
围封对藏北高寒草原土壤矿质元素和群落特征的影响
2022-04-26 03:01郭晨睿杨敬坡李少伟马娇林武建双
草业科学 2022年4期
郭晨睿,杨敬坡,李少伟,牛 犇,马娇林,武建双
(1. 河北科技大学环境科学与工程学院, 河北 石家庄 050018;2. 中国农业科学院农业环境与可持续发展研究所, 北京 100081;3. 中国科学院地理科学与资源研究所, 北京 100101;4. 西藏自治区畜牧总站, 西藏 拉萨 850000)
草地是青藏高原面积最大的生态系统。受气候变化和过度放牧双重影响,近年来青藏高原高寒草地严重退化[1]。2003年我国启动了“退牧还草”工程[2],广泛建设网围栏来排除放牧干扰、促进退化草地自我恢复[3-5]。关于围封年限对植物群落结构和功能的影响存在很大争议。有研究认为,短期禁牧封育可以改善草地群落组成,提高植被覆盖度和地上生物量[6-7],改善土壤理化性质以及土壤微生物群落[8-10]。例如,毛绍娟等[11]发现围栏封育可以提高藏北高寒草原禾草、莎草等可食牧草生物量,降低杂类草优势度。也有研究认为,尽管长期封育可提高植物物种多样性[12],但不利于群落生产力的持续改善[13-14]。当前研究对围栏封育是否显著影响土壤养分含量也存有一定争议。有研究表明,短期围栏封育(2年)能显著提高青藏高原高寒草甸表层土壤磷含量,但对全碳含量无影响[15]。> 10年长期围栏封育可显著改善青藏高寒草甸土壤碳氮含量[16-17]。但尹亚丽等[18]在三江源高寒草甸发现,即使10年围栏封育也不能明显改变土壤有机碳、全氮、全磷含量。在藏北高原仅有少量研究,Lu等[19]发现短期围栏封育对藏西北高寒草地0 - 30 cm土壤有机碳、速效氮和速效磷含量无显著影响,但可能降低土壤表层(0 - 15 cm)总氮和总磷含量。因此,探讨围栏封育对藏北草地土壤的恢复效果需要进行更全面、更深入地研究。
矿质元素是土壤养分重要组成部分,对维持植物正常的生理功能具有重要作用。例如,镁和锌是叶绿素的组成成分;铁、铜、锰等也会影响叶绿素的合成[20]。另外,作为酶的组成成分,某些矿质元素会影响植物体内氧化还原反应及大分子的合成。例如,铜、锰和锌在植物氧化还原反应中起催化作用;镁和锌元素可以影响蛋白质合成[21-22]。此外,矿质元素对于增强植物抗逆性有积极作用。适宜浓度的外源钙和外源锌能够提高植物抗氧化酶活性进而提高植物的抗寒性[23-24]。但对围栏封育是否影响和如何影响草原土壤矿质元素含量的研究较少。张建鹏等[25]发现围封 13 年会显著增加沙化草地0 - 20 cm土层铜、锰和锌的含量,降低10 - 20 cm 土层铁的含量。Radujkovic等[26]对全球草地生物量和土壤养分进行了分析,发现矿质微量元素对草原生产力存在潜在重要影响。因此,针对藏西北干旱气候区围栏封育管理下土壤矿质元素动态进行研究有利于更全面了解围封对草地生态功能的恢复机制。
