
中国板栗36个叶片表型性状的多样性
2022-04-21 08:54郭燕张树航李颖张馨方王广鹏
中国农业科学 2022年5期
郭燕,张树航,李颖,张馨方,王广鹏
中国板栗36个叶片表型性状的多样性
郭燕,张树航,李颖,张馨方,王广鹏*
河北省农林科学院昌黎果树研究所,河北昌黎 066600
【目的】揭示中国板栗(Bl.)叶片表型性状的变异规律及多样性特征,为中国板栗资源的保护、开发和科学利用提供依据。【方法】对中国10个省份(群体)240份资源的36个叶片表型性状(31个数量性状和5个质量性状)进行检测,运用方差分析、多重比较、相关分析和聚类分析等方法,探讨板栗叶片表型多样性水平及其与原产地的地理、环境因子关系。【结果】巢式方差分析表明,31个数量性状在群体内存在极显著差异(<0.01),同时叶片长、叶片宽、叶片长/宽、锯齿高度等15个数量性状及3个质量性状(叶片形状、叶基形状和叶缘锯齿特征)在群体间存在显著或极显著差异(<0.05或<0.01),说明板栗叶片表型在群体内和群体间均存在丰富变异。群体间叶片表型分化系数VST为7.95%,远小于群体内的92.05%,说明叶片表型变异主要来源于群体内部。板栗叶片表型的平均变异系数(CV)和平均多样性指数(H′)分别为18.05%和1.27,5个质量性状中以叶片形状的CV和H′最高(35.67%,0.86),叶缘特征最低(5.60%,0.07);31个数量性状的CV在8.68%(叶片长/宽)—32.73%(锯齿深度),H′在1.54(叶背茸毛密度)—2.09(叶片结构疏松度)。叶片外观形态、生理指标和解剖性状的平均H′分别为2.03、2.04和2.04,CV由高到低分别为15.27%(外观形态)>15.18%(生理指标)>12.17%(解剖结构)。10个群体表型性状CV均值在17.11%(陕西)—19.66%(山东),H′均值在0.99(安徽)—1.80(河北)。板栗叶片大多表型性状与地理气候因子之间存在显著或极显著相关,其中年均气温等温度指标和年降水量与叶基形状、锯齿高度等性状呈负相关(经度和纬度与之呈正相关),与叶片长/宽呈正相关(经度和纬度与之呈负相关)。【结论】板栗叶片表型在群体间和群体内均存在丰富变异,但变异主要来源于群体内(个体基因型间的变异),因而以叶片表型性状作为研究对象,进行抗逆性、高光合效率等特异品种选育或相关研究时,研究重点应该放到广泛的个体基因型调查上,变异较为丰富均匀的群体可作为重点参考。板栗叶片大多外观形态和生理指标性状呈现以温度和降雨量因子为主导的梯度规律性。
板栗;叶片表型性状;群体间;群体内;变异规律;表型多样性
0 引言
【研究意义】遗传多样性的研究是种质资源创新利用的基础。植物表型性状是其各种形态特征的总合,是遗传因子和环境因子综合作用的结果[1-2]。长期的自然选择,植物的表型性状会不可逆地形成新表型性状并能稳定地遗传给后代,即表型性状变异必然蕴藏着遗传变异[3-4]。因此,通过表型性状来检测植物遗传变异规律,对挖掘各种基因型资源并加以利用具有重要意义。【前人研究进展】叶片是植物进行光合、呼吸和蒸腾作用的重要器官,其形态、解剖特征会直接影响生理生化、生长发育及繁殖特性,最终影响植物的适应、生存和进化[5-7]。如叶柄越长,越有利于减少叶片对光能的相互遮挡,光能利用率越高[8-9];叶片长宽比大,有利于植物获得更多的光能[10];叶片锯齿深度越大,液压效率越高,植物对干旱环境的适应能力越强[11];较厚的叶片、表皮细胞和栅栏组织可以贮藏更多的水分,从而提高植物叶片的储水性能而耐旱[12]。叶片性状已经被广泛用于水青树()、山核桃()、梨()和欧洲李()等植物遗传多样性的研究,并明确了相应的叶片表型变异规律[6,13-15]。板栗(Bl.)是中国重要的经济林树种,分布范围极广,种质资源也极其丰富[16],但目前关于板栗叶片遗传多样性的研究所涉及的群体、个体及性状数量非常有限。周连第等[17]仅对8个产地69个板栗品种的3个性状(叶片长、叶片宽和叶片长宽比)进行了分析;江锡兵等[18]调查了10个省份90个板栗地方品种4个性状(叶片长、叶片宽、叶形指数和叶柄长);刘亚斌和郭素娟[19]仅对定植在河北迁西的7个省份的39份板栗资源的8个性状(叶片长、叶片宽、叶片长宽比、叶片面积、叶片周长、叶片干质量、比叶质量和叶绿素含量)进行了分析,总体认为,叶片表型在群体内和群体间均存在广泛变异,群体内变异是板栗叶片表型变异的主要来源,南方板栗的叶片长度和叶形指数整体上比北方板栗品种大。【本研究切入点】有关板栗上叶柄、锯齿、叶脉等形态表型以及与生长发育有着密切关系的解剖结构和生理指标性状的多样性研究鲜见报道,同时,叶片性状与地理因子的关联性也涉及不多。因此,有关中国板栗叶片表型遗传多样性的信息有待进一步完善。【拟解决的关键问题】本研究以来源于我国10个板栗主产区省份(群体)的240份实生良种、地方品种和古树资源为材料,对其叶片形态表型、解剖结构和生理指标的36个性状进行表型多样性、相关性及群体聚类分析,进一步揭示中国板栗叶片表型多样性特征和变异规律,为中国板栗资源的保护、开发和科学利用提供参考依据,为叶片表型相关性状的基因定位奠定前期基础。
