
基于环境DNA技术的夏季东海鱼类物种多样性研究
2022-04-18 10:16李晓玲刘洋王丛丛俞晔伟李纲
海洋学报 2022年4期
李晓玲,刘洋, 2, 3, 4, 5,王丛丛, 2, 3, 4, 5,俞晔伟,李纲, 2, 3, 4, 5
( 1. 上海海洋大学 海洋科学学院,上海 201306;2. 大洋渔业资源可持续开发教育部重点实验室,上海 201306;3. 上海海洋大学 国家远洋渔业工程技术研究中心,上海 201306;4. 农业农村部大洋渔业开发重点实验室,上海 201306;5. 农业农村部大洋渔业资源环境科学观测实验站,上海 201306)
1 引言
生物多样性在支撑生态系统稳定性方面发挥着重要作用,是实现全球可持续发展目标的重要基础[1],通常由遗传多样性、物种多样性和生态系统多样性3个部分组成,其中物种多样性是生物多样性的核心。海洋生态系统是地球上最大的水生生态系统,而鱼类是海洋生态系统的关键类群,也是人类的重要蛋白质来源[2]。在人类活动和全球气候变化的影响下,海洋鱼类物种多样性受到了严重威胁[3],包括大量物种的丧失以及濒危物种的增多,特别是人类活动较为频繁的一些陆缘海,情况更为严重。因此,监测鱼类物种多样性对此类典型海域的生态系统科学管理和资源可持续利用至关重要。
东海海域地处北太平洋西部,海岸线曲折,大陆架面积广阔,是一个较大的陆缘海[4]。由于独特的地理位置,许多河流,包括长江、钱塘江、瓯江等均汇入东海,此外,东海东南部受到黑潮暖流的影响,使得其初级生产力高、生物种类丰富、渔业资源量大。东海中有带鱼(Trichiurus japonicus)、小黄鱼(Larimichthys polyactis)、日本鲐(Scomber japonicus)等重要经济物种,形成了我国重要的渔场[5-6]。为了能够合理利用渔业资源,我国对东海海域的鱼类资源开展了大量调查,调查方法主要为底拖网调查[7-9],此方法往往耗时耗力,对生物体本身也会造成直接伤害。
为避免调查对生物系统的破坏,非侵略性调查方法将会成为生物调查的主流。近年来,环境DNA(Environmental DNA,eDNA)技术越来越多地被应用于生态调查[10-12]。eDNA是指从环境中而不是从生物个体中收集的DNA[13],比如鱼类释放到水中的体表碎片、黏液和排泄物等。