截至2014年,西藏自治区共有2.96亿亩(约1.97 × 107hm2)草原发生不同程度的退化,其中68%集中分布在藏北羌塘那曲市和阿里地区[27]。2003-2012年期间,西藏共建设围栏8 641万亩(约5.76 ×106hm2)[28],其中藏北地区共有围栏4 981万亩(约3.32 × 106hm2),主要集中在紫花针茅(Stipa purpurea)草原区。高寒草原是藏北地区主要草地类型,约占该地区总面积45%[29]。因此,本研究以围栏封育和自由放牧样地为研究对象,通过连续4年野外实地采样与分析,探究围栏封育对高寒草原土壤矿质元素含量是否有显著影响;围栏封育对高寒草原地上生物量及物种多样性是否有显著影响;土壤矿质元素与地上生物量和物种多样性是否存在显著关系。
1 材料与方法
1.1 研究区域概况
样地位于西藏那曲市班戈县(88°56′ ~ 91°18′ E,29°55′ ~ 32°15′ N) 北拉镇和普保镇。班戈县共有草原3 630.93万亩(约2.42 × 106hm2),其中紫花针茅类草原846.55万亩(约5.64 × 105hm2),平均海拔为4 750 m,地势西高南低。气候属高原亚寒带半干旱季风气候,年平均气温0 ℃左右,年平均降水量介于300~450 mm,主要集中在6月 - 9月。土壤类型为高山草原土,植被群落优势物种为紫花针茅,伴生伊凡苔草(Carex ivanovae)、羊茅(Festuca ovina)、早熟禾(Poa annua)等[30]。样地气温与降水数据源于国家气象科学数据中心(表1)。

表1 研究区域概况Table 1 Information of study area
1.2 试验设计与样品采集
所选样地围栏在2006年建成。2010-2013年7月下旬或8月上旬,在无雨无雪的天气条件下对样地围栏内外进行取样。在围栏内外随机抛出30次0.1 m2样圈,记录植物物种名称和出现次数,用以计算各物种相对频度。在北拉镇和普保镇各选择一对围栏封育和自由放牧样地,在围栏内外分别随机设置5个间隔不小于20 m的0.5 m × 0.5 m样方,共计5个重复。分种记录样方内各植物名称、盖度、高度。分物种齐地面剪取植物地上部分,装入信封保存并带回。放牧引起的土壤理化性质变化主要发生在土壤表层[31],这一层次是高寒草原植物根系主要分布区[32],因此本研究土壤取样深度定为0 - 10 cm。在每个样方的东、南、西、北以及中心点用直径3 cm的土钻采集土壤样品,并进行充分混合,去除肉眼可见的碎石、根系和凋落物等杂质,装入土布袋并带回实验室风干后进行元素分析。