1 材料与方法
试验于2019—2020年在河北省农林科学院昌黎果树研究所板栗种质资源圃进行。
1.1 试验地概况
试验地属中国东部季风区暖温带半湿润大陆性气候,年均气温12.2℃,1月平均气温-6.3℃,7月平均气温26.1℃,年均降水量643 mm,年均日照时数为2 663.7 h,海拔20 m,沙壤土。
1.2 试验材料
供试240份种质资源分别来源于河北、山东、北京、湖北、湖南、江苏、浙江、安徽、陕西和广西10个群体(表1),于2004年统一嫁接于3年生‘燕山早丰’实生砧木上,每份资源嫁接5株,株行距4 m×4 m,植株常年管理措施一致。分别于2019年和2020年的7月15—20日,选取每份资源生长势基本一致的植株3株,在每株东、南、西、北向冠层外围中部随机选取1年生健康营养枝3条,在其上选取第7—9节位叶片3片,即每份资源每株树取叶片36片作为一个样品,3株树(3个样品)共取108片叶片。样品以密封袋密封后置于冰盒带回实验室,每份资源的3个样品先分别随机选取10片叶用锡箔纸平展包裹,液氮速冻后放入-80℃冰箱用于解剖结构(5片)和叶绿素含量(5片)观测,其余样品叶片用于叶片保水力、比叶质量、外观形态、叶片颜色及气孔特征测定。
每份种质资源的原产地地理位置和气候因子由当地政府门户网站或气象局提供,经整理后各群体概况见表2。
1.3 测定指标及方法
外观形态:每份资源的3个样品中各随机选取9片叶,分别用于叶片外观形态的20个表型性状(5个质量性状和15个数量性状)测定,取平均值作为实测值(数量性状)。叶柄长、叶柄宽、叶柄长/宽、叶片长、叶片宽、叶片长/宽、叶片面积、锯齿数、锯齿高度、锯齿深度和叶脉间距采用万深LA-S系列植物图像分析软件分析叶片扫描图片测量;锯齿与叶缘间角度采用SPOT图像分析软件分析叶片扫描图片测量;叶背茸毛密度参照刘庆忠[20]方法测定;叶片正面颜色参数(L*、a值)使用精密色差仪(ColorLite-sph 860)测定。叶片形状、先端形状、叶基形状、叶缘特征和叶缘锯齿方向5个质量性状,参照LY/T1851—2009板栗测试指南[21]中的方法观测,选择9次测定中频率最高形态作为该资源的质量性状描述形式。
解剖结构:将冰箱中备用的每份资源的3个样品(各5片)叶片取出,解冻后分别单独制片,每片叶选择完整平展的1个玻片观测,取平均值作为实测值。叶厚、上表皮细胞厚度、上表皮细胞宽度、栅栏组织厚度、海绵组织厚度、下表皮细胞厚度和下表皮细胞宽度采用冷冻切片法制片和测量[22],栅海比、叶片结构紧密度和叶片结构疏松度按照徐扬等[12]的方法计算;气孔长、气孔宽和气孔密度采用指甲油印迹法制片和测量[23]。
生理指标:每份资源3个样品中分别随机选取17片叶,用于叶片生理指标性状的测定。其中比叶质量测定参照刘亚斌和郭素娟[19]的方法;叶片保水力测定采用离体叶片自然失水法[24]。叶绿素含量测定将冰箱中备用的3个样品(各5片)取出,解冻后分别采用80%丙酮直接浸提法[25]测定。
1.4 数据处理
对叶片的36个表型性状采用巢氏设计方差分析,其线性模型为:Y=μ+S+T(i)j(ij)k。式中Y为第个群体第个无性系第个个体的观测值,为总体平均值,S为第个群体的效应值,(i)j为第个群体内第个无性系的效应值;(ij)k为试验误差。表型分化系数(VST)、变异系数(CV)、Shannon’多样性指数(H′)的计算参照李洪果等[26]的方法,其中VST(%)=[2t/S/(2t/S+2S)]×100。式中:2t/S和2S分别为群体间和群体内方差值;CV(%)= s/X× 100,式中:X为性状平均值,s为标准差;根据平均值(X)和标准差(s)将测量性状分为10等级,1级<X-2s,10级≥X+2s,中间每级相差0.5 s,通过计算每一级的相对频率(P)来计算Shannon’多样性指数(H'),H'=-ΣlnP,式中P表示第级别内资源份数占总份数的百分比。
