
黄地老虎幼虫取食诱导的棉花植株挥发物驱避雌蛾产卵
2022-04-19 13:41:54李梦宇夏施珂肖海军陆宴辉
昆虫学报 2022年3期
李梦宇, 张 涛, 夏施珂, 肖海军, 陆宴辉,*
(1. 江西农业大学昆虫研究所, 南昌 330045; 2. 中国农业科学院植物保护研究所, 植物病虫害生物学国家重点实验室, 北京 100193;3. 河北省农林科学院植物保护研究所, 河北保定 071000)
植物在受到植食性昆虫取食危害时会产生虫害诱导植物挥发物(herbivore-induced plant volatiles, HIPVs),这类挥发物常影响植食性昆虫同种个体的取食、产卵等选择行为。例如,很多植物被昆虫取食后会释放(E)-4,8-二甲基-1,3,7-壬三烯[(E)-4,8-dimethyl-1,3,7-nonatriene, DMNT)]作为直接或间接防御物质(Liuetal., 2018)。草地贪夜蛾Spodopterafrugiperda初孵幼虫喜欢利用DMNT搜索寄主植物,当添加过DMNT的豇豆幼苗与未受损豇豆幼苗及机械损伤的豇豆幼苗相比较时,草地贪夜蛾的初孵幼虫对添加DMNT的植株有更明显的偏好(Carrolletal., 2008)。在很多蛾类昆虫中,幼虫危害寄主产生的HIPVs对同种雌蛾具有显著的产卵忌避作用。烟草植株被烟芽夜蛾Heliothisvirescens幼虫取食后,释放的HIPVs能对雌蛾产卵行为产生显著影响(De Moraesetal., 2001)。棉花受植食性昆虫危害后,也能产生HIPVs。如被斜纹夜蛾Spodopteralittoralis取食的棉株能产生10余种能引起雌蛾触角响应的HIPVs,而且在健康棉花植株上不断添加HIPVs后,会明显降低雌蛾产卵量(Zakiretal., 2013)。因其产卵驱避作用,HIPVs有望开发为新型的害虫防控手段(Waretal., 2011)。
黄地老虎Agrotissegetum隶属于鳞翅目(Lepidoptera)夜蛾科(Noctuidae)地夜蛾属Agrotis,是一种重要的地下害虫,主要危害棉花、玉米及蔬菜等的幼苗和低矮草本植物,在我国多地均有分布(王敬儒等, 1983; 郭江龙等, 2016)。黄地老虎雌蛾平均产卵300~600粒,产卵量大且分散(屈年华, 2011)。雌蛾主要将卵产在低矮植物的叶片背面或杂草中,也会产在湿润的土表、土块等地方;其产卵方式会随产卵植物和气候条件等不同产生差异(王敬儒等, 1983; 李国英, 2017)。幼虫孵化后2 h即开始取食,危害植物幼苗及幼根;幼虫共6龄,2龄以下危害植物幼苗,3龄以上潜入土中,危害植物根茎;幼虫白天潜入土中,夜晚再钻出危害。由于幼虫在土壤中危害的隐蔽性,因此对棉花产量影响极大,尤其以第一代幼虫在新疆棉花苗期及蕾期危害最为严重(马祁等, 2000; 李新辉等, 2003; 李国英, 2017; 王盼盼等, 2017)。
本研究通过产卵笼试验评价黄地老虎雌蛾对幼虫取食危害过的棉花植株的产卵选择行为,用气相色谱-质谱联用仪(GC-MS)鉴定黄地老虎幼虫危害后棉花植株产生的HIPVs,通过触角电位仪(EAG)和室内产卵笼试验测试HIPVs组分激发雌蛾的电生理反应及对产卵行为影响,从而阐明黄地老虎幼虫危害棉花植株产生的HIPVs及其对雌蛾产卵选择的影响效应。研究结果将为解析黄地老虎与棉花互作关系、研发黄地老虎行为调控剂提供科学依据。
