
基于脑胰岛素信号通路探讨中药防治代谢性疾病认知障碍研究进展*
2022-04-28 03:16方敏惠杨晓梅张保伟
中医学报 2022年1期
方敏惠,杨晓梅,张保伟,2
1.河南中医药大学,河南 郑州 450046; 2.平煤神马医疗集团总医院,河南 平顶山 467000
肥胖、2型糖尿病(type 2 diabetes mellitus,T2DM)、高血压、血脂异常等均为慢性代谢紊乱性疾病,又称为代谢性疾病。目前,代谢性疾病引起的认知障碍越来越受到人们的关注。流行病学调查和多项纵向研究表明,代谢性疾病人群患轻度认知障碍、痴呆或阿尔茨海默症(Alzheimer′s disease,AD)的风险显著增加[1-2]。长期慢性的代谢紊乱会对中枢神经系统产生负面影响,引起认知障碍等疾病,临床症状主要表现为语言理解能力、注意力、记忆力和行动力等的明显下降。代谢性疾病的病理生理改变及发病机制十分复杂,有待进一步研究。最新研究表明,大脑胰岛素信号在调节能量稳态和神经元生长、突触可塑性、认知保护等方面发挥着重要作用[3],而肥胖和T2DM能够导致大脑胰岛素信号传导异常,损害神经功能,从而导致认知障碍[4-5]。因此,调节大脑胰岛素信号通路可能是防治代谢性疾病认知功能障碍的有效途径。现代医学治疗代谢性疾病认知障碍的靶向药物作用较为单一,不良反应明显,而中药及中药方剂中的多种有效成分能够充分发挥其多靶点、多效能、多途径的作用特点,安全性较高,具有潜在的治疗优势。本文从大脑胰岛素信号通路角度对近年来中医药防治代谢性疾病认知障碍的实验研究进行总结,以期为临床诊疗提供新方法和新思路。
1 脑胰岛素信号通路的作用
胰岛素受体是一种酪氨酸激酶,广泛存在于大脑中,尤其是与记忆相关的大脑区域,如大脑皮质、海马体、下丘脑和杏仁核等[6]。胰岛素由胰岛素受体介导,通过饱和的转运系统跨血脑屏障运输,从而进入中枢神经系统[7]。大脑中的胰岛素与胰岛素受体结合,引发受体β亚基的自磷酸化,导致多种胰岛素受体底物(insulin receptor substrate,IRS)结合和酪氨酸磷酸化,随后,磷酸化的IRS激活磷脂酰肌醇3激酶/蛋白激酶B(phosphoinositide-3 kinase/protein kinase B,PI3K/Akt)、丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)、腺苷酸活化蛋白激酶(adenosine monophosphate activated protein kinase,AMPK)等通路,在大脑中发挥重要作用[8]。越来越多的证据表明,大脑胰岛素能参与调节新陈代谢与认知功能,不仅能控制进食行为、体重、糖脂代谢及能量平衡[9],还能促进神经元增殖、生长和存活,调节突触结构和功能,提高学习记忆能力[3]。因此,大脑胰岛素信号通路是中枢神经系统发挥作用的关键控制因素。相关研究表明,肥胖、糖尿病(diabetes mellitus,DM)动物模型可表现出脑胰岛素信号的传导紊乱,且存在与AD相关的脑部病理变化,如β-淀粉样蛋白(amyloid β-protein,Aβ)沉积增加、神经纤维缠结 (neurofibrillarytangles,NFT)形成和突触可塑性降低,损害认知功能,而通过鼻内注射胰岛素可有效预防患者认知能力下降和其他脑损伤[10-11]。
