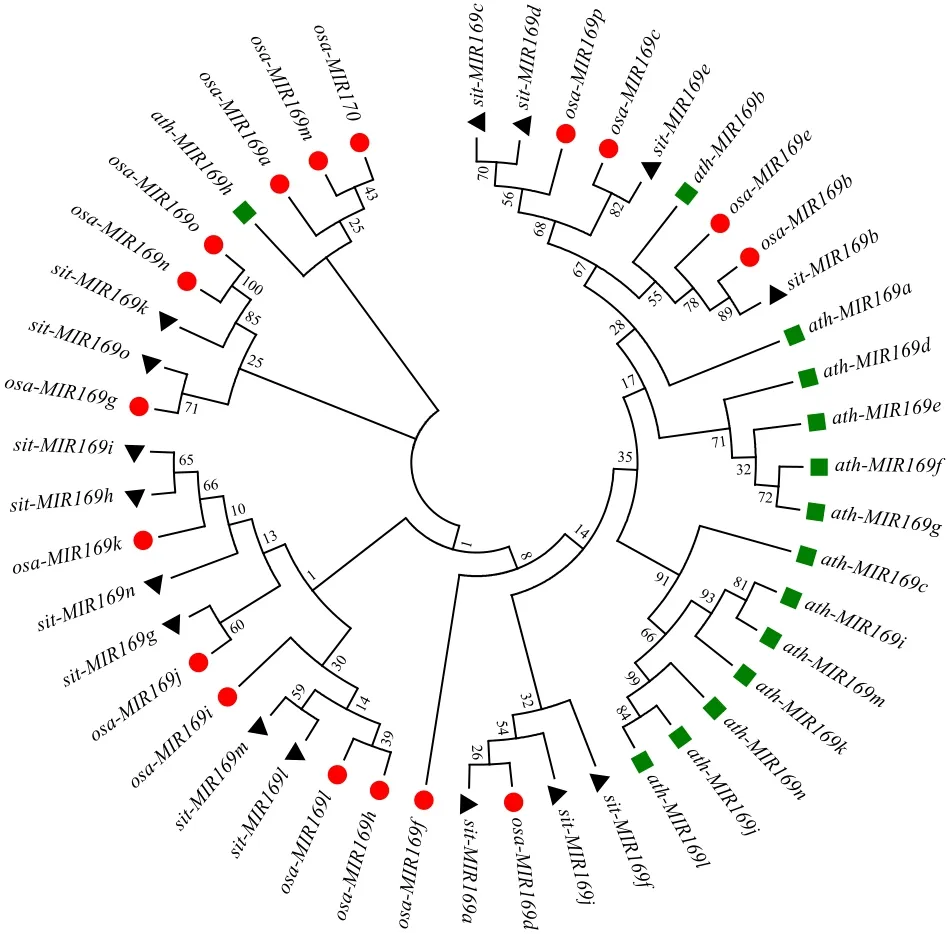
图3 谷子、水稻和拟南芥miR169家族系统进化分析Fig.3 Phylogenetic analysis of the miR169 family in foxtail millet, rice and Arabidopsis
2.3 谷子miR169序列碱基保守性
sit-miR169前体序列和成熟序列的碱基保守性如图4所示。15个sit-MIR169前体序列碱基数在88~149,前体序列1-21和104-123位点的碱基保守性较高,其他位点的保守性较低。成熟体基本产生于前体序列中保守性较高的1-21位点。除sit-miR169k和sit-miR169o成熟序列碱基数为20外,其余13个sit-miR169成熟序列碱基数均为21。15个sit-miR169成熟体中17个碱基完全保守,只有3个碱基保守性略低,说明谷子miR169在进化过程中保守性很强。星标序列中第15位碱基完全保守,第11位比较保守,其他位点的碱基保守性较差。
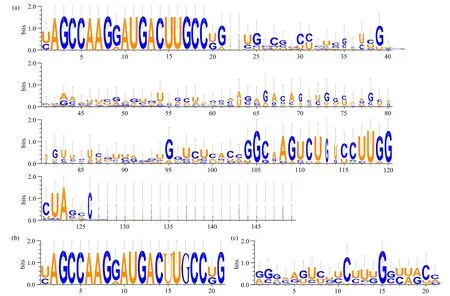
图4 sit-miR169序列碱基保守性Fig.4 Base conservativeness of sit-miR169 sequences
2.4 谷子miR169前体二级结构预测
sit-miR169a前体sit-MIR169二级结构预测结果(图5)显示,15个sit-MIR169均可形成较为稳定的二级茎环结构,且成熟体均产生于前体的5′端臂上,保守性较强。
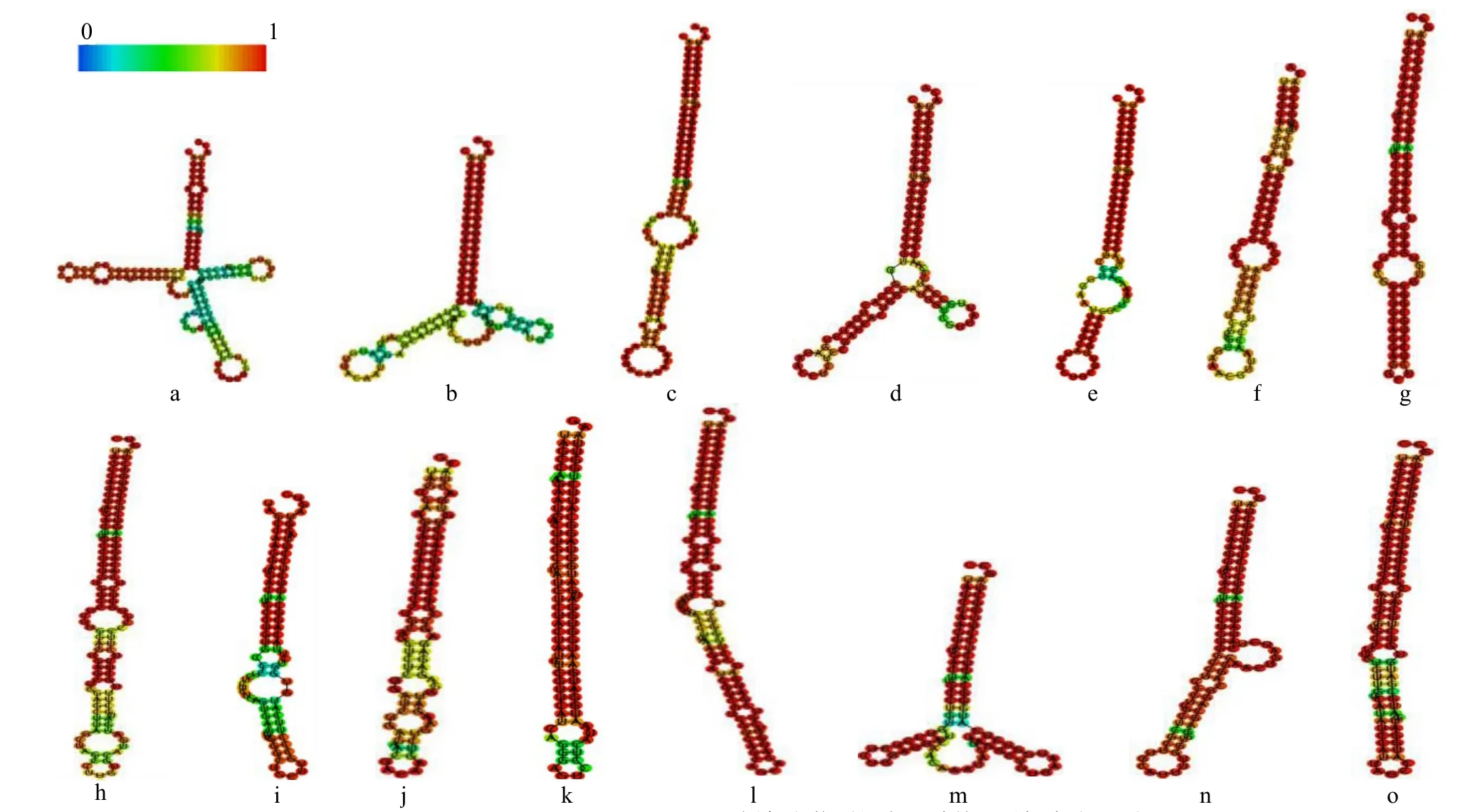
图5 sit-MIR169的二级茎环结构Fig.5 Secondary stem-loop structure of sit-MIR169
2.5 谷子miR169组织表达特异性