1.3 指标测定
地上生物量样品在65 ℃烘干至恒重并称重,精确至0.001 g,分物种记录干重,各样方地上生物量为各物种干重之和,5个样方地上生物量平均值为样地地上生物量。本研究通过样方内各物种相对盖度(relative coverage, Cr)、相对高度(relative height,Hr)和相对频度(relative frequency, Fr)计算Shannon-Wiener 指数。公式如下:
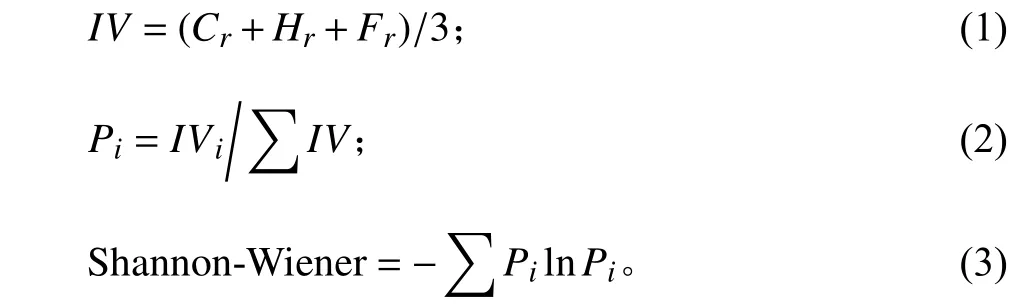
式中:IV为物种重要值,Pi为第i个物种的相对重要值。
土壤风干样品过2 mm筛去除杂质后,使用研磨仪(MM400-德国RETSCH)将样品粉碎研磨。矿质常量元素(Ca和Mg)的测定使用醋酸铵溶液作为提取液,矿质微量元素(Cu、Mn、Fe和Zn)使用二乙烯三胺五乙酸(DTPA)萃取,采用微波密闭消解电感耦合等离子发射光谱法(ICP-AES)对矿质微量和常量元素的含量进行测定[33]。
1.4 数据分析
采用双因素方差(Two-way ANOVA)分析检验各矿质元素含量、地上生物量及Shannon-Wiener 指数在不同年份间及围栏内外的差异。采用单因素方差分析比较同一样地围栏内外土壤矿质元素含量的差异。在方差分析前,对所有数据进行正态性检验(Shapiro-Wilk检验)和方差齐性检验(Bartlett检验),对不满足正态性检验的数据取对数或平方根进行转换。采用一般线性混合模型,将草地利用方式(围封或放牧)、样地位置、采样年份作为影响因子,通过方差分析量化三者主效应及交互作用对各矿质元素含量、植物物种多样性和群落地上生物量的相对影响。采用Spearman 相关系数法探究气候因子、群落特征和土壤各矿质元素间的相关性。利用双变量回归分析探究围栏内外地上生物量和物种多样性随气候因子和土壤矿质元素含量的变化规律。最后,采用协方差分析(ANCOVA)检验围封措施和土壤矿质元素对地上生物量和物种多样性的影响,固定因素为围封(放牧)处理,协变量为土壤矿质元素。所有数值均采用平均值 ± 标准误来表示。数据分析和图表制作均采用R 4.0.3[34]。
2 结果
2.1 围栏内外土壤矿质元素含量的年际变化