1 材料与方法
1.1 材料
1.1.1供试昆虫:黄地老虎为中国农业科学院植物保护研究所廊坊科研中试基地饲养的实验种群,于养虫室内用人工饲料进行继代饲养。饲养环境为温度26±1℃,相对湿度60%±5%,光周期14L∶10D。试验选择健康活跃的2-3龄幼虫用于危害棉花植株,健康活跃且无畸形的3-6日龄成虫进行产卵选择试验及触角电位测定。
1.1.2供试植物:供试棉花品种为中棉49,使用上口直径为8.5 cm、高11 cm的塑料花盆栽培于中国农业科学院植物保护研究所廊坊科研中试基地温室内,用营养土、蛭石及大田土以1∶3∶8的比例混匀培育,生长期间利用防虫网隔绝各种害虫的取食危害。选择健康的3-4叶期、株高25~30 cm的棉花植株进行试验。
1.1.3试剂:试验所需的标准化合物如表1所示。
表1 标准化合物的相关信息
1.2 黄地老虎雌蛾对幼虫危害棉花植株的产卵选择行为
根据孙小旭等(2018)的方法,设计制作了35 cm×35 cm×105 cm的产卵笼,产卵笼分为3个区域,两端为反应区,中间为释放区。试验时将产卵笼两端用纱布封住,剩余四周用保鲜膜包围并使其平整光滑。
在产卵笼一端反应区底部对角线中心位置放置1盆健康棉花植株(1株),另一端反应区底部对角线中心位置放置1盆土,两端土的重量相等。产卵笼释放区底部对角线中心位置放一个直径约2 cm、蘸有10%蜂蜜水的棉球,用于提供成虫补充营养。20∶00时在每个产卵笼中引入10头黄地老虎抱卵雌虫,24 h后检查在产卵笼不同部位的产卵数量。在产卵笼释放区产卵视为没有作出产卵选择,在两端反应区产卵视为作出产卵选择。试验过程中,用黑布遮盖产卵笼以消除周围光照的影响,每次更换试验处理前同时更换保鲜膜防止味道吸附而对试验产生干扰。试验在5个产卵笼中同时进行,共计5个重复。
上述预实验后,开展黄地老虎雌蛾对健康和受害棉花植株产卵选择性评价。在单盆1株健康棉花植株上接上4头饥饿4 h的2-3龄幼虫,叶片、植株基部各接2头。取食12 h后,受害棉花植株用于产卵选择试验。采用与上述相同的程序和方法,在产卵笼两侧分别放置1盆健康棉花植株和1盆被黄地老虎幼虫危害的棉花植株,接入10头黄地老虎抱卵雌虫,24 h后调查雌蛾在两端反应区的产卵量。试验在5个产卵笼中同时进行,共计5个重复。
1.3 黄地老虎幼虫危害棉花植株HIPVs的收集与分析
采用顶空法收集棉花植株挥发物,为了确保产生并收集到高浓度的挥发物,每盆保留3株棉花,将12头饥饿4 h的2-3龄幼虫分别接到3株棉花上,每株棉花叶片、植株基部各接2头。取食12 h后,用锡箔纸包裹盆土,将整盆棉花植株放入20 L玻璃罐中。采用大气采样仪(北京市劳动保护科学研究所,QC-1B型)抽气(500 mL/min),使空气经活性炭净化后进入玻璃罐,再经装有50 mg吸附剂(Porapak Q, 80-100 Mesh, 美国)的吸附柱。连续收集8 h后,用1 000 μL正己烷将收集的挥发物淋洗至进样瓶,然后氮吹浓缩至100 μL,保存于-20℃。每个处理重复3次。