采用Excel 2010和SPSS 20统计软件对各性状进行方差分析、Duncan’s多重比较、相关性分析及聚类分析。椭圆形,占63.33%;椭圆形次之,占31.25%;铍针状、卵状和倒卵状极少,3种形状的种质仅占5.42%。2种先端形状中,240份材料里急尖仅12份,渐尖较为普遍(228份)。在叶基形状3种类型中,楔形和钝形较为普遍,楔形略少于钝形,两者占97.5%;心形极少,仅占2.5%。叶缘特征多为具锯齿,所调查种质中仅有3份为具短尖。叶缘锯齿方向多为外向,占83.33%;直向次之;内向最少,仅占3.75%。

表1 240份板栗种质资源编号、来源、名称及资源类型

续表1 Continued table 1
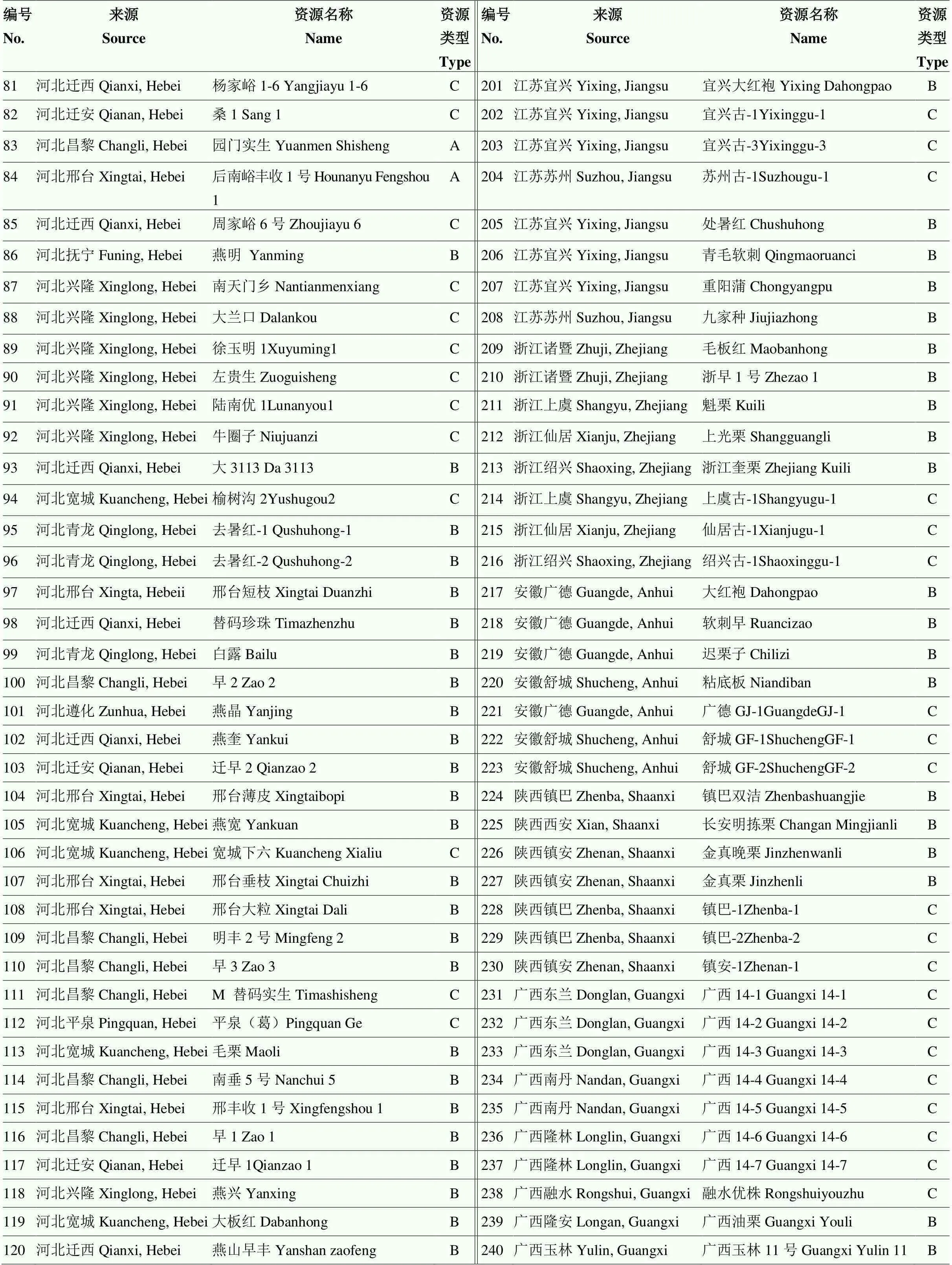
续表1 Continued table 1
A:实生良种;B:地方品种;C:古树资源 A: Excellent seed variety; B: Local variety; C: Ancient trees resource
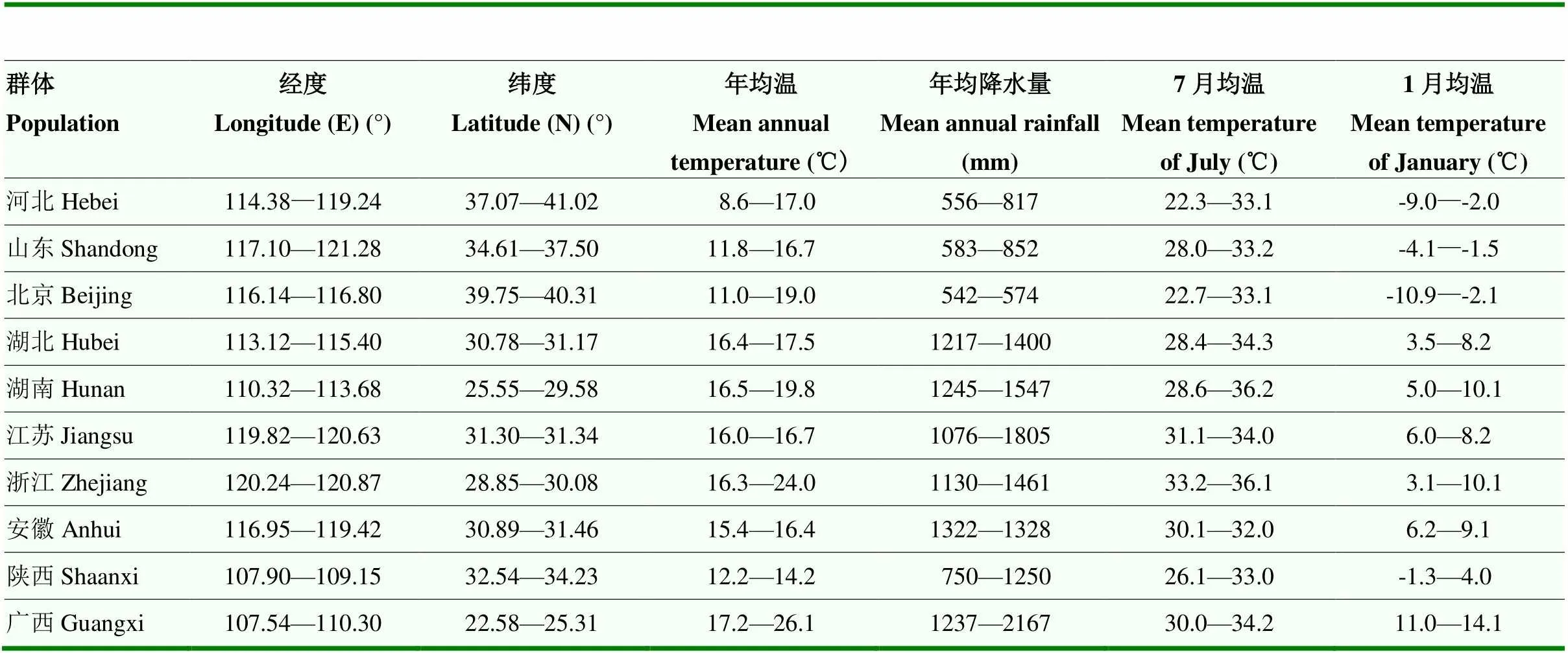
表2 板栗10个群体的地理位置及主要气候因子范围
2 结果
2.1 板栗种质资源叶片表型性状多样性分析
240份板栗种质资源5个叶片质量性状频率分布见表3,叶片典型形态特征对应图形见图1。从表3可知,叶片形状和叶基形状变异系数均在30%以上,多样性指数均在0.7以上,而先端形状和叶缘特征的变异系数和多样性指数均较小,变异系数分别为11.2%和5.60%,多样性指数分别为0.20和0.07。进一步分析发现,5种叶片形状中,收集的240份资源多为长

A:叶片形状:1:椭圆形(双台大红袍);2:长椭圆形(燕山早丰);3:披针形(金杖子1号);4:卵形(山东红栗);5:倒卵形(乳山短枝)。B:先端形状:1:急尖(乳山短枝);2:渐尖(大板红)。C叶基形状:1:楔形(玫瑰红);2:钝形(大板红);3:心形(塔54)。D:叶缘特征:1:具短尖(融水优株);2:具锯齿(大板红)。E:叶缘锯齿方向:3:外向(大板红);5:直向(黄前);7:内向(晚早5)
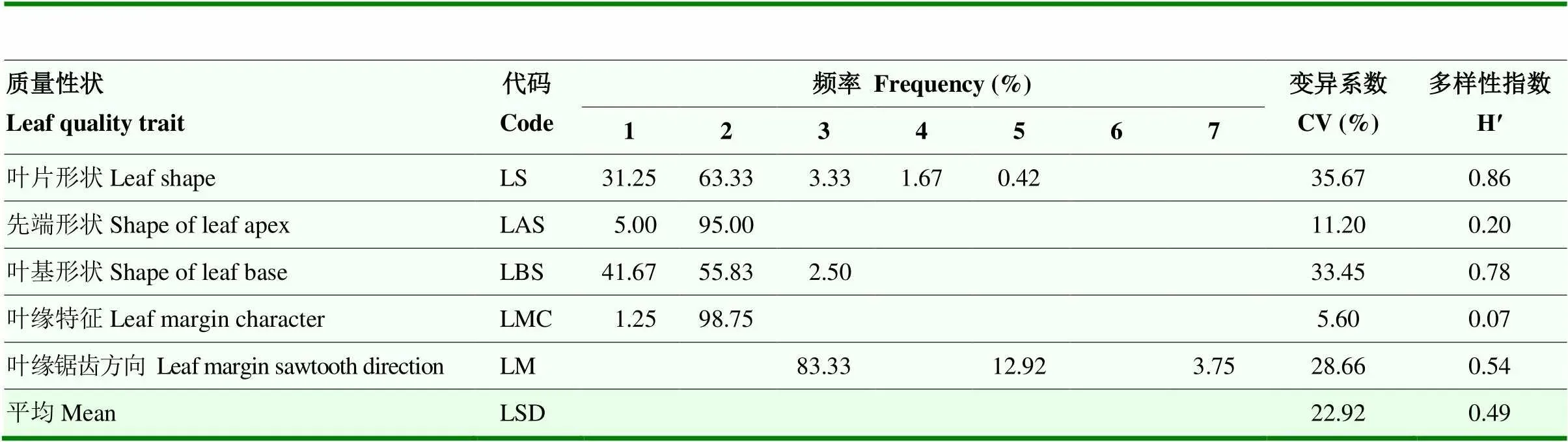
表3 板栗种质资源叶片质量性状频率分布及多样性
由表4可知,31个数量性状变异系数介于8.68%(叶片长/宽)—32.73%(锯齿深度),15个叶片外观形态性状中,锯齿深度的变异系数最大(32.73%),变异幅度为1.03—6.63 mm,叶片长/宽的变异系数最小(8.68%),变异幅度为2.03—3.35;另外,叶片长/宽的变异系数小于叶片长和叶片宽,说明叶片长/宽较单个叶片长或叶片宽性状稳定。13个叶片解剖结构性状中,栅栏组织/海绵组织的变异系数最大(15.60%),变异幅度为0.69—2.47,叶片结构疏松度变异系数最小(9.81%),变异幅度为24.51—41.30。3个生理性状的变异系数均值为15.18%。结合表2,由板栗叶片表型性状的平均变异系数来看,叶片表型性状的变异离散程度由高到低分别为:叶片质量性状(22.44%)>外观形态数量性状(15.27%)>生理指标数量性状(15.18%)>解剖结构数量性状(12.17%)。
31个数量性状的遗传多样性指数(H′)介于1.54— 2.09,均值2.04。结合表3和表4分析,叶片外观形态H′均值为2.03,解剖结构和生理数量性状的H′均值均为2.04,远高于叶片质量性状(0.48),说明板栗叶片3种类型数量性状的遗传变异均匀度较好,而质量性状的遗传变异虽然离散程度大,但是变异较为集中,均匀度较差。
植物不同表型性状的极值可以反映该性状在不同环境条件下的进化及适应性潜力。31个数量性状的极大值为极小值的1.65(气孔长)—6.44(锯齿深度)倍。15个外观形态性状中,锯齿深度(SD)的极大值与极小值的比值最大(6.44),叶片长/宽(LLW)的极大值与极小值的比值最小(1.65);叶片解剖的13个性状中,下表皮细胞厚度(LET)的极大值与极小值的比值最大(3.80),气孔长(SL)的极大值与极小值的比值最小(1.65)。3个生理性状的极大值与极小值的比值均高于2.27。31个数量性状的极值分析表明,叶片大小、锯齿深度、各组织结构厚度、气孔密度和生理指标等性状具有较高的进化或适应性潜力,叶片长/宽、气孔大小等性状的进化或适应性潜力较小。
2.2 板栗叶片表型性状在群体间的变异和分化
5个质量性状的群体间值介于1.10—9.27,其中叶片形状(LS)、叶基形状(LBS)和叶缘锯齿方向(LSD)在不同群体间存在极显著差异,而先端形状(LAS)和叶缘特征(LM)在群体间差异不显著(表5)。