在矿质常量元素中,仅普保镇围栏内外土壤Ca含量在各采样年份间变化规律一致(图1);北拉镇样地围栏内外土壤Ca含量的差异逐年减小。北拉镇样地2010年和2011年围栏外土壤Ca和Mg含量显著高于围栏内。在矿质微量元素中,围栏内外北拉镇的Zn含量和普保镇的Mn含量随年份增减规律完全一致(图2)。北拉镇围栏内外Cu含量在前3年保持稳定,但在第4年变化趋势完全相反且差异显著(P< 0.05),其他样地围栏内外微量元素含量大致呈先减小后增加的趋势。同一元素围栏内外含量差异的年际变化在样地间不一致。

图1 各采样年份矿质常量元素含量Figure 1 Mineral macronutrient contents in each sampling year

图2 各采样年份矿质微量元素含量Figure 2 Mineral trace element contents in each sampling year
与放牧地相比,围栏封育对土壤Ca、Cu和Mg含量的影响显著(P< 0.05) (表2),其中围栏封育在北拉镇对Ca、Mg和Cu的含量有显著或极显著的影响(分别降低了57.1%、24.1%和37.4%),围栏封育对普保镇Cu含量有显著影响(增加了23.5%) (图3)。围栏 × 样地及围栏 × 年份对土壤Ca、Cu和Mg含量有显著的影响(P< 0.05) (表2),围栏、样地和年份三者的交互作用对土壤Ca、Cu、Fe和Mn含量的影响显著(P< 0.05)。仅土壤Cu含量在不同样地间差异显著(P< 0.05),仅土壤Mn含量在不同采样年份间差异显著(P< 0.05);任何一项因素对Zn无显著影响(P> 0.05)。围栏内外表层土壤中,Fe含量最高,Cu含量最低,各元素含量从高到低排序为Fe > Ca >Mg > Mn > Zn > Cu (图3)。
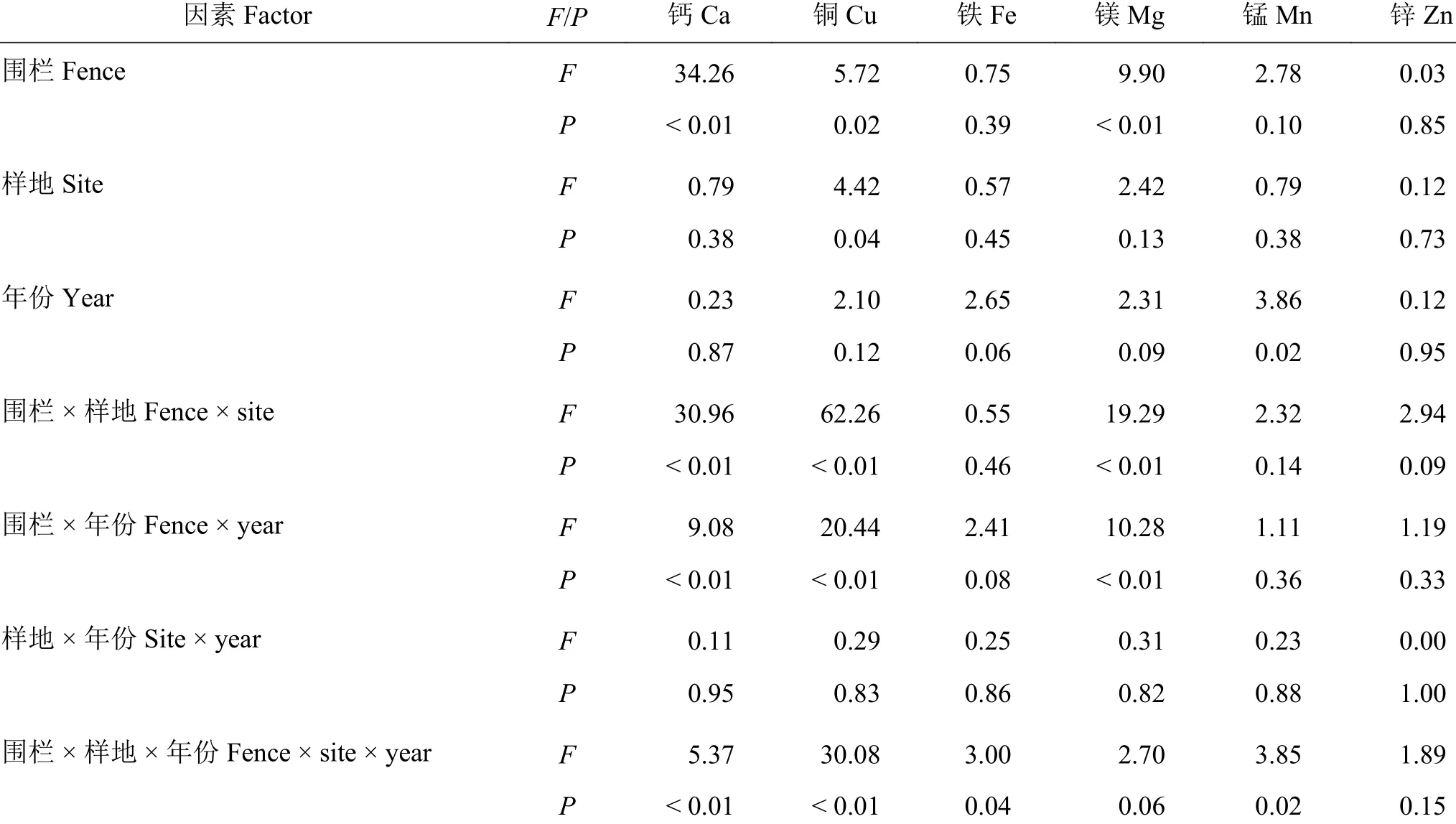
表2 围栏、样地、年份及其交互作用对土壤矿质元素影响的混合效应模型分析Table 2 Effects of fencing, site, year, and their interactions on soil mineral elements from mixed-effect model analysis
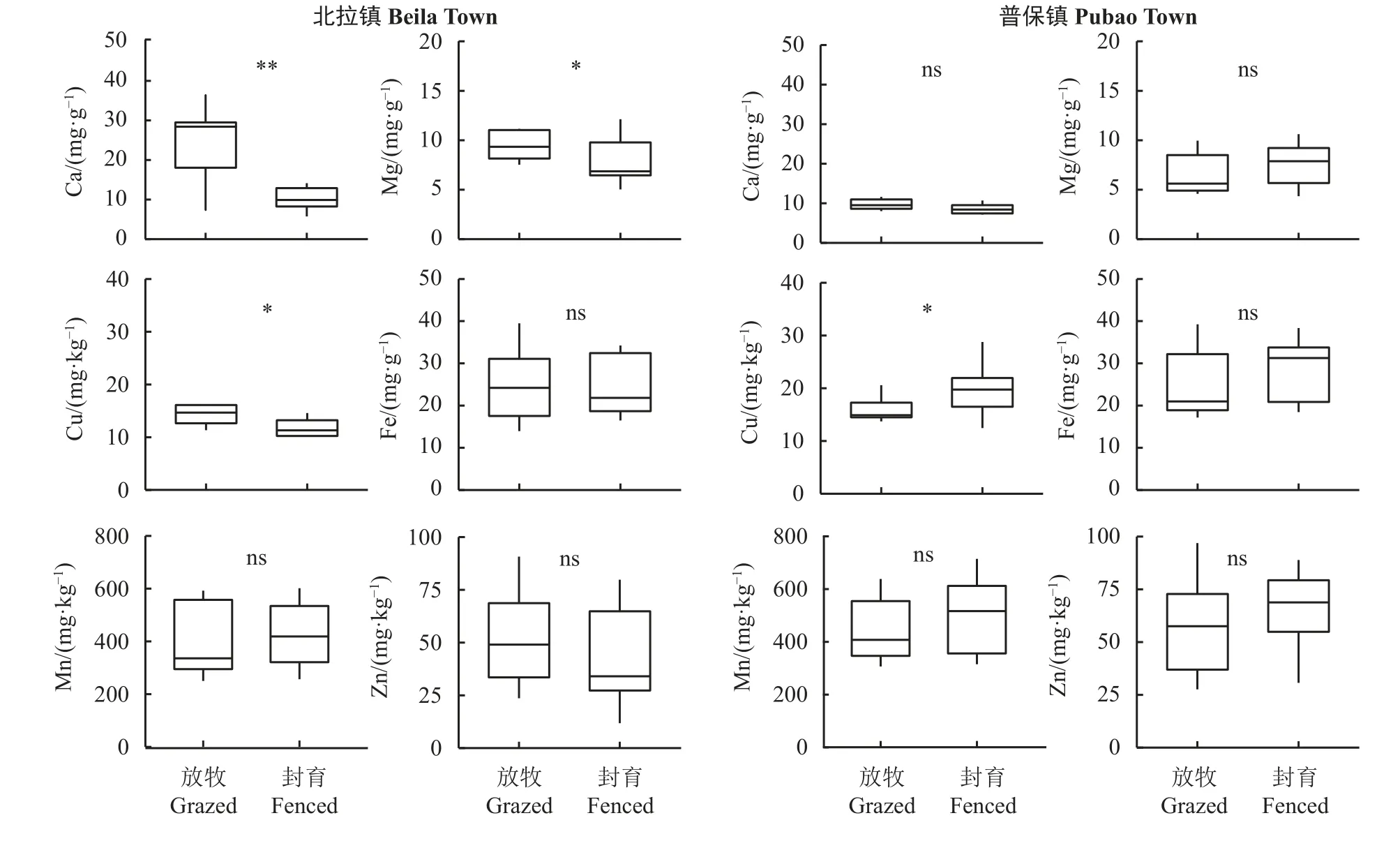
图3 各样地围栏内外矿质元素含量统计Figure 3 Mineral element contents inside and outside the fences of each site