而eDNA技术是指从环境样本(采集的海水)中直接提取DNA片段后利用PCR和测序技术进行定性或定量分析,从而确定目标生物在该环境中的分布的研究方法[14],研究过程中无需与研究生物直接接触。Sigsgaard等[15]通过eDNA技术检测到丹麦沿海29种鱼类,与浮潜观察所得结果有高度的一致性。Fraija-Fernández等[16]利用eDNA技术揭示了东北大西洋鱼类群落组成,且物种序列读数和生物量之间存在总体相关性。目前,国内对于eDNA的研究也日趋丰富,在象山港水域[17]和洱海[18]的两项研究中,通过eDNA序列丰度获得的优势鱼类物种和渔业资源调查结果基本一致。Wang等[19]基于eDNA浓度对东海小黄鱼进行渔业资源评价,发现其区域分布和水层分布与传统拖网捕捞结果一致。本研究利用eDNA技术初步分析东海海域鱼类物种的多样性,以期为鱼类物种的保护和合理利用以及东海海域生态系统的稳定性保护提供参考。
2 材料与方法
2.1 环境样本采集
本实验所用环境样本为2020年9月19-21日采自东海海域的海水样本。本次调查共在航行过程中依次选取14个站点(间隔50~160 km),具体采样站点见图1,甲板上采水器的下放点远离调查船出水口,每个站点采集5 L表层水。水样采集完毕立即在调查船实验室中使用滤膜对海水中的eDNA进行富集,所选滤膜为直径47 mm、孔径0.45 μm的尼龙膜(Nynon)。过滤所用器材提前经过稀释配置的84消毒液(有效氯含量为0.3%)浸泡消毒处理,过滤实验水样前先过滤3个空白水样,然后再过滤实验水样,用规格1 L的无菌可密封广口瓶量取过滤水样,每张滤膜过滤2.5 L,过滤后得到的滤膜用镊子卷曲放入1.5 mL棕色离心管中密封,标记站点和采样时间,-80℃冷冻保存至提取DNA。使用DNeasy Blood and Tissue Kit试 剂 盒(Qiagen, Hilden, Germany)进 行eDNA提取,提取方法参照试剂盒说明书并做适当调整,提取的eDNA混匀后分装,每管20 μL,并立即使用琼脂糖凝胶电泳法和Nanodrop分光光度计对其进行质量检测,记录对应样本eDNA测定浓度及OD260/280值(即260 nm波长和280 nm波长下吸光度的比值),剩余eDNA置于-80℃冰箱中保存备用。
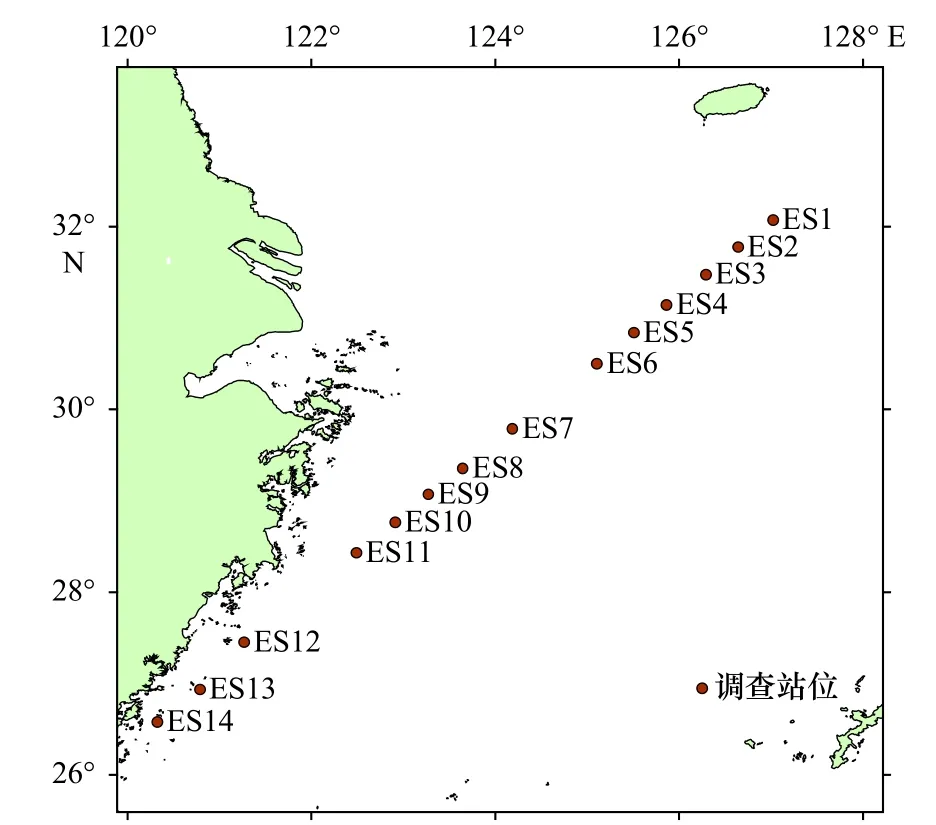