利用GC-MS分析植物挥发物组分,GC-MS为安捷伦公司7890A-5975C,色谱柱为DB-wax(30 m×0.25 mm×0.25 μm)。GC进样口230℃,升温程序为柱箱初温50℃保持1 min;以5℃/min的速率升温至180℃,保持2 min;然后10℃/min升温至230℃,保持2 min。载气是氦气,流速是1 mL/min。MS离子源温度设定为230℃。所有挥发物组分均采用NIST14谱库检索初步鉴定,并通过与标准化合物保留时间比对进行确认,进一步用外标法对所有挥发物含量进行定量分析。通过健康棉花植株和受害棉花植株中各种挥发物组分含量对比,确定HIPVs的主要组分。
1.4 黄地老虎雌蛾对HIPVs组分的电生理反应
采用EAG测定3-6日龄活跃的黄地老虎雌蛾对HIPVs的电生理反应,EAG设备(荷兰Syntech公司)由探头数据采集控制器IDAC-2、触角固定器PRG-2、电极固定器、气味刺激控制装置CS-55、触角电位记录显示输出装置组成。将整个触角完整剪下,在解剖镜下切除触角末端部造成伤口。将触角基部连接到参比电极,端部连接到记录电极。触角电极采用填有电生理盐水(0.9%的氯化钠水溶液)的毛细管玻璃电极,电极内银丝连接PRG-2。
将标准化合物按照0.001, 0.01, 0.1, 1和10 μg/μL浓度配制。每次取10 μL不同浓度标准化合物滴到滤纸(0.5 cm×4 cm)上,将其塞入巴斯德管中,进行EAG测定。每次刺激时间为0.5 s,每2次刺激间隔30 s。以顺-3-己烯-1-醇为参照物评价黄地老虎对各个HIPVs组分的EAG反应,所有处理均重复10次。
1.5 黄地老虎雌蛾对具有电生理活性HIPVs组分的产卵选择行为
将对触角激发强烈电生理反应的HIPVs组分逐一开展产卵选择试验,操作同1.2节。在产卵笼两端各放置一盆土,然后将10 μL浓度为10 μg/μL的标准化合物滴到滤纸上放在处理端盆土上,对照端盆土上放置滴了10 μL石蜡油的滤纸,接入10头黄地老虎抱卵雌虫,24 h后调查雌蛾在两端反应区的产卵量。试验在5个产卵笼中同时进行,共计5个重复。
1.6 数据分析
计算EAG平均相对反应值,评价黄地老虎雌蛾对各HIPVs组分的电生理反应。 EAG相对反应值=(CT-CK)/(CF-CK)。其中,CT为对待测样品的EAG反应值,CK为对对照石蜡油的平均EAG反应值,CF为对参照顺-3-己烯-1-醇的平均EAG反应值。
黄地老虎雌蛾产卵量和EAG反应值分别用t检验和单因素方差分析(one-way ANOVA)进行统计分析(GraphPad Prism 9.0.1)。
2 结果
2.1 黄地老虎雌蛾对棉花植株的产卵选择
无棉花植株的盆土一端黄地老虎产卵量为9.60±0.51粒,有棉花植株一端的产卵量为22.20±1.02粒,两者差异显著(t=11.05,df=8,P<0.001)(图1: A)。这表明,上述自制产卵笼适用于黄地老虎成虫产卵选择行为试验。
在健康和受害棉花植株之间,黄地老虎雌蛾更偏好在健康棉花植株一端产卵,产卵量为87.00±7.64粒,而在被幼虫取食过的棉花植株一端产卵量仅为13.20±1.99粒,健康棉株上的产卵量为受幼虫危害棉株上的6.59倍,两者之间差异极显著(t=14.63,df=8,P<0.001)(图1: B)。试验中发现,黄地老虎雌蛾多在盆边、纱布及保鲜膜上产卵,而不直接在棉花植株上产卵。

图1 黄地老虎雌蛾对棉花植株的产卵选择
2.2 黄地老虎幼虫危害棉花植株HIPVs组分鉴定