2.2 围栏封育对植物群落特征的影响
围栏内外地上生物量年际变化无明显规律。围栏内外地上生物量及Shannon-Wiener 指数相对大小无一致规律(图4)。2013年普保镇围栏内地上生物量(78.48 g·m-2)比围栏外(41.48 g·m-2)高89.2% (P<0.05),其他封育年份围栏内外地上生物量及Shannon-Wiener 指数均无显著差异(P> 0.05) (图4)。仅2012年北拉镇封育样地的地上生物量与2011年相比差异显著,2011-2012年和2012-2013年普保镇放牧样地的Shannon-Wiener 指数年际变化显著(P< 0.05),其他年份间地上生物量及Shannon-Wiener 指数的变化均不显著(P> 0.05)。
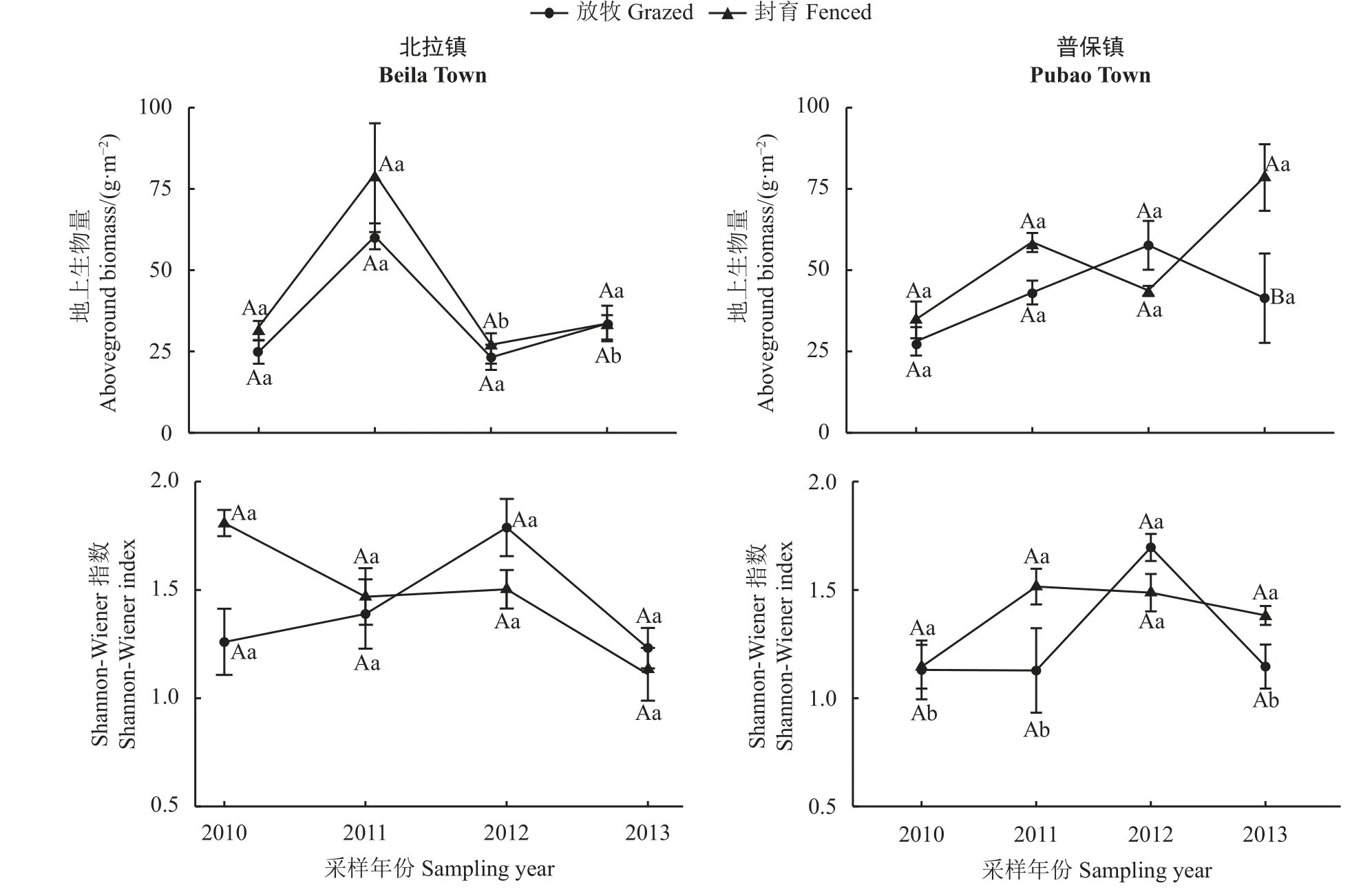
图4 各采样年份地上生物量及Shannon-Wiener 指数Figure 4 Aboveground biomass and Shannon-Wiener index in each sampling year
与放牧地相比,围栏封育对地上生物量的影响显著(P< 0.05) (表3)。其中,围栏封育仅在普保镇对地上生物量有显著影响(增加了43.8%) (图5)。围栏 × 样地、围栏 × 年份及围栏、样地和年份三者的交互作用对地上生物量的影响显著(P< 0.05) (表3)。仅围栏 × 年份的交互作用对物种多样性有显著影响。但与放牧样地相比,围栏封育对两个样地物种多样性没有显著影响(P> 0.05) (图5)。

图5 各样地围栏内外地上生物量及Shannon-Wiener 指数Figure 5 Aboveground biomass and Shannon-Wiener index inside and outside the fences of each site

表3 围栏、样地、年份及其交互作用对群落特征影响的混合效应模型分析Table 3 Effects of fencing, site, year, and their interactions on plant community characteristics from mixed-effect model analysis
2.3 气候特征、群落特征和土壤矿质元素间的关系