板栗叶片31个数量性状在群体间和群体内的方差分析结果和表型分化系数见表6。由表6可知,31个数量性状在群体内均存在极显著差异,值介于1.43— 16.44,同时15个数量性状在群体间存在显著或极显著差异(值介于1.88—8.78),说明板栗叶片表型性状在群体内和群体间均存在一定程度变异。31个数量性状在群体间的表型分化系数(V)值介于3.01%— 25.57%,平均为7.95%,群体内为92.05%,群体内变异远高于群体间变异,说明群体内变异是板栗叶片表型变异的主要来源。另外,进一步比较发现,3类数量性状的VST由高到低分别为叶片外观形态(9.74%)>生理指标(8.22%)>解剖结构(5.82%),说明板栗叶片外观形态和生理指标性状的群体间变异相对较大,其易受地理环境因素的影响,解剖结构性状的群体间变异相对较小,不易受到地理环境因素的影响。
由表7可知,河北、北京和山东板栗群体普遍表现出叶片呈椭圆形、叶基钝形,叶缘锯齿外向、叶片长宽比值较小,锯齿深度、锯齿高度和锯齿与叶缘间角度较大的特征;湖北、湖南、江苏、浙江、安徽和广西群体普遍表现出叶片呈长椭圆形、叶缘锯齿直向、叶基楔形、叶片长宽比值大,锯齿深度、锯齿高度和锯齿与叶缘间角度较小的特征;安徽、陕西和江苏群体的叶背茸毛密度和|a|显著高于其他群体;河北和山东群体均表现出上表皮细胞厚度和叶片保水力较大,下表皮细胞厚度较小的特征;湖南、浙江和安徽的叶柄长度显著大于其他群体;广西和浙江群体的叶绿素含量显著低于其他群体。
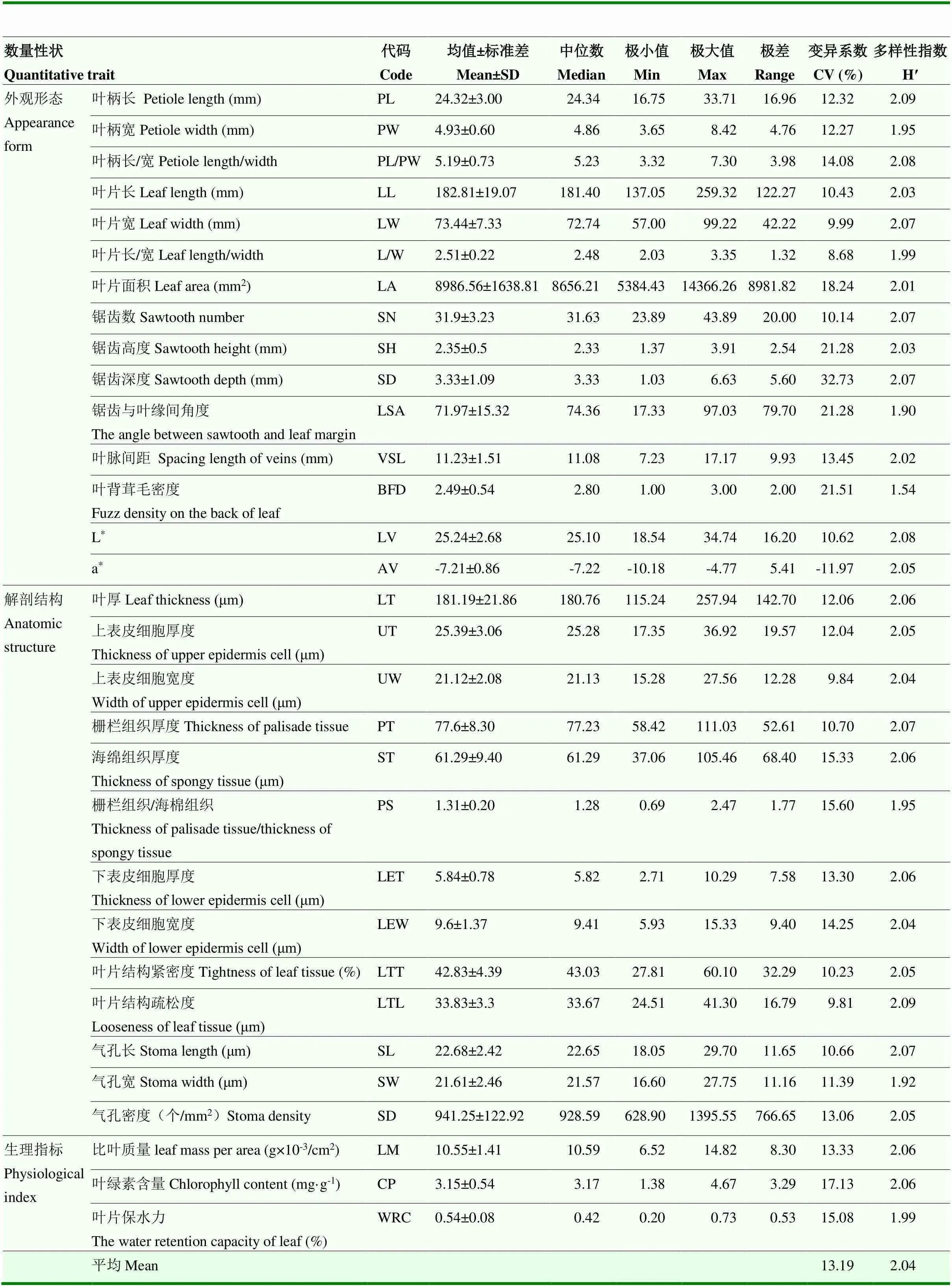
表4 板栗叶片数量性状变异

表5 板栗叶片质量性状群体间方差分析结果
*:<0.05,**:<0.01。下同The same as below
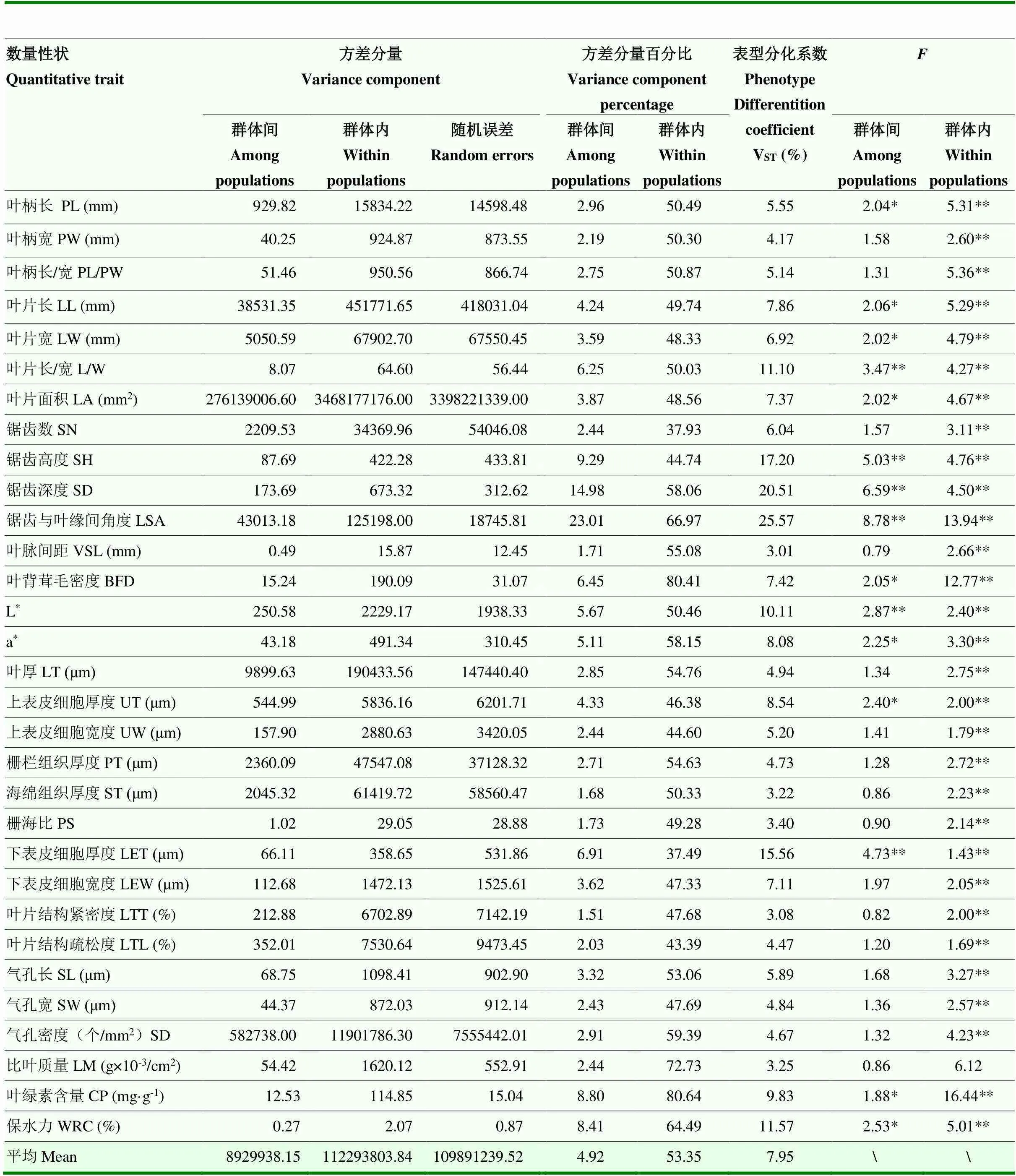
表6 板栗叶片数量性状的方差分析结果和表型分化系数
表7 10个板栗群体18个叶片表型性状的多重比较(均值±标准误)
Table 7 Multiple comparison of 18 leaf phenotypic trait of 10 chestnut populations (Mean±SD)
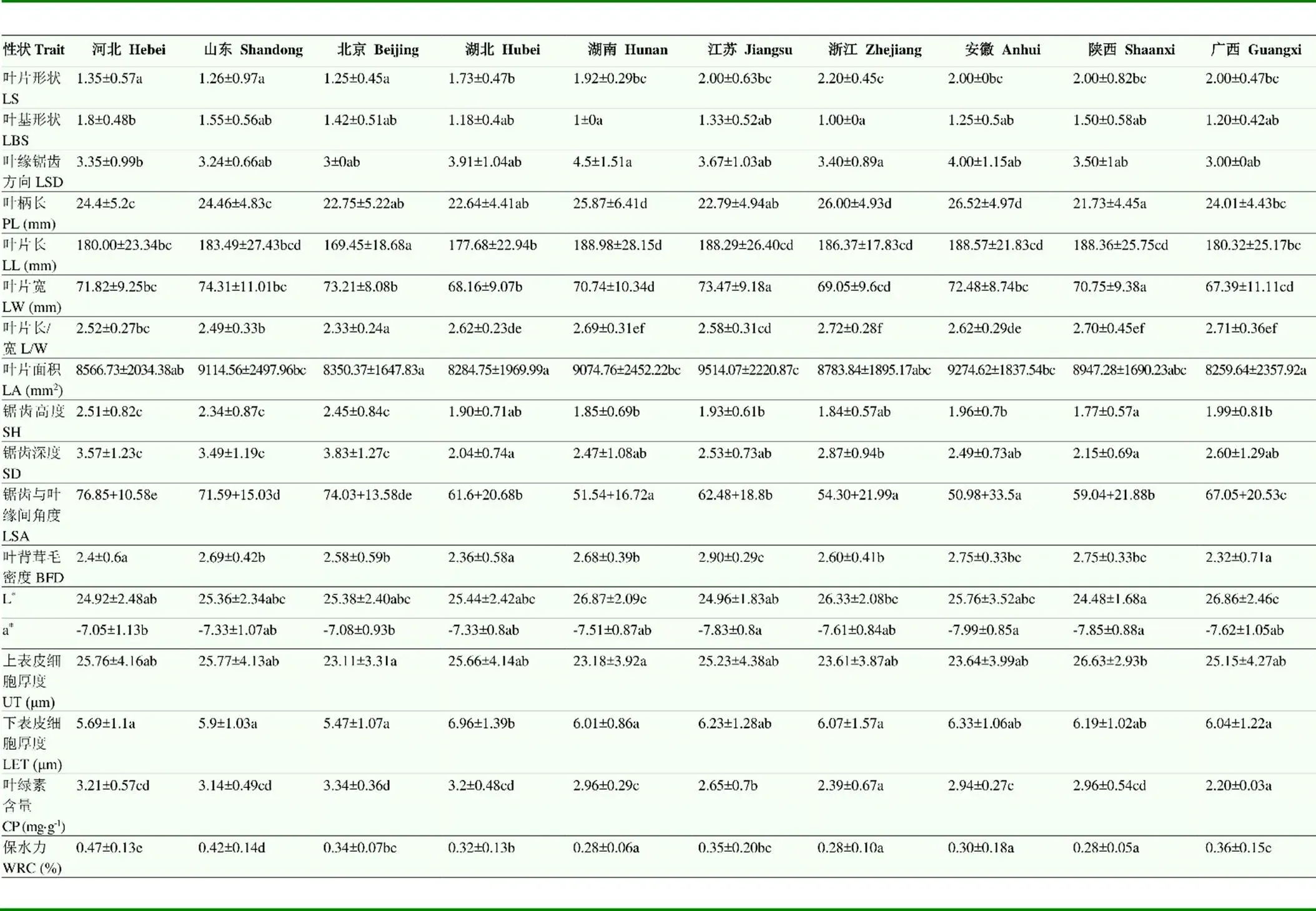
两两群体间不同字母表示差异显著(<0.05) Paired populations without the same letter are significantly different (<0.05)
2.3 板栗群体内叶片表型变异特征和表型多样性
从10个群体种质资源的表型性状变异系数均值来看,其群体内表型变异离散程度由高到低分别为:山东(19.66%)>广西(19.17%)>河北(18.67%)>北京(18.30%)>江苏(18.26%)>湖北(18.20%)>安徽(17.97%)>湖南(17.68%)>浙江(17.63%)>陕西(17.11%)。依据10个群体的均值,其群体内变异丰富及均匀程度由高到低分别为:河北(1.80)>山东(1.71)>北京(1.51)>湖北(1.48)>湖南(1.47)>广西(1.42)>江苏(1.30)>浙江(1.13)>陕西(1.04)>安徽(0.99)(表8)。
2.4 板栗叶片表型性状与原产地地理气候因子相关性
由表9可知,原产地经度和纬度与叶基形状、叶片宽、锯齿高度、锯齿深度、锯齿与叶缘间角度、a*值、叶绿素含量及保水力间均呈显著或极显著正相关,与叶片长/宽、L*值及下表皮细胞厚度间呈极显著负相关。年均气温、年降水量、7月平均气温和1月平均气温与叶基形状、锯齿高度、锯齿深度、锯齿与叶缘间角度、叶绿素含量及保水力间呈极显著或显著负相关,与叶片长/宽、L值、下表皮细胞厚度、下表皮细胞宽度呈极显著或显著正相关。7月平均温度与栅栏厚度、叶片结构紧密度和气孔密度呈极显著负相关。
2.5 基于板栗叶片表型性状的群体聚类分析
使用与年均温和年降水量均具有显著或极显著相关性的11个叶片表型性状对10个群体240份板栗资源采用组间连接法基于欧氏距离进行聚类(图2)。由图可知,在欧氏距离10时,10个板栗群体根据叶片表型性状可以分为3类:第1类为河北、山东和北京群体,均位于北方属于北方板栗品种群,其年均温、年降水量、1月和7月均温较低(表1),叶片主要特征为叶片长/宽较小,锯齿深度、锯齿高度、叶绿素含量、叶片保水力较大。第2类为浙江和广西群体,属于南方板栗群,其年均温、年降水量、1月和7月均温相对较高,主要特征为叶片长/宽较大,叶绿素含量和叶片保水力较小。第3类为江苏、安徽、陕西、湖北和湖南,除了陕西群体,其他4个群体均属于中南部板栗群,主要特征为叶片长/宽、叶绿素含量和叶片保水力居中。