由图6可知,sit-miR169g、sit-miR169h、sitmiR169i、sit-miR169j、sit-miR169k、sit-miR169l、sitmiR169m、sit-miR169n和sit-miR169o主要在谷子根部表达,sit-miR169a、sit-miR169b、sit-miR169c、sit-miR169d和sit-miR169e在叶和根中表达量较高,sit-miR169f在花中优势表达。结果表明,sit-miR169广泛参与谷子各器官和组织的生长发育。
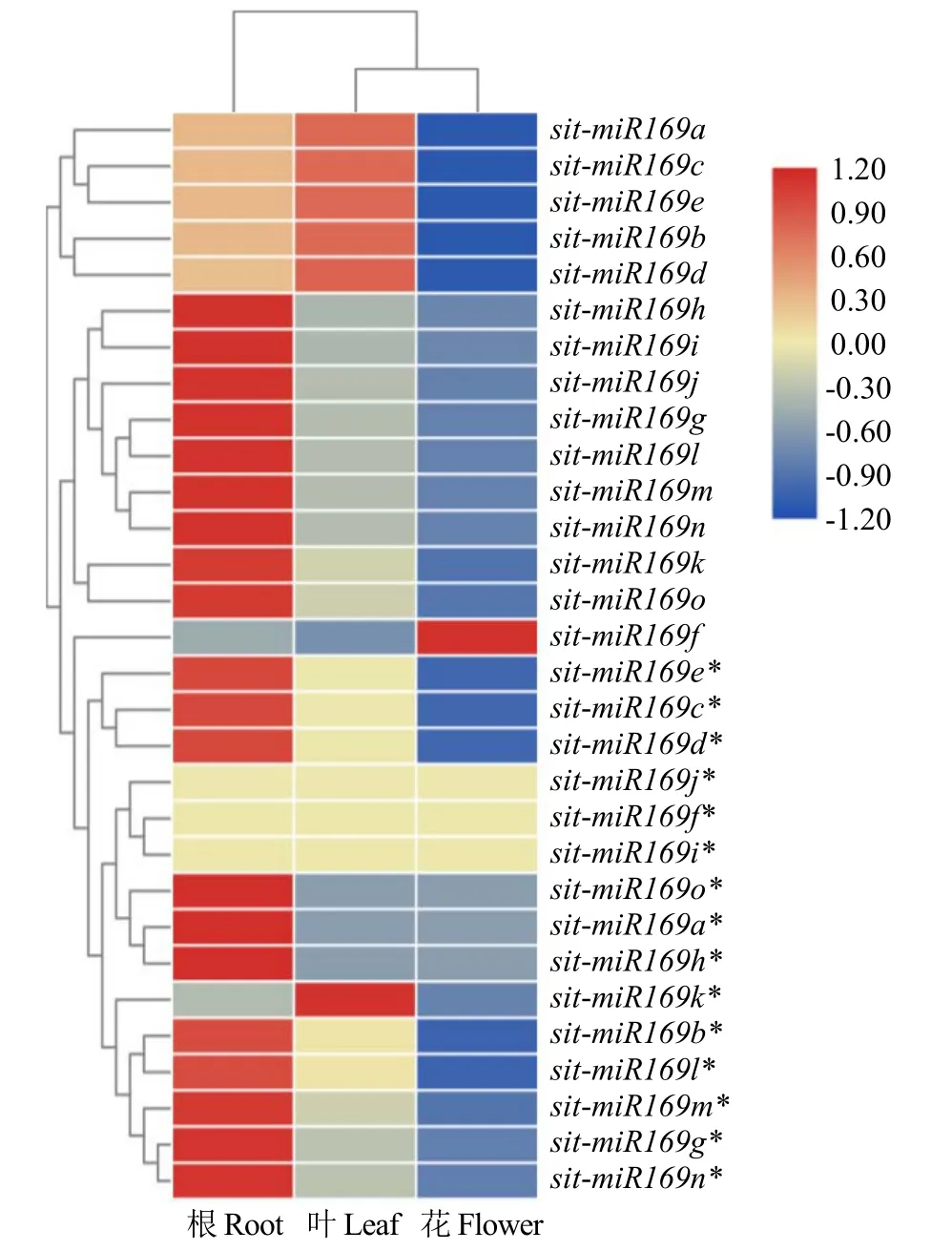
图6 sit-miR169在不同组织中的表达情况Fig.6 Expression of sit-miR169 in different tissues
2.6 谷子miR169靶基因预测
靶基因预测(表2)显示,谷子miR169靶基因编码的蛋白质大多是 NF-YA转录因子家族成员,除此之外,还有L型凝集素类受体蛋白激酶、ω-6脂肪酸去饱和酶、转运蛋白Sec1a、SDG40蛋白、环指跨膜蛋白和核糖体大亚基假尿苷合酶SVR1。NF-Y是一类结合CCAAT-box的重要转录因子,包括3个亚基NF-YA、NF-YB和NF-YC。所有sitmiR169的靶基因都有NF-YA,说明NF-YA亚基基因是sit-miR169最主要的靶基因。谷子miR169靶基因中的NF-YA家族成员有NF-YA1、NF-YA4、NFYA5、NF-YA7和NF-YA10。
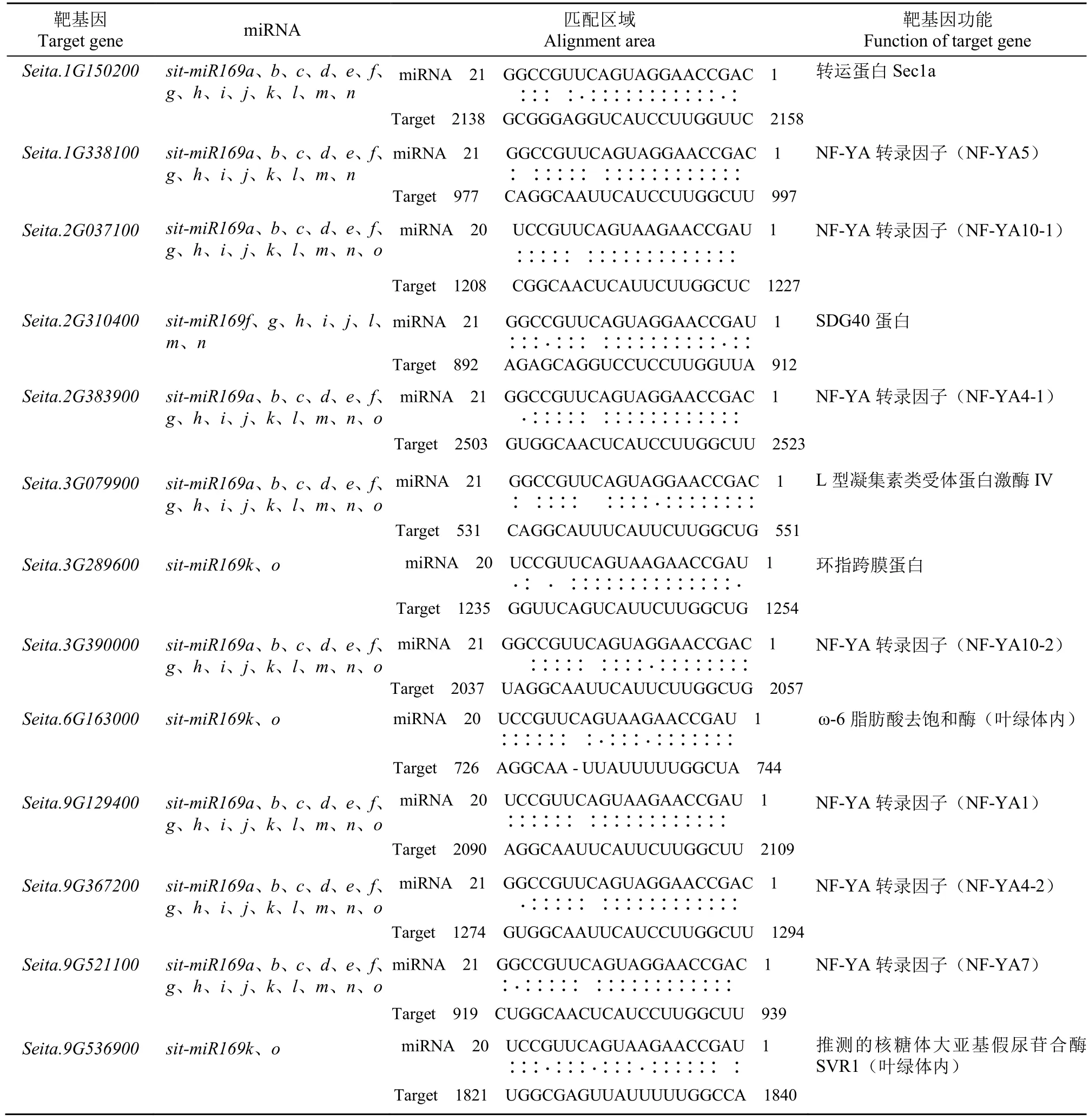
表2 谷子miR169靶基因预测Table 2 Prediction of miR169 target genes in foxtail millet
2.7 谷子miR169靶基因的组织特异性和环境诱导表达分析
miR169通过调控靶基因,尤其是NF-YA的表达量来影响植物的生长发育和对环境的应答。谷子miR169靶基因NF-YA主要在根和穗中表达(图7),表现出明显的组织表达特异性,表明其可能主要参与调控谷子根和穗部的生长发育。Seita.2G037100和Seita.2G383900在穗中优势表达,其蛋白质序列相似度为52%,推测这2个基因可能是产生于串联复制的同源基因,其编码的 NF-YA转录因子可能具有相似功能。