图1 2020年9月东海采样站点Fig. 1 Sampling stations in the East China Sea in September 2020
2.2 PCR扩增与测序
使用已报道的FISH eDNA通用引物MiFish-E-F:5′-GTTGGTAAATCTCGTGCCAGC-3′和MiFish-E-R:5′-CATAGTGGGGTATCTAATCCTAGTTTG-3′进 行PCR扩增,该引物针对12S rRNA基因的一个高变区[20]。扩增体系25 μL,包含5×反应缓冲液和5×GC 缓冲液各5 μL,dNTP(2.5 mmol/L)2 μL,正反向引物(10 μmol/L)各1 μL,模板DNA(20 ng/μL)2 μL,双蒸水8.75 μL,Q5DNA聚合酶0.25 μL。采用两步PCR方法制备成对末端文库,PCR扩增参数:95℃预变性5 min,95℃变性30 s,60℃退火30 s,然后进行55个循环。PCR扩增仪器为ABI 2720型PCR扩增仪。同时建立阴性对照,检测来自环境或试剂的微生物污染。在随后的实验中,任何有条带扩增的阴性对照组都没有被使用。最后,在Illumina MiSeq平台(委托上海派森诺生物科技有限公司)上对文库进行双端测序。
2.3 数据分析
测序原始数据以FASTQ格式保存,并根据序列质量进行初步筛查;对问题样本进行重测、补测。使用Vsearch(v2.13.4_linux_x86_64)、cutadapt(v2.3)软件对原始双端测序数据进行拼接、去重,再按照97%相似度水平聚类为可操作分类单元(Operational Taxonomic Unit,OTU),并获得代表序列和OTU表。代表OTU序列与参考序列数据库NCBI(https://www.ncbi.nlm.nih.gov/)和 MitoFish(http://mitofish.aori.u-tokyo.ac.jp/download.html)进行比对作物种分类学注释。统计各样本不同分类水平各自含有的分类单元的数量,并在种水平上对优势物种做进一步分析。使用QIIME2软件[21]进行α多样性分析,α多样性指数表征物种在生境内的多样性[22],其中Chao1指数[23]和观察到的物种数(Observed species )指数表征丰富度,Shannon指数[24-25]和Simpson指数[26]表征多样性,Pielou指数[27]表征均匀度,具体计算方法参考http://scikit-bio.org/docs/latest/generated/skbio.diversity.alpha.html#moduleskbio.diversity.alpha。