相关性分析显示,生长季气温与地上生物量和多样性分别极显著负相关和极显著正相关,生长季降水与地上生物量和多样性分别极显著正相关和极显著负相关(P< 0.01) (图6)。生长季气温与各矿质元素极显著正相关,但生长季降水与各矿质元素显著负相关。与生长季降水相比,生长季气温与矿质元素的相关系数绝对值更大。各矿质元素之间,Ca 与Mg、Cu与Zn、Mn与Zn、Fe与Mg、Mn和Zn之间极显著正相关(P< 0.01)。仅土壤Mg与地上生物量显著负相关(P< 0.05),但围栏内或围栏外地上生物量随土壤矿质元素含量增加变化不显著(P> 0.05) (图7)。土壤矿质元素与物种多样性均无显著关系(P> 0.05)(图6),但围栏外物种多样性随土壤Zn含量增加呈先减小后增加的趋势,且变化显著(P< 0.05) (图8)。
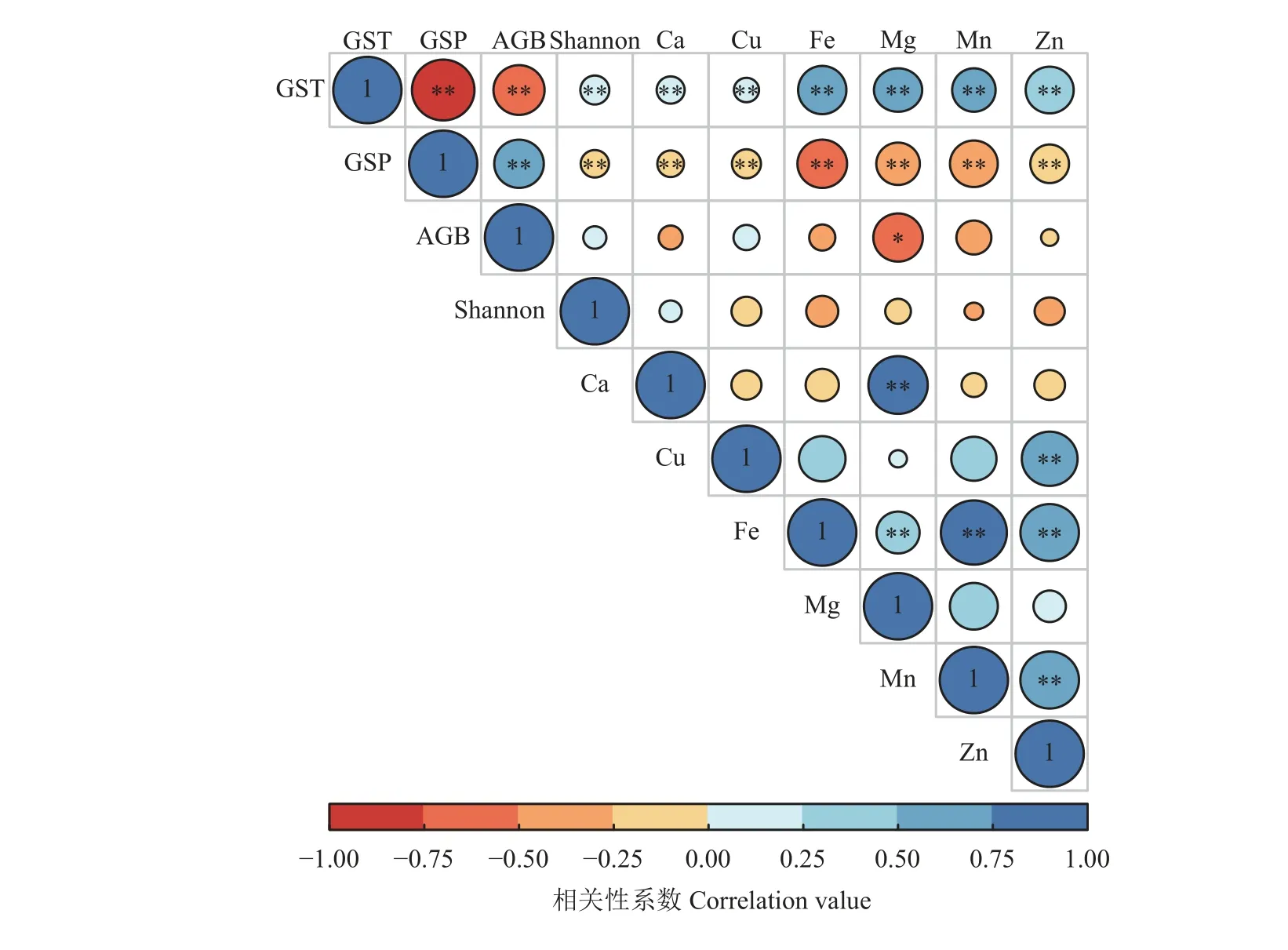
图6 气候因子、群落特征与土壤矿质元素之间的相关系数矩阵图Figure 6 Correlation coefficient matrix of climatic variables, plant community regimes and soil mineral elements

图7 地上生物量与气候变量和土壤矿质元素的关系Figure 7 Relationships of aboveground with climatic variables and soil mineral element contents
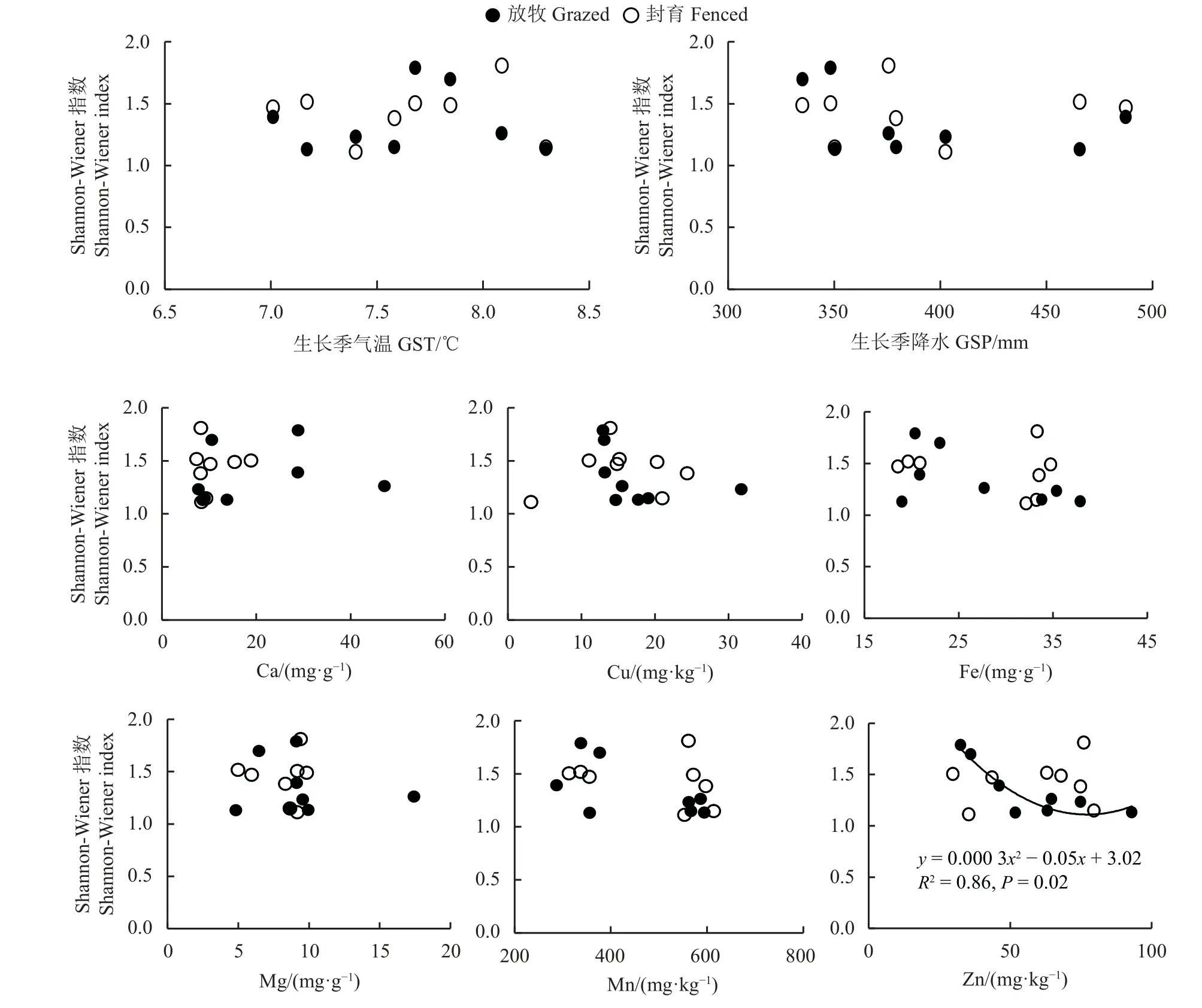
图8 Shannon-Wiener 指数与气候变量和土壤矿质元素的关系Figure 8 Relationships of the Shannon-Wiener index with climatic variables and soil mineral element contents
以各矿质元素为协变量进行分析,结果显示围栏封育对高寒草原地上生物量和物种多样性无显著影响(P> 0.05) (表4)。仅土壤Mg对地上生物量有显著影响(P< 0.05),围栏封育与土壤Zn的交互作用对物种多样性有显著影响,其他土壤矿质元素及其与围栏封育的交互作用对地上生物量和物种多样性均无显著影响(P> 0.05)。

表4 地上生物量和物种多样性的协方差分析Table 4 Results of ANCOVA models for above-ground biomass and biodiversity
3 讨论
3.1 围栏封育对土壤矿质元素含量的影响