图7 sit-miR169靶基因在不同组织和环境诱导条件下的表达情况Fig. 7 Expression of sit-miR169 target genes in different tissues and environment conditions
基因表达数据(图7)表明,sit-miR169靶基因的表达受环境诱导,例如Seita.1G338100、Seita.3G390000、Seita.9G129400、Seita.9G367200和Seita.9G521100基因在根中都有较高表达,但在不同环境诱导条件下的根中表达量表现出显著差异。根中Seita.1G338100表达量在光照处理下最高,在氨或硝酸盐处理下次之,在尿素处理下最低;而Seita.3G390000在尿素处理下最高,在氨或硝酸盐处理下次之。Seita.9G521100在光照、尿素、氨或硝酸盐处理下根中表达量差异不显著,但在干旱条件下根中表达量较高,说明干旱能诱导其在根中穗的表达。
3 讨论
谷子是我国古老的传统抗旱、耐贫瘠作物。目前全球水资源缺乏,化肥施用过度,谷子在现代农业绿色发展中的重要作用日益突显。小米具有保健功能,受到越来越多的关注,谷子研究和小米产业迎来新的机遇。microRNA是一小段非编码单链RNA,参与复杂的基因表达调控,控制着植物的发育程序,并参与生物和非生物胁迫应答过程。miR169是植物中较为庞大和保守的miRNA家族之一。miR169在一些植物中的功能已经得到初步的研究,其能广泛参与植物的逆境胁迫响应(如盐碱、干旱胁迫及致病疫霉菌侵染)和生长发育调控(如开花调控、花和根发育)[10-19]。谷子miR169的靶基因大多是NF-YA转录因子基因,miR169通过调控靶基因,尤其是NF-YA的表达量影响植物的生长发育和环境应答反应。鉴定并分析miR169家族对解析miR169家族成员在植物生长发育及逆境胁迫响应中的功能具有重要意义[28]。