与健康植株相比,黄地老虎幼虫取食危害后,棉花植株中有8种挥发物含量发生了显著变化,分别为十四烷、苯甲醛、β-石竹烯、芳樟醇、十六烷、葎草烯及2种无法确定种类的化合物(图2)。对前面6种HIPVs组分进行了定量分析,发现β-石竹烯和葎草烯含量变化最大,分别增加了5.33倍和3.68倍,而十四烷含量增加了0.76倍,苯甲醛含量增加了0.68倍,十六烷含量增加了1.53倍,增幅较小;同时,芳樟醇仅在幼虫危害的棉花植株中被发现(表2)。

图2 健康棉花植株(B)和黄地老虎幼虫危害棉花植株(A)挥发物的GC FID图

表2 黄地老虎幼虫危害棉花植株中6种已知挥发物组分的含量变化
2.3 黄地老虎雌蛾对HIPVs组分的电生理反应
在2.2节鉴定到的6种HIPVs组分中,β-石竹烯(F=11.54,df=4, 45,P<0.001)、苯甲醛(F=205.30,df=4, 45,P<0.0001)和芳樟醇(F=56.73,df=4, 45,P<0.001)引起的雌蛾触角反应随浓度增大而增大,其中苯甲醛引起的反应值最高,芳樟醇次之,β-石竹烯引起的反应最弱(图3)。在苯甲醛浓度为10 μg/μL时,EAG相对反应值大于1,说明其引起的触角反应程度强于参照物顺-3-己烯-1-醇;同时,葎草烯(F=30.90,df=4, 45,P<0.001)、十四烷(F=3.28,df=4, 45,P=0.019)和十六烷(F=1.01,df=4, 45,P=0.413)引起的触角反应则并不随浓度增大而增大(图3)。

图3 黄地老虎雌蛾对不同HIPVs组分的触角电位反应
2.4 黄地老虎雌蛾对HIPVs组分的产卵选择行为
对2.3节中EAG活性最强的芳樟醇和苯甲醛,分别进行产卵选择试验。结果发现,与对照相比,黄地老虎雌蛾更偏好在苯甲醛一端产卵,对照端产卵量为15.40±3.03粒,苯甲醛端产卵量为71.00±23.67粒(t=2.33,df=8,P=0.048)(图4: A)。而芳樟醇则完全相反,雌蛾在对照一端产卵更多,对照端产卵量为94.20±14.95粒,芳樟醇端产卵量为46.60±11.69粒(t=2.508,df=8,P=0.037)(图4: B)。初步确认黄地老虎幼虫危害棉花植株产生的HIPVs中,具有产卵驱避活性的化合物主要是芳樟醇。
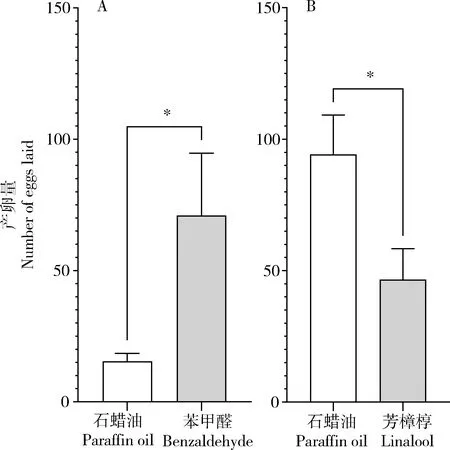
图4 黄地老虎雌蛾对芳樟醇和苯甲醛的产卵选择
3 讨论与结论
植物在生长过程中会面对各种植食性昆虫的侵害,在长期共同进化中植物对植食性昆虫形成了一套稳定的防御系统,其中释放HIPVs以驱避害虫取食及产卵等行为是植物防御的重要手段之一。植物HIPVs释放防御这一生物学现象在亚洲玉米螟Ostriniafurnacalis、二斑叶螨Tetranychusurticae和细纹黛齿螟Heortiavitessoides(Horiuchietal., 2003; Huangetal., 2009; Qiaoetal., 2018)等害虫危害中均有报道。本研究发现棉花植株被黄地老虎幼虫危害后也会产生类似的防御反应,能够通过释放HIPVs降低雌蛾在受害植株上的产卵量。这种产卵行为可能是源于避免同种不同个体间在同一植株上的生态位竞争。