矿质元素是土壤养分重要成分,其含量和分布与母质类型、成土过程、土壤理化性质、土地利用方式和环境污染等因素有关[35]。在青藏高原退化草地和内蒙古典型草原的研究发现,围栏封育会降低部分样地表层土壤Ca、Mg和Cu的含量[36-38]。与前人研究相似,本研究在北拉镇的结果表明,高寒草原实施5~8年围栏封育后表层土壤Ca、Mg和Cu的含量显著低于自由放牧样地土壤。这可能是因为家畜排泄物含有Ca元素[39],且Ca与Mg相关性较强(表3),造成围栏外土壤Ca和Mg含量偏高;另外,在本研究中研究区域每公顷草的放牧强度小于1只羊[40],放牧强度不高,没有对土壤养分造成过度破坏,这可能更有利于土壤养分循环和肥力保持[41-42]。
此外,实施围栏封育后普保镇地土壤Cu含量增加了23.5%。这与杨红善等[43]在肃南县山地草原的研究一致。他们的研究发现禁牧使0 - 10 cm土层Cu含量增加了13%。排除牲畜对草地植物的采食与践踏后,围栏封育可能降低土壤容重,增加植被盖度和高度,提高土壤持水能力,促进有机质和凋落物分解[44-45],进而改善土壤矿质元素含量。
本研究发现8年围封没有显著改善高寒草原表层土壤Fe、Mn和Zn的含量。这与在青藏高原东北部的研究[46]相似,高寒草甸土壤微量元素含量也不受围栏封育的影响。藏北地区气候寒冷干燥,且本研究封育时间较短。这可能会限制土壤中矿物质分解速率[47],影响土壤微量元素有效性。另外,班戈县土壤砂粒较多[48],对微量元素吸附能力较差,造成微量元素易被淋溶,从而限制了土壤养分的恢复。
高寒草原表层土壤Mn含量在不同年份间差异显著,其他矿质元素含量在各年间无显著差异(表2)。可能是各样地每年的降水和气温有差异,并且与其他矿质元素相比,Mn与气候因素的相关性更强(图6),意味着土壤中Mn含量更容易受到气候的影响[49]。
3.2 围栏封育对群落特征的影响
气候变化和人类活动引起草地退化,对植物群落的影响往往表现为生物量下降,物种多样性降低等[1]。为了恢复退化草地,人们常采用围栏封育等方法[2]。围封是提高草地地上生物量的有效途径[6-7]。与前人结果研究相似,本研究发现5~8年的围栏封育增加了普保镇地上生物量。这可能是因为排除放牧压力后消除了食草动物对植物的干扰,植被高度和盖度增加,光合效率随之增加,从而生物量得到恢复。
对退化草地实施围封,消除了外界干扰源,对生物量和物种多样性的增加有积极作用[12],但关于围栏封育对地上生物量和群落多样性的影响仍存在争议。有研究发现地上生物量和物种多样性并未随着封育时间增加而持续增加[13,50]。一项在内蒙古典型草原的研究表明[50],较短期封育(2年和7年)并未使群落多样性得到显著恢复。相似地,在本研究中,围栏封育未显著增加北拉镇地上生物量和班戈县植被多样性。不同研究地点放牧历史和放牧率不同,这可能导致物种多样性对围栏封育的响应不同[51]。藏北高原海拔高、气温低,可能使植被收到水分或氮的限制,从而限制了植被生长和物种多样性的恢复[52]。