本研究表明,谷子基因组共有15个sit-miR169家族成员,系统进化分析显示,sit-miR169大致分为2簇,部分sit-MIR169成员可能产生于串联复制(如sit-MIR169c和sit-MIR169d、sit-MIR169h和sit-MIR169i)。sit-miR169成熟序列碱基保守性很高,星标序列碱基保守性较低。sit-miR169家族成员均能形成较稳定的二级茎环结构,成熟序列位于 5′端。sit-miR169广泛参与谷子各个器官和组织的生长发育。sit-miR169靶基因包括NF-YA转录因子、L型凝集素类受体蛋白激酶、ω-6脂肪酸去饱和酶、转运蛋白Sec1a、SDG40蛋白、环指跨膜蛋白和假尿苷合酶 SVR1基因等,其中最主要的是NF-YA基因。sit-miR169k和sit-miR169o分别位于3和8号染色体,在sit-miR169家族成员中只有sitmiR169k和sit-miR169o的成熟序列碱基数目是20,并且成熟序列完全相同。sit-miR169k和sit-miR169o的靶基因有环指跨膜蛋白、假尿苷合酶SVR1和ω-6脂肪酸去饱和酶基因,与其他sit-miR169家族成员截然不同,这表明二者同源性较高,可能具有相同功能。谷子NF-YA基因主要在根和穗中表达,表现出明显的组织表达特异性。Seita.2G037100和Seita.2G383900在穗中优势表达,其蛋白质序列相似度为52%,推测二者可能是产生于串联复制的同源基因,其编码的 NF-YA转录因子可能具有相似功能。sit-miR169靶基因的表达受环境诱导,如干旱条件能进一步诱导Seita.9G521100基因在根中的表达。