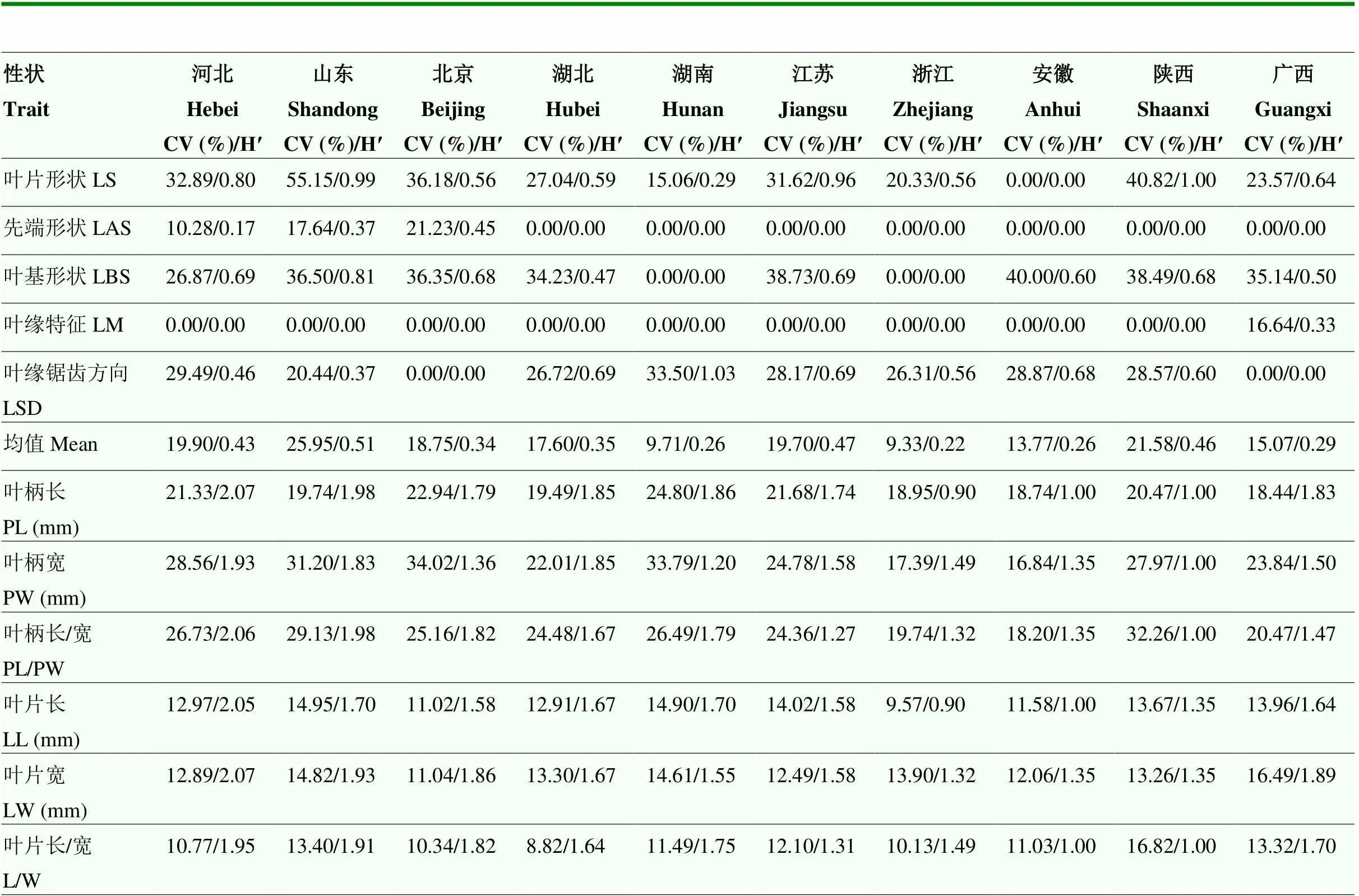
表8 板栗10个群体的叶片表型性状变异系数和遗传多样性指数
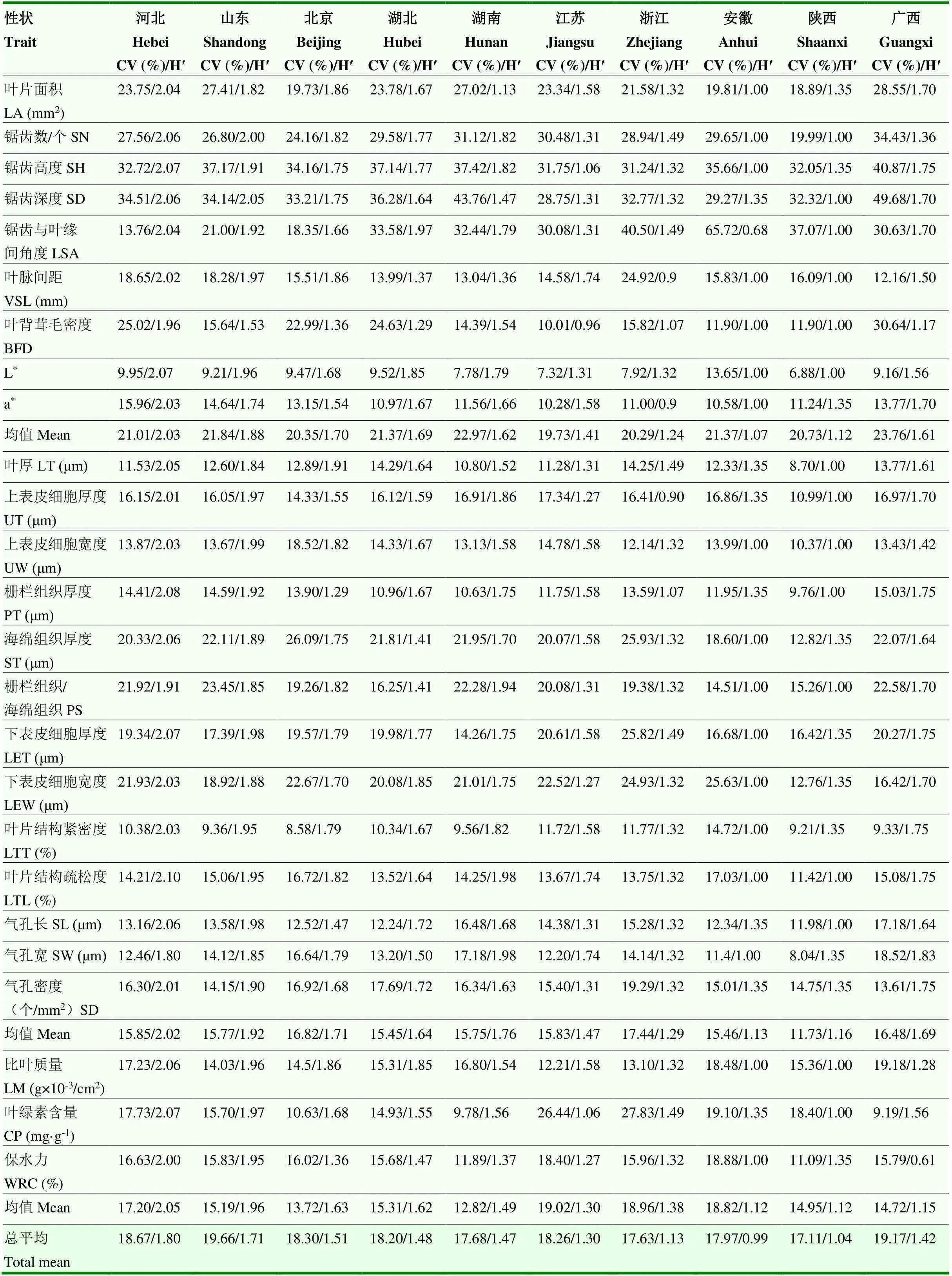
续表8 Continued table 8
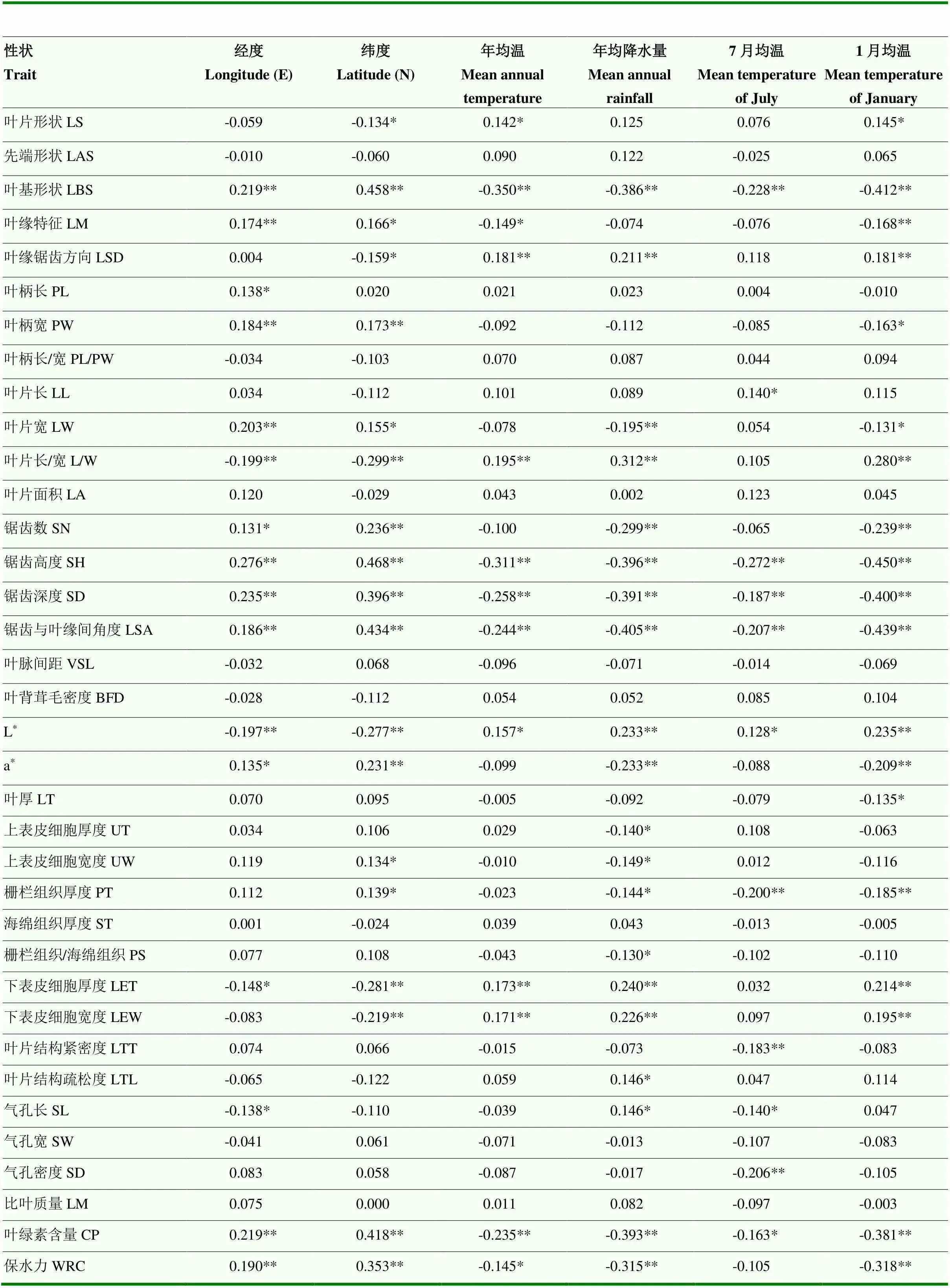
表9 板栗叶片表型性状与地理生态因子间的相关性
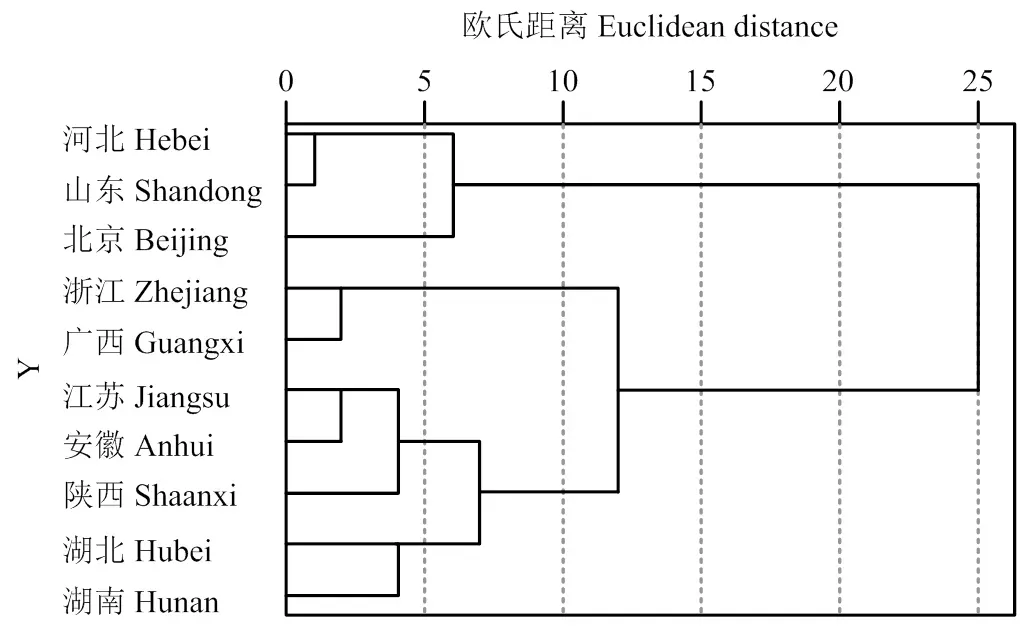
图2 基于叶片表型的板栗群体树状聚类图
3 讨论
3.1 板栗叶片表型性状变异规律
叶片表型性状能有效反映植物的光合和抗逆等特性,其变异与整个植物和生态系统的结构、功能相关[16,27]。本研究通过对源自中国10个板栗群体的240份资源的36个叶片表型质量和数量性状进行方差分析发现,31个数量性状在群体内均存在极显著差异,其中15个性状(大多是外观形态和生理性状)在群体间和群体内均存在显著或极显著差异,这与周连弟[17]、江锡兵[18]和刘亚斌[19]等对板栗叶片长、叶片宽和叶片长/宽性状多样性的研究结果基本相同,说明板栗叶片表型性状在群体内和群体间均存在一定程度变异。
群体内的变异反映的是种质资源个体基因型间的表型变异。种质资源中丰富的高产、优质、抗逆以及高光合效率基因型是品种改良和新品种选育的基础,因此,研究种质资源个体基因型间(不同品种/系间)的性状变异是提高育种效率的根本[28]。本研究结果表明,240份板栗资源叶片表型的平均变异系数和平均多样性指数分别为18.05%和1.27,说明板栗叶片表型在个体水平上存在较为丰富的变异。另外,本研究中31个数量性状的群体间平均表型分化系数(V)为7.95%,群体内为92.05%,说明群体内变异远高于群体间变异,板栗叶片表型在群体间的遗传分化程度较低,这与聂兴华等[29]利用荧光SSR分子标记评估中国栗属植物遗传多样性的结果及田华等[30]和张馨方等[31]基于SSR标记分别对板栗种质资源和自然居群的遗传多样性分析结果一致,也与周连弟[17]、江锡兵[18]和刘亚斌[19]等对板栗叶片长、叶片宽和叶片长/宽表型性状多样性的研究结果相同,这可能是由于引种基因交流频繁等原因造成群体间遗传分化程度较低所致。因此,在以叶片表型性状作为评价指标或筛选目标进行板栗抗逆性相关研究或品种选育时,研究重点应放到广泛的基因型调查上,群体资源的表现只具有参考意义。
本研究结果表明,板栗叶片不同表型性状的个体基因型间变异程度不同预示着板栗不同叶片表型性状在抗逆、分类等相关研究或育种工作中的重要性程度不同,需要先根据不同研究目的筛选出适宜的性状再进入下一步工作,才能确保高效率、准确地获取试验结果。叶片长/宽的变异系数和多样性指数均小于叶片长和叶片宽,这与谢一青等[32]在光皮桦上的研究结果一致,说明叶片长/宽较单个叶片长或叶片宽性状稳定,在植物分类或遗传育种中应该重点考察叶片长/宽,而不是单独考察叶片长或叶片宽。5个质量性状的平均变异系数(22.44%)高于3类数量性状(13.19),而平均多样性指数(0.48)远远低于3类数量性状(2.04),说明质量性状的遗传变异虽然离散程度大,但是变异较为集中,均匀度不好。3类数量性状的多样性指数较为接近,但是平均变异系数由高到低分别为外观形态>生理>解剖结构,而且3类数量性状的表型分化系数排序与变异系数相同,说明无论是在个体基因型水平上还是群体水平上,叶片外观形态和生理性状变异均较为丰富,而解剖结构性状相对较为稳定。
群体间的变异反映了存在地理和生殖隔离的不同群体间的表型变异,是不同环境长期选择的结果,是群体分化的源泉[33-34]。尽管板栗叶片表型群体内变异远远大于群体间变异,但是群体间变异具有独特意义,因为群体间变异真正反映了群体遗传与环境交互效应的复杂关系,其大小在某种程度上说明了该生物对地理环境异质性的敏感程度,值越大受环境异质性影响越大。本研究发现叶片3类数量性状的平均ST由高到低分别为外观形态>生理>解剖结构,说明解剖结构性状最不易发生群体分化,而叶片外观形态和生理指标性状发生群体分化的可能性要高,群体间整体遗传差异的利用潜力较解剖结构大。同时,本研究发现叶片形状、叶片长宽比等18个性状在不同群体间达到显著或极显著差异,不同群体叶片表型性状的平均变异系数(CV)和多样性指数(H′)也不尽相同,李颖等[35]也认为板栗花序表型性状在不同群体间的遗传变异特征和均匀度不一致,说明板栗叶片表型不同群体间的差异和基因库价值存在不等值性,在相关研究或育种工作中除了重点调查个体基因型外,多样性较为丰富的群体可以作为种质资源创新利用的重点参考区域。