2 中药及其复方通过调节脑胰岛素信号通路防治代谢性疾病认知障碍的作用机制
2.1 通过调节脑胰岛素信号通路抑制Tau磷酸化及Aβ沉积Tau是一种微管相关蛋白,能促进微管蛋白组装成微管,并发挥稳定神经元微管的作用[12]。过度磷酸化的Tau最终聚集成具有毒性的NFT,影响神经细胞功能[13]。糖原合成酶激酶-3β (glycogen synthase kinase-3β,GSK-3β)是一种参与胰岛素信号传导和Tau高磷酸化的重要激酶。高糖或高脂肪饮食可抑制PI3K/Akt信号通路的激活,增加GSK-3β活性,并导致Tau蛋白的过度磷酸化,加速海马细胞凋亡,损害认知功能[14]。此外,活化的GSK-3β也参与了Aβ的沉积过程,干扰神经元的生长和存活[15]。Aβ沉积也能引起神经元凋亡级联反应,导致神经退行性变。大脑胰岛素信号受损可影响Aβ蛋白的积累与清除[16],相反,Aβ蛋白也可通过下调IRS破坏胰岛素信号传导[17]。大脑胰岛素降解酶(insulin degrading enzyme,IDE)是Aβ清除的主要蛋白酶,能被胰岛素竞争性抑制,导致Aβ降解减少。IDE作为胰岛素受体级联反应的下游靶点,可被胰岛素负反馈调节[18]。有研究报道,DM转基因小鼠脑部IDE活性下降,并伴随Aβ积累增加,而胰岛素信号通路激活剂能显著提高IDE的表达水平,减轻空间学习和认知障碍[19]。
Wang等[20]研究发现,黄芪提取物毛蕊黄酮能增强链脲佐菌素(streptozotocin,STZ)诱导的DM大鼠Akt磷酸化活性,降低GSK-3β和Tau蛋白表达水平,提高空间学习记忆能力,而经PI3K/Akt抑制剂LY294002处理后,未对这些蛋白产生影响。Li等[21]以13.40%黄连总生物碱混合0.5%羧甲基纤维素钠治疗STZ联合高脂高糖饮食(high-fat and high-fructose diet,HFFD)诱导的T2DM模型大鼠,结果发现大鼠脑内IRS、PI3K和Akt的磷酸化表达增加,并抑制GSK-3β过度激活,证明黄连总生物碱能够通过增强IRS/PI3K/Akt/GSK-3β胰岛素信号传导,减少Aβ沉积,从而改善记忆障碍。另外,许多中药单品及其提取物如小檗碱[22]、黄芩素[23]、麝香草酚[24]、人参皂苷[25]、桑白皮[26]、芍药苷[27]、豆蔻提取物[28]等均可以通过调节大脑PI3K/Akt信号通路,抑制GSK-3β活性,降低Aβ蛋白沉积和Tau蛋白磷酸化,改善神经细胞功能,提高学习和记忆能力。中药及中药复方成分复杂,疗效确切,具有多靶点、多功能的天然优势。高晓斐等[29]在转基因T2DM动物模型KK-Ay小鼠的实验中,发现醒脑益智剂能提高小鼠大脑海马内PI3K的表达水平,降低GSK-3β和Tau活性。其中,中剂量(生药 7.2 g·kg-1)复方组治疗效果最佳,能显著改善T2DM小鼠海马胰岛素信号传导通路,进而增强学习、记忆功能。已知IDE参与Aβ的降解,有学者对滋补脾阴方防治DM认知障碍大鼠机制进行研究,发现大鼠皮质p-IRS-1丝氨酸磷酸化水平减弱,p-Akt 及IDE蛋白水平增加,抑制可溶性与不可溶性Aβ1-42表达,提示滋补脾阴方可上升IDE蛋白表达,改善胰岛素信号通路,加速Aβ降解,从而减少学习记忆障碍[30]。见表1。