谷子和水稻同为单子叶禾本科二倍体作物,在进化上具有较近的亲缘关系[4],遗传图谱比较显示,谷子与水稻的基因组序列有很高的保守性[29]。本文对谷子、水稻和拟南芥miR169进行系统发育分析,结果显示,谷子和水稻的部分miR169亲缘关系很近,例如sit-MIR169b/osa-MIR169b和sit-MIR169e/osa-MIR169c等。在水稻中,miR169与盐胁迫应答、干旱诱导相关,参与负调控水稻对稻瘟病菌的免疫[11-12]。例如osa-miR169g是osa-miR169家族中唯一能被干旱诱导的成员,它的启动子上游区域有2个相邻的DRE(干旱响应元件),表明其可能直接受 CBF/DREB转录因子的调控,在水稻干旱响应中扮演重要角色[12]。谷子抗旱性和节水性突出,是研究抗旱调控机制的重要作物,然而与水稻、玉米等其他禾本科作物相比,谷子抗逆研究较为滞后,模式植物水稻miR169的研究可以为谷子提供很好的思路。
miR169与靶基因共同调控植物生长发育的各个方面,miR169与靶基因调控模块研究为miRNA功能分析提供了思路。例如miR169/NF-YA调控模块对植物的发育和逆境响应起着重要作用,miR169/NF-YA参与逆境诱导早花调控[30]。部分sit-miR169及其靶基因的组织表达部位具有一致性,sitmiR169a、sit-miR169b、sit-miR169c、sit-miR169d和sit-miR169e主要在叶片中表达,它们亲缘关系较近。它们的主要靶基因为转运蛋白、NF-YA转录因子和L型凝集素类受体蛋白激酶基因,其中L型凝集素类受体蛋白激酶基因主要在叶片中表达,这与sit-miR169a-sit-miR169e的组织表达部位一致,因此sit-miR169a-sit-miR169e可能参与叶片中L型凝集素类受体蛋白激酶基因的表达调控。sit-miR169f在花中优势表达,其靶基因Seita.2G037100(NF-YA10-1)和Seita.2G383900(NF-YA4-1)在穗中优势表达,因此sit-miR169f与Seita.2G037100、Seita.2G383900可能共同参与花发育调控。