本研究发现健康棉花植株几乎不释放芳樟醇,但是被黄地老虎幼虫危害后,棉花植株会产生芳樟醇(表2)。电生理实验发现,黄地老虎雌蛾对芳樟醇反应明显,且存在显著的剂量反应关系(图3);进一步行为实验表明,黄地老虎雌蛾在芳樟醇附近产卵量明显更少(图4)。因此,我们推断芳樟醇是棉花受到黄地老虎危害后产生的具有产卵驱避活性的HIPVs。作为一种重要的HIPVs,芳樟醇在其他植物受到危害时也会产生,并起到产卵驱避作用。例如,Kessler和Baldwin(2001)的研究表明,当烟草受到烟芽夜蛾H.virescens危害后芳樟醇产生的量会明显增多,进一步在田间试验利用人工合成的芳樟醇标准品模拟烟芽夜蛾危害烟草后的芳樟醇释放量,结果表明芳樟醇对烟芽夜蛾的产卵具有显著的驱避作用。棉花被斜纹夜蛾危害后会产生芳樟醇等气味,并进一步驱避斜纹夜蛾的产卵行为(Zakiretal., 2013)。为了更好发挥芳樟醇的产卵驱避作用,科学家拟通过转基因技术,让作物本身释放芳樟醇以驱避害虫。结果发现,当转芳樟醇基因烟草与正常烟草相对比时,能释放芳樟醇的转基因烟草上棉铃虫Helicoverpaarmigera的产卵量会显著减少(McCallumetal., 2011);桃蚜Myzuspersicae在转芳樟醇基因烟草植株上的产卵量显著少于非转基因烟草上的(Huangetal., 2018)。因此,芳樟醇是一种对多种害虫都有产卵驱避活性的HIPVs组分,对棉田生态系统中多种害虫的生态功能有待系统评价。
与健康棉花植株相比,受到黄地老虎危害的棉花植株会释放出6.33倍的β-石竹烯和4.68倍的葎草烯,同时会产生更多的苯甲醛、十四烷等物质(表2)。虽然这些挥发物在健康棉花中也存在,但是被害虫危害后释放量显著增加,我们也将之确认为黄地老虎危害棉花产生的HIPVs。其中,苯甲醛不仅能够引起黄地老虎雌蛾的强烈触角电位反应,而且会吸引黄地老虎雌蛾产卵。苯甲醛是常见的花香物质(Hoballahetal., 2005),在棉花、烟草、玉米等植物挥发物中均普遍存在(Loughrinetal., 1994; Yanetal., 2005; Yan and Wang, 2006),可吸引黄地老虎雌蛾前往觅食,从而导致苯甲醛端产卵量更多。本试验对含量变化不大及触角电生理反应小的物质没有进行重点评价,但是否对芳樟醇等功能挥发物存在影响效应,值得进一步探索。
本研究发现棉花植株受到黄地老虎幼虫危害后会显著降低雌蛾在受害植株的产卵量,其中芳樟醇是主要的产卵驱避活性物质,从而阐明黄地老虎幼虫危害棉花植株产生的HIPVs及其组分对雌蛾产卵选择的影响效应,结果为进一步开发基于HIPVs的黄地老虎防控技术提供了科学依据。
猜你喜欢
幼儿智力世界(2024年1期)2024-05-24 17:38:54
广州化工(2021年19期)2021-10-25 14:03:02
华人时刊(2020年19期)2021-01-14 01:17:20
能源(2018年5期)2018-06-15 08:56:20
金色少年(奇趣科普)(2017年4期)2017-06-05 15:03:48
中国调味品(2017年2期)2017-03-20 16:18:22
农村农业农民·B版(2016年7期)2016-10-21 10:44:23
广西林业科学(2016年2期)2016-03-20 05:53:43
经济林研究(2015年3期)2015-12-21 01:23:25
天然产物研究与开发(2014年8期)2014-04-27 14:16:30