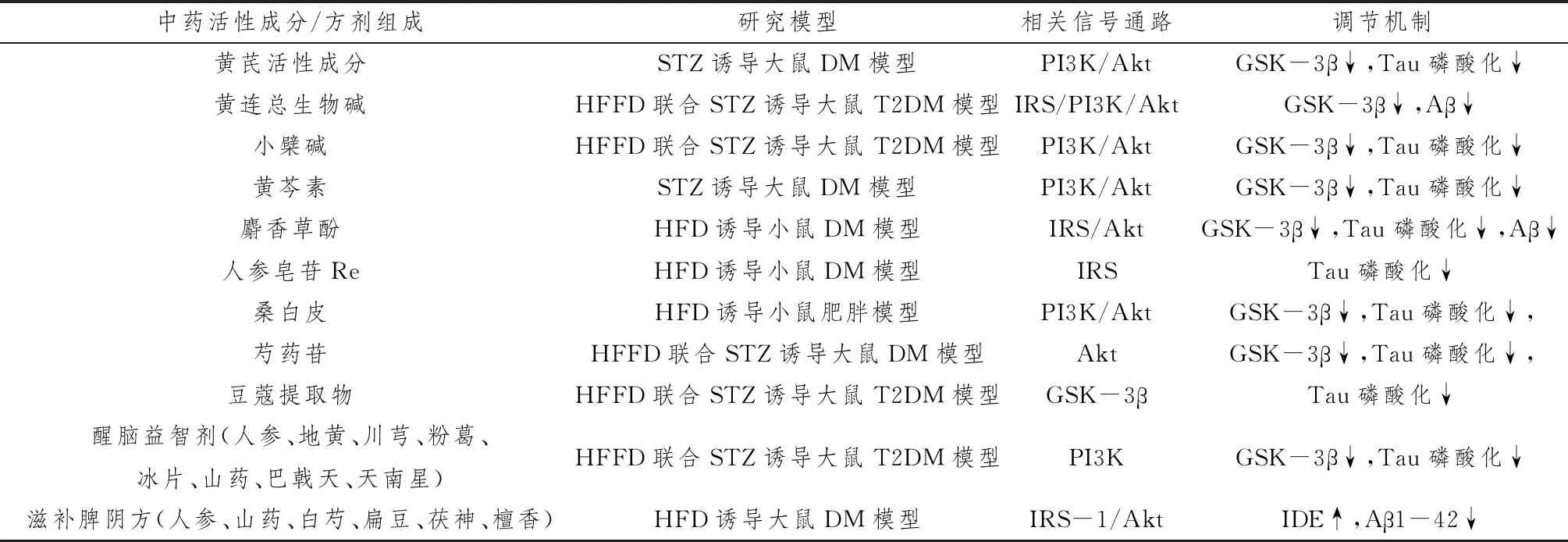
表1 中药及复方对Tau磷酸化及Aβ沉积的调节机制
2.2 通过调节脑胰岛素信号通路抑制神经细胞凋亡海马体是大脑相关记忆功能和认知功能的重要结构。海马神经元在遭受高血糖损伤时会出现凋亡细胞死亡[31]。已有研究证明,PI3K/Akt信号通路能够参与拮抗神经细胞凋亡,维持神经细胞的生存,从而影响大脑记忆形成[32]。活化的Akt能够促使促凋亡蛋白B淋巴细胞瘤-2基因相关启动子(Bad)磷酸化,导致抗凋亡蛋白突触素B细胞淋巴瘤-2(B-cell lymphoma-2,Bcl-2)的释放。该蛋白通过与促凋亡因子Bax结合,形成复合物,维持线粒体膜的完整性[33]。当Akt失活时,线粒体外膜过渡孔的异常打开可以释放细胞色素C(Cytochrome C,Cyt C),依次激活半胱氨酸天冬氨酸蛋白酶(cysteinylasparate specific proteinase,caspase)-9、caspase-3、caspase-7,最终导致细胞凋亡,损害大脑认知功能[34]。Akt失活也能激活GSK-3β,通过控制抗凋亡蛋白Bcl-2的失活来调节细胞凋亡[35]。HFFD还能够刺激JNK通路的激活,与Akt改善神经元存活的作用相反,神经元中产生的炎症因子能够刺激磷酸化c-Jun氨基末端激酶(phosphorylated c-Jun N-terminal kinase,JNK)的异常激活,导致IRS-1丝氨酸磷酸化,抑制Akt活性,增加神经细胞凋亡[36]。
Nan等[37]在体外实验中检测人参皂苷Rb1对甲基乙二醛(methylglyoxal,MGO)诱导的SH-SY5Y细胞损伤的影响及其机制,发现Rb1可以剂量依赖性促进细胞增殖,且在1.5 μmol·L-1时治疗效果最佳,能显著提高磷酸化Akt水平,上调Bcl-2/Bax比值,抑制caspase-3和caspase-9的表达,发挥细胞保护作用。此外,SH-SY5Y细胞加入Rb1 1 h前,用10 μmol·L-1的PI3K抑制剂进行预处理,明显阻断了Rb1逆转细胞活力和细胞凋亡作用,说明Rb1可通过激活PI3K/Akt信号通路,从而保护细胞免受MGO诱导的凋亡损伤。杨利[38]以300 mg·kg-1的知母总皂苷治疗STZ联合HF诱导的DM大鼠,结果显示,与模型组比较,治疗组大鼠海马JNK、Bax表达水平降低,Bcl-2表达升高,表明知母总皂苷能通过抑制JNK信号影响下游促凋亡通路的过度激活,从而改善认知和记忆形成。另有实验研究显示,牛磺酸[39]、淫羊藿苷、槲皮素[40]、桑白皮[26]、知母-黄柏药对[41]及中药复方涤痰汤[42]等均可以激活动物模型海马PI3K/Akt信号通路,提高Bcl-2/Bax比值或下调Bad、caspase-3蛋白表达,抑制神经细胞凋亡,改善DM或肥胖诱导的认知障碍。见表2。
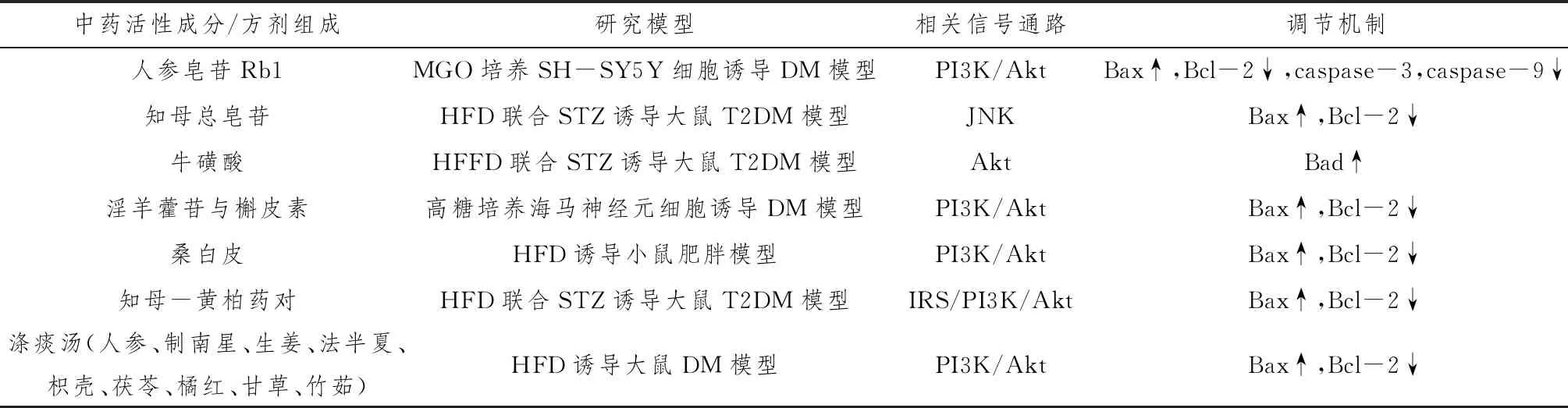
表2 中药及复方对神经细胞凋亡的调节机制