此外,所有sit-miR169靶基因都有NF-YA基因,NF-YA主要在根和穗部中表达,因此sit-miR169与NF-YA可能共同参与谷子根和穗部发育调控。除NF-YA基因外,miR169的靶基因还包括ω-6脂肪酸去饱和酶基因,植物脂肪酸组成很大程度上受到脂肪酸去饱和酶种类和数量的调控,深入了解脂肪酸去饱和酶的种类、数量及其编码基因对于定向改变植物的脂肪酸组成具有重要的理论和实际意义[31],这些靶基因的研究为miR169作用机制分析奠定了理论基础。
4 结论
在谷子全基因组水平上鉴定了15个sit-miR169,sit-MIR169前体进化树大致分为2簇,其中部分sitmiR169与水稻的miR169亲缘关系较近。sit-miR169家族成员的成熟序列碱基保守性高,星标序列碱基保守性较低。15个sit-MIR169均可形成较为稳定的二级茎环结构,成熟体产生于5′端。
sit-miR169广泛参与谷子各组织的生长发育。谷子miR169靶基因有NF-YA转录因子、L型凝集素类受体蛋白激酶、ω-6脂肪酸去饱和酶、转运蛋白Sec1a、SDG40蛋白、环指跨膜蛋白和核糖体大亚基假尿苷合酶SVR1的基因,其中NF-YA转录因子基因最多。谷子NF-YA基因主要在根和穗中表达。
猜你喜欢
当代陕西(2021年21期)2022-01-19 02:00:32
天津医科大学学报(2021年1期)2021-12-05 11:11:05
教学考试(高考生物)(2020年6期)2020-11-23 05:25:56
食品与生物技术学报(2020年8期)2020-01-06 08:00:56
科学24小时(2019年5期)2019-06-11 08:39:38
发明与创新(2019年9期)2019-03-26 02:22:48
现代检验医学杂志(2016年5期)2016-08-20 03:17:08
现代农业(2015年5期)2015-02-28 18:40:53
都市家教·上半月(2014年4期)2014-05-30 19:24:19
茶叶通讯(2014年2期)2014-02-27 07:55:40