3.2 板栗叶片表型变异与地理生态因子间具有相关性
不同植物对环境因子的适应机制和敏感程度的差异会导致其性状呈现不同的地理变异规律,如苦楝()的叶宽、叶面积和果核长度等表型由南向北有增大的趋势[36-37],楠木()叶片由南向北表现出变狭长的变化趋势[27],高州油茶()的叶片长宽比与经度和纬度呈显著负相关[38]。本研究发现,板栗叶片36个表型性状中外观形态和生理指标性状与地理生态因子之间大多呈显著或极显著相关,总体而言,随着纬度的升高(由南向北),气温降低和降雨量减少,叶片表现出长宽比变小,锯齿深度、锯齿高度、叶绿素含量和叶片保水力变大的趋势,这可能是其在气温低、降雨量少的环境压力下为了提高光合效率或抗旱性而形成的适应性特征。另外,本研究中除了浙江和广西群体地理位置相隔较远但被聚为1类外(可能是由于这两个群体的试材由于长期引种驯化、变异导致与原产地的性状表现有所不同,或是群体试材数量相对较少,不足以代表各自群体多样性水平所致),板栗群体基本上按地理距离而聚类,由此得出板栗叶片大多数外观形态和生理性状呈现出以温度和降雨量为主导的梯度规律性。同时,本研究发现多数解剖结构性状与气候因子之间无显著相关性,这与Nicotra等[39]在天竺葵属()植物上的研究结果一致,说明板栗叶片解剖结构性状变异没有地理规律性,这可能是在长期的进化过程中,板栗上控制这些性状形成的基因片段比较稳定,其不易受外界环境影响而产生变异所致。
4 结论
板栗叶片表型在群体内和群体间均存在一定变异,但板栗叶片表型变异主要来源于群体内部,因此,以叶片表型性状作为研究对象,进行抗逆性、高光合效率等特性品种选育或相关研究时,研究重点应该放到广泛的个体基因型调查上,变异较为丰富均匀的群体可以作为重点参考。另外,板栗叶片大多外观形态和生理指标性状呈现以温度和降雨量因子为主导的梯度规律性。
[1] 徐永杰, 韩华柏, 王滑, 陈凌娜, 马庆国, 裴东.大巴山区核桃实生居群的坚果表型和遗传多样性.林业科学, 2016, 52(5): 111-119.doi:10.11707/j.1001-7488.20160513.
XU Y J, HAN H B, WANG H, CHEN L N, MA Q G, PEI D.Phenotypic and genetic diversities of nuts of walnut() populations originated from seedlings in Daba mountains.Scientia Silvae Sinicae, 2016, 52(5): 111-119.doi: 10.11707/j.1001-7488.20160513.(in Chinese)
[2] WANI G A, SHAH M A, TEKEU H, reshi z a, ATANGANA A R, KHASA D P.Phenotypic variability and genetic diversity of phragmites australis in Quebec and Kashmir reveal contrasting population Structure.Plants, 2020, 9(10): 1392.
[3] 程诗明, 顾万春.苦楝表型性状梯度变异的研究.林业科学, 2006, 42(5): 29-35.doi:10.3321/j.issn: 1001-7488.2006.05.006.
CHENG S M, GU W C.Studies on phenotypical characteristics gradient variation of.Scientia Silvae Sinicae, 2006, 42(5): 29-35.doi: 10.3321/j.issn: 1001-7488.2006.05.006.(in Chinese)
[4] TAKENAKA A.Effects of leaf blade narrowness and petiole length on the light capture efficiency of a shoot.Ecological Research, 1994, 9(2): 109-114.doi: 10.1007/BF02347485.