2.3 通过调节脑胰岛素信号通路促进神经元再生和突触可塑性研究显示,DM大鼠模型认知受损与海马神经元活性降低、突触可塑性减少有关[43]。脑源性神经营养因子(brain derived neurotrophic factor,BDNF)是大脑中最丰富的神经营养素,能促进神经元发生、神经保护和突触可塑性,以及海马体和额叶皮层的记忆[44-45]。胰岛素信号传导通过介导PI3K/Akt途径和下游信号传导控制大脑中神经元的存活和突触可塑性。RAS/ERK及IRS下游信号PI3K/Akt通过刺激cAMP应答元件结合蛋白(response element binding protein,CREB)的磷酸化,在调节BDNF转录中发挥着关键作用[35],增加突触可塑性蛋白,促进神经元再生。在学习记忆与认知功能方面发挥着重要作用的胰岛素样生长因子1(insulin like factor1,IGF-1),作为PI3K/Akt途径的上游调节信号,同样参与了调节神经元生长与突触的形成[46]。
胰岛素信号的激活与ERK/CREB/BDNF通路相关,可影响大脑中多种形式的学习和记忆。Mi等[47]在食物中补充2 g·L-1没食子儿茶素-3-没食子酸酯(Epigallocatechin-3-gallate,EGCG)治疗肥胖导致的认知缺陷,结果发现,EGCG可显著增加重组N-乙基马来酰亚胺敏感因子(N-ethylmaleimide-sensitive factor,Nsf)和突触结合蛋白-1(synaptotagmin-1,Syt-1),上调IRS-1、PTP1B及Akt酪氨酸磷酸化。此外,与模型组相比,EGCG可改善小鼠脑内ERK失活,逆转BDNF、神经营养因子-3/4(neurotrophin-3/4,NT-3/4)和神经生长因子(nerve growth factor,NGF)表达的下调,表明EGCG能通过刺激IRS-1/Akt活化和减少PTP1B表达使胰岛素信号正常化,随后介导ERK/CREB/BDNF信号,保护突触功能,有效预防小鼠的记忆丧失。体外研究发现,积雪草苷抑制了高糖诱导的 SH-SY5H 细胞中Akt丝氨酸磷酸化,且在 0.1 μmol·L-1、1 μmol·L-1浓度时显著上调了突触后密度蛋白95(postsynaptic density proteins 95,PSD-95)和突触素(synapsin,SYN)表达,提示积雪草可以通过调节PI3K/Akt通路促进突触蛋白表达,恢复DM认知障碍[48]。T2DM在长期慢性高血糖情况下易造成血脑屏障损伤,影响紧密连接蛋白信号传递。袁有才[49]使用免疫印迹法(Westen-blot,WB)检测发现,中、小剂量新加葛根芩连汤增加了STZ联合HFFD致T2DM小鼠闭锁蛋白、封闭蛋白及胞浆黏附蛋白(zonula occluden l,ZO-1)的表达,有效减少连接黏附分子(junctional adhesive molecule1,JAM-1)蛋白表达,保护血脑屏障和脑微循环,改善神经元代谢异常。在进一步的机制研究中发现,与对照组比较,中、小剂量组IGF-1、PI3K、Akt和CREB磷酸化表达显著提高,提示新加葛根芩连汤能够有效改善IGF-1和PI3K/Akt/CREB信号通路,促进神经元再生、突触形成及学习记忆。另有相关研究表明,五甲基槲皮素[50]、青蒿素[51]、芍药苷[52]、沙棘黄酮[53]、加味生脉散[54]等中药提取物及中药方剂均能通过改善胰岛素信号通路,上调突触可塑性相关蛋白表达,促进神经元再生,从而提高代谢性疾病患者的学习记忆与认知能力。见表3。