基于各采样点物种序列丰度,利用R语言pheatmap包绘制物种组成热图。并使用R语言VennDiagram包绘制OTU花瓣图,分析不同采样点的共有OTU和独有OTU。
3 结果与分析
3.1 eDNA测序的结果
共测定14个样本,经原始双端测序数据处理,总共获得656 863条原始序列,其中高质量序列547 435条。eDNA样本的高通量测序统计结果如表1所示。
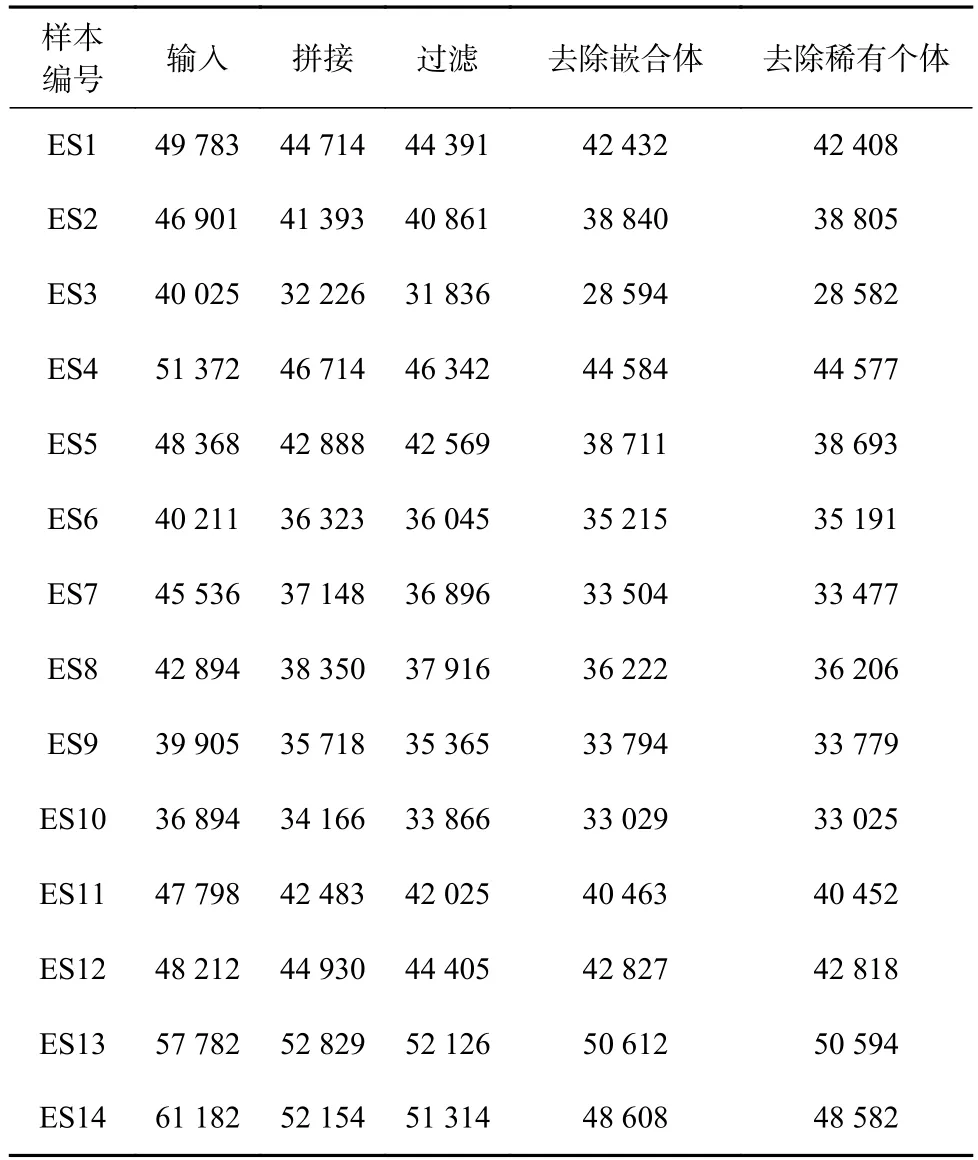
表1 各站点的序列量(eDNA)结果Table 1 Sequence quantity (eDNA) results of each station
3.2 鱼类物种组成
高质量序列最终聚成的OTU与数据库NCBI(https://www.ncbi.nlm.nih.gov/) 和MitoFish(http://mitofish.aori.u-tokyo.ac.jp/download.html)进行比对注释,并手动移除非鱼类的OTU,非鱼类的注释序列主要包括鸟类、两栖类和藻类。鱼类注释序列的OTU划分和分类地位鉴定结果如图2所示。筛选并删除淡水鱼类物种,共检测到11种淡水鱼类,主要为鲤形目鱼类,出现这种现象的原因很可能是样品出现污染,需对阴性对照进行测序分析,找到污染源头。共检测出海水鱼类2纲,23目,29科,42属,44种,具体结果如表2所示。

表2 东海环境DNA检出的鱼类物种Table 2 Fish species detected by eDNA in the East China Sea
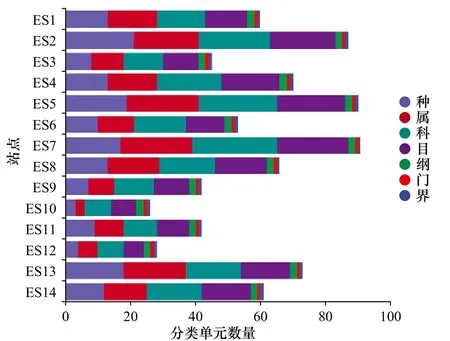
图2 东海鱼类OTU划分和分类地位鉴定结果Fig. 2 Result of OTU division and classification taxon identification of fish in the East China Sea