[5] MAGUYLO K, BASSETT C.Phenotypingcollections from Kazakhstan for leaf traits and tree architecture.Acta Horticulturae, 2014, 1058: 335-341.
[6] 李珊, 甘小洪, 憨宏艳, 许宁, 侯正扬, 陈杨, 邓晓娟.濒危植物水青树叶的表型性状变异.林业科学研究, 2016, 29(5): 687-697.doi: 10.3969/j.issn.1001-1498.2016.05.010.
LI S, GAN X H, HAN H Y, XU N, HOU Z Y, CHEN Y, DENG X J.Leaf phenotypic traits of, an endangered plant species.Forest Research, 2016, 29(5): 687-697.doi: 10.3969/j.issn.1001-1498.2016.05.010.(in Chinese)
[7] 崔思明, 郝亚涵, 周玮, 任颖, 周鹏, 陈晓阳.构树不同种源叶性状变异研究.中南林业科技大学学报, 2020, 40(5): 104-110.doi:10.14067/j.cnki.1673-923x.2020.05.013.
CUI S M, HAO Y H, ZHOU W, REN Y, ZHOU P, CHEN X Y.Variation of leaf morphological characters offrom different provenances.Journal of Central South University of Forestry & Technology, 2020, 40(5): 104-110.doi: 10.14067/j.cnki.1673-923x.2020.05.013.(in Chinese)
[8] BACILIERI R, DUCOUSSO A, KREMER A.Genetic, morphological, ecological and phenological differentiation between(MATT.) LIEBL.andL.in a mixed stand of Northwest of France.Silvae Genetica, 1995, 44(1): 1-10.
[9] 曾杰, 郑海水, 甘四明, 白嘉雨.广西西南桦天然居群的表型变异.林业科学, 2005, 41(2): 59-65.doi:10.3321/j.issn: 1001-7488.2005.02.010.
ZENG J, ZHENG H S, GAN S M, BAI J Y.Phenotypic variation in natural populations ofin Guangxi, China.Scientia Silvae Sinicae, 2005, 41(2): 59-65.doi: 10.3321/j.issn: 1001-7488.2005.02.010.(in Chinese)
[10] 尚帅斌, 郭俊杰, 王春胜, 赵志刚, 曾杰.海南岛青梅天然居群表型变异.林业科学, 2015, 51(2): 154-162.doi: 10.11707/j.1001-7488.20150219.
SHANG S B, GUO J J, WANG C S, ZHAO Z G, ZENG J.Phenotypic variations in natural populations ofin Hainan, China.Scientia Silvae Sinicae, 2015, 51(2): 154-162.doi: 10.11707/ j.1001-7488.20150219.(in Chinese)
[11] 李晓屿, Ralf Müller-Xing, Shahid Khan, 郑玉彩, 李玉花, 邢倩.植物叶缘锯齿发育的研究进展.中国农学通报, 2019, 35(1): 50-56.
LI X Y, MÜLLERXING R, KHAN S, ZHENG Y C, LI Y H, XING Q.Development of plant leaf margin: advances in research.Chinese Agricultural Science Bulletin, 2019, 35(1): 50-56.(in Chinese)
[12] 徐扬, 陈小红, 赵安玖.川西高原4种苹果属植物叶片解剖结构与其抗旱性分析.西北植物学报, 2015, 35(11): 2227-2234.doi:10.7606/j.issn.1000-4025.2015.11.2227.
XU Y, CHEN X H, ZHAO A J.Drought resistance evaluation and leaf anatomical structures of four species ofplants in western Sichuan plateau.Acta Botanica Boreali-Occidentalia Sinica, 2015, 35(11): 2227-2234.doi:10.7606/j.issn.1000-4025.2015.11.2227.(in Chinese)
[13] 张深梅, 奚建伟, 洪俊彦, 夏国华, 李岩, 黄兴召, 朱先富, 黄坚钦.大别山山核桃果实与叶片性状的表型多样性研究.林业科学研究, 2020, 33(1): 152-161.doi: 10.13275/j.cnki.lykxyj.2020.01.020.
ZHANG S M, XI J W, HONG J Y, XIA G H, LI Y, HUANG X Z, ZHU X F, HUANG J Q.A study on phenotypic diversity of fruit and leaf traits in.Forest Research, 2020, 33(1): 152-161.doi: 10.13275/j.cnki.lykxyj.2020.01.020.(in Chinese)
[14] 张莹, 曹玉芬, 霍宏亮, 徐家玉, 田路明, 董星光, 齐丹, 张小双, 刘超, 王立东.基于枝条和叶片表型性状的梨种质资源多样性.中国农业科学, 2018, 51(17): 3353-3369.doi: 10.3864/j.issn.0578-1752.2018.17.010.
ZHANG Y, CAO Y F, HUO H L, XU J Y, TIAN L M, DONG X G, QI D, ZHANG X S, LIU C, WANG L D.Diversity of pear germplasm resources based on twig and leaf phenotypic traits.Scientia Agricultura Sinica, 2018, 51(17): 3353-3369.doi: 10.3864/j.issn.0578-1752.2018.17.010.(in Chinese)
[15] 经建永, 颉刚刚, 欧阳丽婷, 陈曦, 马百强, 耿文娟.新疆野生欧洲李表型性状多样性分析.植物资源与环境学报, 2020, 29(2): 28-37.doi: 10.3969/j.issn.1674-7895.2020.02.04.
JING J Y, XIE G G, OUYANG L T, CHEN X, MA B Q, GENG W J.Analysis on diversity of phenotypic traits of wildin Xinjiang.Journal of Plant Resources and Environment, 2020, 29(2): 28-37.doi: 10.3969/j.issn.1674-7895.2020.02.04.(in Chinese)
[16] 王广鹏, 陆凤勤, 孔德军.板栗高效栽培技术与主要病虫害防治.北京: 中国农业出版社, 2016.
WANG G P, LU F Q, KONG D J.Highly Effective Cultivation and Control Technique of Main Pests and Diseases of Chestnut.Beijing: Chinese Agriculture Press, 2016.(in Chinese)
[17] 周连第, 兰彦平, 曹庆昌, 李淑英, 兰卫宗.板栗叶片性状表型多样性研究.林业科学, 2005, 21(9) :136-139.
ZHOU L D, lan Y P, Cao Q C, Li S Y, Lan W Z.Geographical variation of morphologic characteristics ofseeds and legumes.Scientia Silvae Sinicae, 2005, 21(9):136-139.(in Chinese)
[18] 江锡兵, 龚榜初, 刘庆忠, 陈新, 吴开云, 邓全恩, 汤丹.中国板栗地方品种重要农艺性状的表型多样性.园艺学报, 2014, 41(4): 641-652.
JIANG X B, GONG B C, LIU Q Z, CHEN X, WU K Y, DENG Q E, TANG D.Phenotypic diversity of important agronomic traits of local cultivars of Chinese chestnut.Acta Horticulturae Sinica, 2014, 41(4): 641-652.(in Chinese)
[19] 刘亚斌, 郭素娟.迁西地区39份板栗种质叶片的功能性状多样性.东北林业大学学报, 2020, 48(4): 8-14, 19.doi:10.3969/j.issn.1000- 5382.2020.04.002.
LIU Y B, GUO S J.Leaf functional traits of 39 chestnut germplasms in Qianxi region.Journal of Northeast Forestry University, 2020, 48(4): 8-14, 19.doi: 10.3969/j.issn.1000-5382.2020.04.002.(in Chinese)
[20] 刘庆忠.板栗种质资源描述规范和数据标准.北京: 中国农业出版社, 2006.
LIU Q Z.Descriptors and Data Standard for Chestnut (Blume).Beijing: Chinese Agriculture Press, 2006.(in Chinese)
[21] 国家林业局.植物新品种特异性、一致性、稳定性测试指南板栗: LY/T 1851—2009.北京:中国标准出版社,2009.
State Forestry Bureau.Guidelines for the conduct of tests for distinctness, uniformity and stability-Chestnut (Bl.): LY/T 1851-2009.Beijing: Standards Press of China, 2009.(in Chinese)
[22] 郭燕, 张树航, 李颖, 张馨方, 王广鹏.中国板栗238份品种(系)叶片形态、解剖结构及其抗旱性评价.园艺学报, 2020, 47(6): 1033-1046
GUO Y, ZHANG S H, LI Y, ZHANG X F, WANG G P.Studies on the leaf morphology, anatomical structure and drought resistance evaluation of 238 Chinese chestnut varieties (strains).Acta Horticulturae Sinica, 2020, 47(6): 1033-1046.(in Chinese)
[23] 孙志超, 程慧, 郭文磊, 王红红, 王正加.山核桃枝叶耐旱特性分析.果树学报, 2015, 32(4): 633-640.doi:10.13925/j.cnki.gsxb.20150068.
SUN Z C, CHENG H, GUO W L, WANG H H, WANG Z J.Analysis of drought resistance of branch and leaf structure in.Journal of Fruit Science, 2015, 32(4): 633-640.doi: 10.13925/j.cnki.gsxb.20150068.(in Chinese)
[24] 张小全, 许志文, 阚洪赢, 张林, 张鋆鋆, 郭传滨, 杨立均, 夏宗良.不同叶位和离体失水时间对烤烟叶片保水能力的影响.中国烟草学报, 2017, 23(4): 33-39.doi: 10.16472/j.chinatobacco.2017.093.
ZHANG X Q, XU Z W, KAN H Y, ZHANG L, ZHANG Y Y, GUO C B, YANG L J, XIA Z L.Effects of stalk positions and natural dehydration after priming on water retention capacity of flue-cured tobacco leaves.Acta Tabacaria Sinica, 2017, 23(4): 33-39.doi: 10.16472/j.chinatobacco.2017.093.(in Chinese)
[25] 舒展, 张晓素, 陈娟, 陈根云, 许大全.叶绿素含量测定的简化.植物生理学通讯, 2010, 46(4): 399-402.
SHU Z, ZHANG X S, CHEN J, CHEN G Y, XU D Q.The Simplification of chlorophyll content measurement.Plant Physiology Communications, 2010, 46(4): 399-402.(in Chinese)
[26] 李洪果, 陈达镇, 许靖诗, 刘光金, 庞晓东, 叶金辉, 莫小文, 谌红辉.濒危植物格木天然种群的表型多样性及变异.林业科学, 2019, 55(4): 69-83.doi:10.11707/j.1001-7488.20190408.