表3 中药及复方对神经元再生和突触可塑性的调节机制
2.4 通过调节脑胰岛素信号通路改善大脑葡萄糖代谢大脑中海马体记忆的形成受到葡萄糖代谢和胰岛素信号的限制。葡萄糖转运蛋白4(glucose transporter 4,GLUT4)是一种胰岛素敏感性葡萄糖转运体,存在于参与学习和情景记忆的特定区域,尤其是海马体的大脑神经元中[55-56]。神经元葡萄糖代谢异常被认为是由胰岛素信号转导水平方面的代谢通路障碍引起的。大脑胰岛素与胰岛素受体结合,通过刺激PI3K/Akt、AMPK信号通路,影响GLUT3、GLUT4的表达以及从胞浆到细胞膜的转运,从而促进大脑葡萄糖代谢和细胞存活[35],增强海马体记忆过程。研究表明,高脂饮食(high fat diet,HFD)诱导的DM模型大鼠海马神经元细胞溶质GLUT4蛋白水平[57]和p-Akt介导的GLUT4膜转位水平的降低,可导致模型大鼠空间记忆能力下降[32]。
You等[26]在HFD致肥胖小鼠研究中发现,100 mg·kg-1或200 mg·kg-1桑白皮均可以提高IRS、PI3K和Akt酪氨酸磷酸化活性,增加GLUT4蛋白表达水平,促进大脑葡萄糖正常代谢过程,发挥调节认知和行为的作用。Lakshmanan等[58]研究姜黄素对高糖联合STZ介导大鼠DM认知损伤的作用,结果表明姜黄素能显著降低Tak1、AMPKα1蛋白磷酸化,下调GLUT4蛋白表达,说明姜黄素能通过调节AMPK信号通路,减少DM大鼠大脑中的葡萄糖转位,保护大脑认知功能。有研究报道,在星形胶质细胞中,GSK-3β对糖原合成的主要限速酶的活性有负反馈调节作用。杨蕙等[59]研究发现,左归降糖解郁方能提高DM模型海马星形胶质细胞中PI3K、Akt磷酸化水平,降低GSK-3β活性,增加GLUT3表达,从而调节神经元细胞的葡萄糖摄取,改善模型大鼠认知功能。还有报告显示,荨麻提取物[60]、沙棘黄酮[53]、槲皮素[61]等均能恢复DM诱导的胰岛素信号障碍,上调GLUT4表达水平,保护神经元的葡萄糖摄取功能,改善认知障碍。见表4。

表4 中药及复方对葡萄糖代谢的调节机制
2.5 通过调节脑胰岛素信号通路减缓氧化应激氧化应激是氧化自由基和相关活性氧(reactive oxygen species,ROS)过量产生的结果。有研究证明,氧化应激会损害肥胖、DM胰岛素信号转导与认知能力[62]。ROS可与脑组织中丰富的脂质结合,生成过氧化氢自由基,诱导神经细胞凋亡,参与认知缺陷的发生和发展[63]。核因子E2相关因子2(nuclear factor E2-related factor 2,Nrf2)是维持内源性抗氧化系统氧化还原状态稳态的关键介质。一旦激活,Nrf2从细胞质转位到细胞核并与抗氧化反应元件(anti-oxidative response element,AREARE)结合,然后启动抗氧化酶基因的转录,包括超氧化物歧化酶(superoxide dismutase,SOD)、谷胱甘肽(γ-glutamyl-l-cysteinylglycine,GSH)、烟酰胺腺嘌呤二核苷酸磷酸(nicotinamide adenine dinucleotide phosphate,NADPH)和血红素加氧酶 1(heme oxygenase-1,HO-1)等[64]。PI3K/Akt通路的激活可诱导Nrf2向细胞核移位,并增加海马神经元中HO-1的表达,保护认知功能[65]。除了PI3K/Akt信号通路外,AMPK也参与了胰岛素信号传递。最近的研究表明,AMPK可以通过激活Akt信号[66]与靶向p38丝裂原活化蛋白激酶促进Nrf2[67]核转位,改善氧化还原平衡,保护神经元细胞。