3.3 矿质元素对群落特征的影响
矿质元素的组成与含量对植物个体生长发育、群落动态演替与生态系统服务功能发挥具有关键调控作用[53]。在对甘肃5种类型草地的研究[54]中发现,土壤微量元素对植被特征(盖度、高度和生物量)有重要影响,尤其是Mg和Zn。本研究所测土壤Mg含量约为8.25 mg·g-1,高于其他在青藏高原进行的研究[55]。围栏封育显著改变了土壤Mg含量,土壤Mg对地上生物量有显著负影响。
本研究所测土壤Zn含量为29.78~79.68 mg·kg-1,低于我国土壤Zn含量平均值[56](100 mg·kg-1);围栏封育未显著改变高寒草原土壤Zn含量,土壤Zn含量与对上生物量无显著影响,但围栏封育与土壤Zn的交互作用对物种多样性有显著影响。研究区域土壤砂粒土含量多且偏碱性,容易缺乏微量元素[57]。土壤矿质微量元素对草地生产力存在潜在的调控作用[26],围栏封育可能通过影响土壤矿质元素对生物量物种多样性等群落特征产生间接影响。因此,在草地恢复过程中,除关注土壤养分和常见理化性质外,还应关注研究较少的矿质元素(特别是Mg和Zn)在土壤和地上植被中的含量,从而更全面地探讨围栏封育等恢复措施对草地生态的恢复机制。
4 结论
本研究分析了围栏封育对藏北高寒草原植物群落特征和土壤矿质元素含量的影响。5~8年的围栏封育只显著改变了表层土壤Ca、Mg和Cu的含量,没有显著改变土壤中Fe、Mn和Zn的含量。围栏封育只显著增加了普保镇地上生物量,对两个样地植物物种多样性均无显著影响。在各土壤矿质元素中,仅Mg与地上生物量存在显著相关性。这可能是因为围栏封育对高寒草原土壤矿质元素含量和植物群落的恢复效果有限。建议设置长期固定样地探究围栏封育下土壤矿质元素变化对草地生态系统功能的调控机制。
猜你喜欢
绿色天府(2022年9期)2022-11-19
中国交通信息化(2022年7期)2022-10-27
农业工程学报(2022年10期)2022-08-22
世界热带农业信息(2022年8期)2022-07-19
贵州畜牧兽医(2022年3期)2022-06-28
林业调查规划(2022年3期)2022-06-13
红蜻蜓(2020年12期)2020-06-20
安徽农学通报(2020年7期)2020-05-26
好孩子画报(2019年8期)2019-09-19
发明与创新·大科技(2018年2期)2018-03-17