LI H G, CHEN D Z, XU J S, LIU G J, PANG X D, YE J H, MO X W, CHEN H H.Phenotypic diversity and variation in natural populations of, an endangered plant species.Scientia Silvae Sinicae, 2019, 55(4): 69-83.doi: 10.11707/j.1001-7488.20190408.(in Chinese)
[27] 欧汉彪, 林建勇, 李娟, 梁瑞龙.不同种源楠木叶片表型性状变异分析.西南农业学报, 2020, 33(3): 637-644.doi: 10.16213/j.cnki.scjas.2020.3.026.
OU H B, LIN J Y, LI J, LIANG R L.Analysis of variation of leaf phenotypic traits in different provenances of.Southwest China Journal of Agricultural Sciences, 2020, 33(3): 637-644.doi: 10.16213/j.cnki.scjas.2020.3.026.(in Chinese)
[28] 张云跃, 马常耕, 林睦就, 李柏海.我国银杏遗传变异研究之一: 种核性状的群体间和群体内变异.林业科学, 2001, 37(4): 35-40.doi:10.3321/j.issn: 1001-7488.2001.04.006.
ZHANG Y Y, MA C G, LIN M J, LI B H.Study on one of genetic variations for ginkgo biloba in China the var iation of breeding fruit-stone characters among and within population.Scientia Silvae Sinicae, 2001, 37(4): 35-40.doi: 10.3321/j.issn: 1001-7488.2001.04.006.(in Chinese)
[29] 聂兴华, 郑瑞杰, 赵永廉, 曹庆芹, 秦岭, 邢宇.利用荧光SSR分子标记评估中国栗属植物遗传多样性.中国农业科学, 2021, 54(8): 1739-1750.doi: 10.3864/j.issn.0578-1752.2021.08.013.
NIE X H, ZHENG R J, ZHAO Y L, CAO Q Q, QIN L, XING Y.Genetic diversity evaluation ofin China based on fluorescently labeled SSR.Scientia Agricultura Sinica, 2021, 54(8): 1739-1750.doi: 10.3864/j.issn.0578-1752.2021.08.013.(in Chinese)
[30] 田华, 康明.李丽, 姚小洪, 黄宏文.中国板栗自然居群微卫星(SSR)遗传多样性.生物多样性, 2009, 17(3): 296-302.
TIAN H, KANG M, LI L, YAO X H, HUANG H W.Genetic diversity in natural populations ofinferred from nuclear SSR markers.Biodiversity Science, 2009, 17(3): 296-302.(in Chinese)
[31] 张馨方, 张树航, 李颖, 郭燕, 王广鹏.基于SSR标记的板栗种质资源遗传多样性分析.分子植物育种, 2020, 18(15): 5164-5175.doi: 10.13271/j.mpb.018.005164.
ZHANG X F, ZHANG S H, LI Y, GUO Y, WANG G P.Genetic diversity analysis ofgermplasm resources based on SSR markers.Molecular Plant Breeding, 2020, 18(15): 5164-5175.doi: 10.13271/j.mpb.018.005164.(in Chinese)
[32] 谢一青, 黄儒珠, 李志真, 黄勇, 杨宗武.福建光皮桦野生种群遗传变异及其与生境的关系.林业科学, 2009, 45(9): 60-65.doi: 10.3321/j.issn: 1001-7488.2009.09.011.
XIE Y Q, HUANG R Z, LI Z Z, HUANG Y, YANG Z W.Genetic variation of natural populations ofin Fujian and its relationship with the habitat.Scientia Silvae Sinicae, 2009, 45(9): 60-65.doi: 10.3321/j.issn: 1001-7488.2009.09.011.(in Chinese)
[33] FUCHS E J, HAMRICK J L.Genetic diversity in the endangered tropical tree,(Zygophyllaceae).Journal of Heredity, 2010, 101(3): 284-291.doi: 10.1093/jhered/esp127.
[34] 白潇, 李毅, 苏世平, 赵小仙.不同居群唐古特白刺叶片解剖特征对生境的响应研究.西北植物学报, 2013, 33(10): 1986-1993.
BAI X, LI Y, SU S P, ZHAO X X.Response of leaf anatomical characteristics ofbobr.from different populations to habitats.Acta Botanica Boreali-Occidentalia Sinica, 2013, 33(10): 1986-1993.(in Chinese)
[35] 李颖, 张树航, 郭燕, 张馨方, 王广鹏.211份板栗种质资源花序表型多样性和聚类分析.中国农业科学, 2020, 53(22): 4667-4682.
LI Y, ZHANG S h, GUO Y, ZHANG X F, WANG G P.Catkin phenotypic diversity and cluster analysis of 211 Chinese chestnut germplasms.Scientia Agricultura Sinica, 2020, 53(22): 4667-4682.
[36] 陈丽君, 邓小梅, 丁美美.苦楝种源果核及种子性状地理变异的研究.北京林业大学学报, 2014, 36(1): 15-20.
CHEN L J, DENG X M, DING M M.Geographic variation intraits of fruit stones and seeds of.Journal of Beijing Forestry University, 2014, 36(1): 15-20.(in Chinese)
[37] 陈丽君, 刘明骞, 廖柏勇, 丁美美, 陈晓阳.苦楝不同种源叶片性状变异研究.西南林业大学学报, 2016, 36(5): 16-20.doi:10.11929/ j.issn.2095-1914.2016.05.003.
CHEN L J, LIU M Q, LIAO B Y, DING M M, CHEN X Y.Variation of leaf morphological characters offrom different provenances.Journal of Southwest Forestry University, 2016, 36(5): 16-20.doi: 10.11929/j.issn.2095-1914.2016.05.003.(in Chinese)
[38] 张鹏, 杨颖, 奚如春, 黄容容, 许叶.高州油茶种群叶片性状变异分析.林业与环境科学, 2018, 34(5): 13-19.
ZHANG P, YANG Y, XI R C, HUANG R R, XU Y.Leaf phenotypic variation in populations of.Forestry and Environmental Science, 2018, 34(5): 13-19.(in Chinese)
[39] NICOTRA A B, COSGROVE M J, COWLING A, SCHLICHTING C D, JONES C S.Leaf shape linked to photosynthetic rates and temperature optima in South Africanspecies.Oecologia, 2008, 154(4): 625-635.doi: 10.1007/s00442-007-0865-1.
Diversity Analysis of 36 Leaf Phenotypic Traits of Chinese Chestnut
GUO Yan, ZHANG ShuHang, LI Ying, ZHANG XinFang, WANG GuangPeng*
Changli Institute of Pomology, Hebei Academy of Agricultural and Forestry Sciences, Changli 066600, Hebei
【Objective】The objective of this study was to reveal the variation patterns and diversity characteristics of leaf phenotypic traits of Chinese chestnut, so as to provide the scientific basis for the protection, development and utilization of Chinese chestnut resources.【Method】Thirty-six leaf phenotypic traits (including 31 quantitative traits and 5 quality traits) of 240 Chinese chestnut germplasms from 10 provinces (populations) were measured.Variance analysis, multi-comparison, correlation analysis, and cluster analysis were used to explore the phenotypic diversity of chestnut leaf and the correlation between leaf phenotypic traits and climate factors.【Result】There were extremely significant differences (<0.01) in 31 leaf quantitative traits within the population, and there were significant or extremely significant differences in 15 quantitative traits (leaf length, leaf width, leaf length/width, sawtooth height, etc) and 3 quality traits (leaf shape, shape of leaf base, and leaf margin sawtooth direction) among populations, indicating the abundant variation among and within populations.The average of phenotypic differentiation coefficient among populations was 7.95%, which was much less than that within populations (92.05%), illustrating that the variation within populations was the main source of leaf phenotypic variation.The average variation coefficient (CV) and the Shannon-Wiener index (H′) of leaf phenotypic traits were 18.05% and 1.27, respectively.Among the five quality traits, the CV and H' of leaf shape were the highest (35.67%, 0.86), the CV and H' of leaf margin character were the lowest (5.60%, 0.07).The CVof 31 quantitative traits ranged from 8.68% (leaf length/width) to 32.73% (sawtooth depth), and the H′ranged from 1.54 (fuzz density on the back of leaf) to 2.09 (looseness of leaf tissue).The average H′of appearance, physiological and anatomical structure were 2.03, 2.04 and 2.04, respectively, and the average CV from high to low was 15.27% (appearance) >15.18% (physiological) >12.17% (anatomical structure).The average CV of leaf phenotypic traits in 10 populations ranged from 17.11% (Shaanxi) to 19.66% (Shandong), and the average H' in 10 populations ranged from 0.99 (Anhui) to 1.80 (Hebei).Most of the 36 leaf phenotypic traits of chestnut were significant or extremely significant correlated with geographical and ecological factors.The temperature indexes, such as annual mean temperature and average annual rainfall, were negatively correlated with leaf base shape, sawtooth height and other traits (longitude and latitude were positively correlated with them), and were positively correlated with leaf length/width (longitude and latitude were negatively correlated with them).【Conclusion】There were abundant variations in leaf phenotypic traits among and within populations, but the variation within populations was the main source of chestnut leaf phenotypic variation.Therefore, the research should focus on a wide range of surveys of individual genotypes, and the populations with relatively rich and uniform variation could be used as the key reference areas when the phenotypic traits of leaf were taken as the research object for breeding varieties with stress resistance and high photosynthetic efficiency or the related research.Most of the appearance and physiological characters of chestnut leafs showed gradient regularity dominated by temperature and rainfall factors.
chestnut; leaf phenotypic trait; among population; within population; variation pattern; phenotypic diversity
2021-05-19;
2021-08-04
河北省财政专项(F20R06)、河北省自然科学基金(C2020301053)
郭燕,E-mail:529127092@qq.com。通信作者王广鹏,E-mail:wangguangpeng430@163.com
(责任编辑 赵伶俐)
猜你喜欢
作物学报(2022年2期)2022-11-06
中国现代医生(2022年21期)2022-08-22
天天爱科学(2021年10期)2021-10-11
文萃报·周五版(2021年25期)2021-08-06
瞭望东方周刊(2021年6期)2021-03-30
快乐作文(1.2年级)(2021年12期)2021-03-21
三农资讯半月报(2020年2期)2020-03-09
含笑花(2019年6期)2019-12-09
农产品市场周刊(2015年36期)2015-10-30
意林·少年版(2009年7期)2009-07-08