HO-1是一种由氧化应激反应产生的内源性细胞保护酶,具有清除ROS和抗氧化应激的潜在特性。Zhai等[68]体内、外研究发现,以三七皂苷(notoginsenoside R1,NGR1)处理高糖刺激HT22细胞和db/db小鼠均可提高Akt活性、Nrf2核转位水平及相关抗氧化酶HO-1、SOD表达,降低MDA、蛋白羰基的mRNA含量,下调神经元caspase-3活性和乳酸脱氢酶(lactate dehydrogenase,LDH)释放,且采用PI3K抑制剂LY294002可抑制NGR1的抗氧化和神经元保护作用,表明NGR1可能是通过激活Akt/Nrf2/HO-1通路而改善胰岛素信号,达到抑制氧化应激和保护神经元功能的目的,从而减轻认知障碍。NADPH氧化酶是各种刺激下产生超氧自由基的主要来源,其催化亚单元主要包括gp91phox、调节子单元(p22phox、p47phox和p67phox)以及Rac1,慢性高血糖诱导的氧化应激可通过激活NADPH氧化酶亚单位产生线粒体ROS。在STZ诱导的DM大鼠模型研究中发现[58],姜黄素降低了大鼠大脑MDA水平,p67phox、gp91phox、3-NT和caspase-12的表达水平明显恢复正常化,同时下调了转化生长因子β活化激酶(transforming growth factor β activated kinase,Tak1)和AMPKα1蛋白表达。AMPK可以介导代谢变化以抵抗ROS的积累,说明姜黄素可能作为一种强抗氧化剂,维持氧化还原平衡,促进神经细胞存活。中药实验研究证明,人参皂苷[37]、麝香草酚[24]等均能调节胰岛素信号传导通路,上调海马组织抗氧化基因的表达,减少ROS产生,保护神经元,从而改善DM认知障碍。见表5。

表5 中药活性成分对氧化应激的调节机制
3 中药及复方通过其他途径改善脑胰岛素信号通路作用机制
3.1 通过抑制神经炎症反应改善脑胰岛素信号通路肥胖与DM作为一种炎症性疾病,能引起包括大脑在内的全身炎症反应。小胶质细胞作为大脑炎症过程的主要效应器,在高血糖与高脂肪状态下被迅速激活,触发核因子-κB(nuclear factor kappa-B,NF-κB)、JNK等多潜能信号转导级联,并释放多种炎症因子,如肿瘤坏死因子(tumor necrosis factor,TNF)、白细胞介素6(interleukin 6,IL-6)和白细胞介素1(interleukin 1,IL-1),抑制IRS-1酪氨酸磷酸化,阻断下游效应物如PI3K/Akt级联,扰乱大脑胰岛素途径[48,69]。而胰岛素途径对中枢神经系统突触可塑性功能、认知过程和记忆能力具有重要的保护作用[70]。研究证实,在胰岛素信号失活和认知功能受损的DM小鼠中,大脑JNK和NF-κB炎症调节信号通路上调[71-72]。细胞因子信号转导抑制因子2(supressors of cytokine signaling 2,SOCS2)也能调节小胶质细胞的炎症信号[73],并通过与IRS-1的特定磷酸化残基结合,成为公认的胰岛素信号通路的潜在抑制剂[74]。当SOCS2被抑制时,中枢炎症反应引起的异常胰岛素信号可被逆转。值得注意的是,受损的脑胰岛素信号通路会失去对炎症反应的控制,从而导致更多的炎性细胞因子释放。
Mulati等[53]对HFFD致肥胖小鼠补充0.06%或0.31%的沙棘黄酮类化合物,结果显示其能显著抑制p38和NF-κB的磷酸化及其下游炎症介质/细胞因子,如诱导型一氧化氮合酶(inducible nitric oxide synthase,iNOS)、IL-1β和环氧化酶 2(cyclooxygenase2,COX-2)表达。Sun等[27]体内研究发现,STZ诱导DM大鼠中海马IL-1β和TNF-α含量显著升高,与4 mg·kg-1的罗格列酮作用相似,应用15.30 mg·kg-1芍药苷可消除大鼠炎症细胞因子IL-1β和TNF-α,使SOCS2的过度表达得到有效抑制,并增加了IRS-1的活性及Akt-Ser473和GSK-3βSer9的磷酸化水平,减少Tau过度磷酸化。说明芍药苷具有较强的抗炎作用,能通过调节SOCS2/IRS-1信号传导,恢复胰岛素信号传导,阻断Tau高磷酸化,减轻DM引起的认知功能下降。与上述作用机制不同,毛小元等[75]研究蛇床子素对DM认知障碍的抗炎作用,发现1 mg·kg-1、10 mg·kg-1、50 mg·kg-1蛇床子素均可明显抑制NF-κB p65、TNF-α 和 IL-1β活性。蛋白质免疫印迹(Western Blot,WB )检测显示,治疗组大鼠海马中 p-Akt 蛋白表达减少,使用PI3K阻断剂后,蛇床子素抗炎作用更为显著,且进一步下调了p-Akt蛋白表达,提示蛇床子素对抗DM认知障碍炎症反应的机制可能是通过抑制PI3K/Akt 信号通路实现的。此外,实验研究显示,EGCG[47]、积雪草苷[48]、山茱萸有效成分[71]、黄连温胆汤[76]等均可显著降低大脑神经炎症因子的释放,减轻炎症反应,改善胰岛素信号,进而保护认知功能。见表6。

表6 中药及复方对神经炎症反应的调节机制
3.2 通过调节内质网应激改善脑胰岛素信号通路内质网应激(endoplasmic reticulum stress,ERS)是由未折叠或错误折叠的蛋白质于内质网腔内积聚引起的,对海马神经元细胞的生存与凋亡具有重要的调节作用,可参与认知学习过程[77]。抑制物阻抗性酯酶1(inositol requiring enzyme1,IRE1)是内质网的重要感觉元件和主要损伤途径起始因子。研究报道,在ERS持续刺激情况下,未折叠蛋白反应(unfolded proteinresponse,UPR)介导的适应性反应不足以恢复正常的细胞功能,使IRE1α自身磷酸化,并激活下游凋亡信号调节激酶1(apoptosis signal-regulating kinase 1,ASK1)。活化的ASK1通过增加其转录诱导JNK表达,抑制IRS-1酪氨酸磷酸化,进一步引起DM海马胰岛素信号通路障碍与神经细胞凋亡,促进认知损害[78-79]。自噬是细胞内的一个降解过程,通过选择性清除受损、老化的细胞器或错误折叠的蛋白质来维持神经元稳态[80-81]。自噬减弱则清除内质网蛋白能力下降,导致ERS发生,加速上述过程的发生。
梁丽娜[30]在HFD联合STZ诱导的DM大鼠体内发现,滋补脾阴方治疗能够减弱大鼠皮质、下丘脑内质网应激标志分子IRE1α丝氨酸磷酸化及其下游效应分子JNK1、JNK2活性,同时增加LC3-Ⅱ、IRS-1及Akt表达。LC3-Ⅱ是自噬体膜上的标志性蛋白质,其蛋白水平一定程度上反应自噬水平的高低,说明滋补脾阴方能够纠正DM大鼠脑胰岛素信号转导障碍,提高学习记忆的作用机制,可能与其加强自噬、减轻ERS、抑制JNK活性有关。Chang等[82]体内研究发现,金枝达方能降低T2DM大鼠海马中IRE-1表达,抑制JNK、IRS-1(ser307)磷酸化,增加Akt磷酸化,促进海马胰岛素信号转导的恢复,改善DM认知功能。见表7。

表7 中药复方对内质网应激及自噬的调节机制
4 总结与展望
综上所述,代谢性疾病认知障碍与脑胰岛素信号通路有着密切的联系,脑胰岛素信号通路异常导致的Tau磷酸化、神经细胞凋亡、神经元损伤、突触可塑性减少、糖代谢异常、氧化应激等因素相互作用,影响大脑认知功能。现代医学治疗代谢性疾病认知障碍以单纯的控制血糖为主,未能达到理想的治疗效果。中医注重整体观念,强调从整体出发阐述病机和治疗疾病,结合中医学治未病思想,防治疾病及其并发症具有较大的优势。研究表明,中药生物活性成分及复方作用于代谢性疾病后,可通过直接激活脑胰岛素信号通路,抑制Tau磷酸化及Aβ沉积,减少神经细胞凋亡,促进神经元再生与突触可塑性,改善大脑葡萄糖代谢,减缓氧化应激,有效防治代谢性疾病认知障碍。同时,通过抑制代谢性疾病诱导的中枢神经炎症及调节内质网应激,可间接改善脑胰岛素信号通路,恢复代谢性疾病患者的大脑学习记忆与认知功能。中药与脑胰岛素信号通路相互作用,充分发挥了中医药多靶点、多效能、多途径的特性,使代谢性疾病认知障碍得到了有效改善。但由于大脑结构、功能的复杂性和脑胰岛素信号通路的多样性,中药调节脑胰岛素信号通路、缓解认知障碍的作用机制仍有待进一步阐明。随着研究方法的发展,可借助蛋白组学技术和网络药理学方法寻找新的中药治疗靶点。在中医整体观念和辨证论治理论的指导下,中药复方的组方运用有效地提高了代谢性疾病的认知功能,而现有文献研究主要集中在中药单体及活性成分,对中药复方的研究相对较少,需进一步研发以脑胰岛素信号通路为靶点的中药复方新药,为代谢性疾病认知障碍的治疗提供新思路、新方法。此外,中药干预脑胰岛素信号的临床研究尚不充足,不同中药复方以及同一复方不同剂量中药的随机对照临床研究也值得探索。此外,肠道菌群的改变可影响代谢性疾病认知障碍的发展,从肠道菌群角度深入探究中药及其复方治疗代谢性疾病认知障碍的作用机制,或将成为新的研究方向。
猜你喜欢
中国现代医生(2022年19期)2022-11-04
按摩与康复医学(2022年19期)2022-09-27
中国现代医生(2022年19期)2022-08-25
祝您健康(2019年8期)2019-08-09
环球时报(2019-06-21)2019-06-21
中国医药导报(2019年7期)2019-05-13
养生大世界(2018年10期)2018-10-18
科教导刊·电子版(2018年9期)2018-06-07
分析化学(2018年2期)2018-03-02
分析化学(2017年12期)2017-12-25