图3展示了利用eDNA分析技术检出的丰度前10位的鱼类优势物种,分别为赤鼻棱鳀(Thryssa kammalensis)、蓝点马鲛(Scomberomorus niphonius)、日本鲐(Scomber japonicus)、小黄鱼(Larimichthys polyactis)、鲻(Mugil cephalus)、日本鳀(Engraulis japonicus)、远东拟沙丁鱼(Sardinops melanostictus)、海鳗(Muraenesox cinereus)、七星底灯鱼(Benthosema pterotum)和龙头鱼(Harpadon nehereus)。其中赤鼻棱鳀、日本鳀和远东拟沙丁鱼均属于鲱形目(Clupeiformes),日本鲐和蓝点马鲛均属于鲭形目(Scombriformes)。
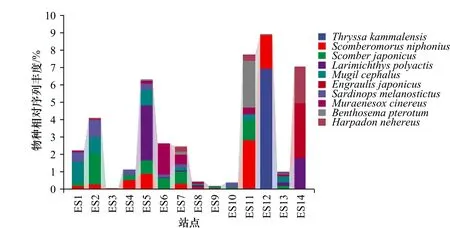
图3 各采样站点优势鱼类物种组成Fig. 3 The composition of dominant fish species at each sampling station
3.3 鱼类物种多样性分析
图4为本次测序所得的鱼类物种稀疏曲线,由图可见所有站点测序深度均达到了平台期,反映当前测序数据足够用于后续多样性分析。
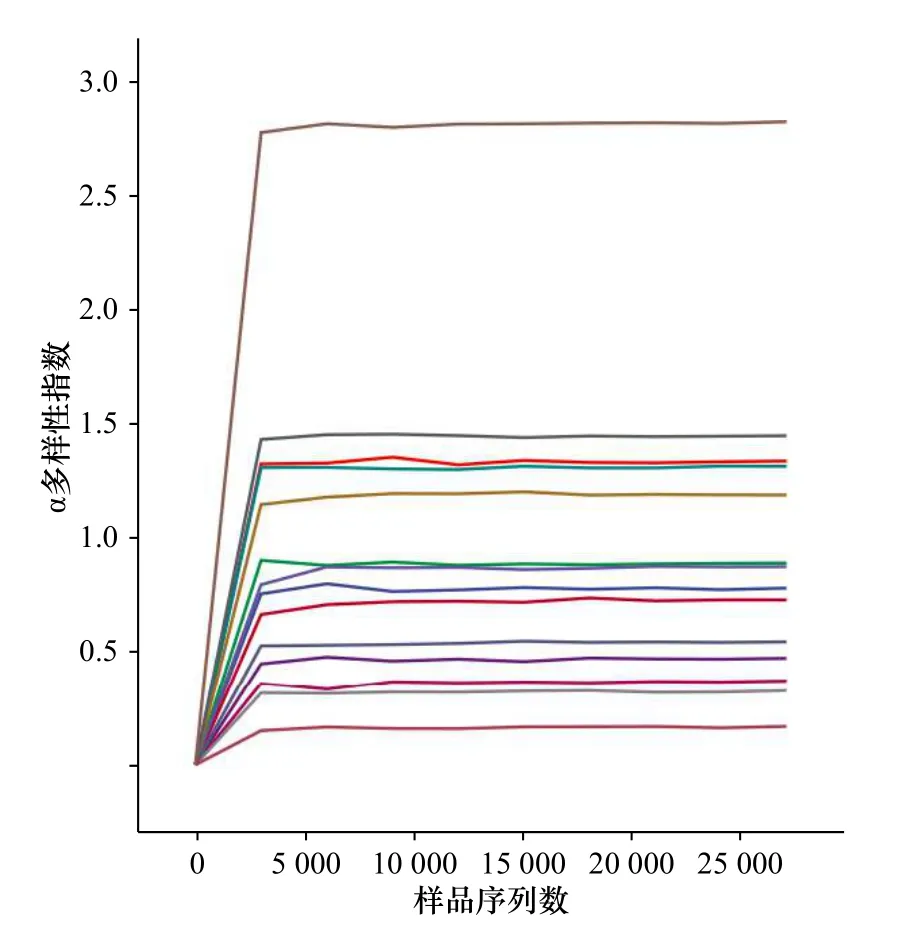
图4 东海不同采样站点(每条线代表一个站点)物种α多样性指数稀疏曲线Fig. 4 Sparse curve of species alpha diversity index at different sampling stations (each line represents a different station) in the East China Sea
表3所示为反映鱼类群落相对丰度的α多样性指数。其中,Chao1指数范围为52.628 ~180.581,Observed species指数范围为41.3~158.0,两个指数的分布趋势基本一致;Simpson多样性指数范围为0.028 676~0.644 819,Shannon多 样 性 指 数 范 围 为0.168 019~2.827 730,两者分布趋势也基本对应。Pielou均匀度指数范围为0.031 302~0.396 211。各个样品所测得的鱼类物种α多样性指数存在差异,其中ES12和ES14样品具有较高的Simpson和Shannon多样性指数,而ES5和ES7具有较高的Chao1和Observed species指数。
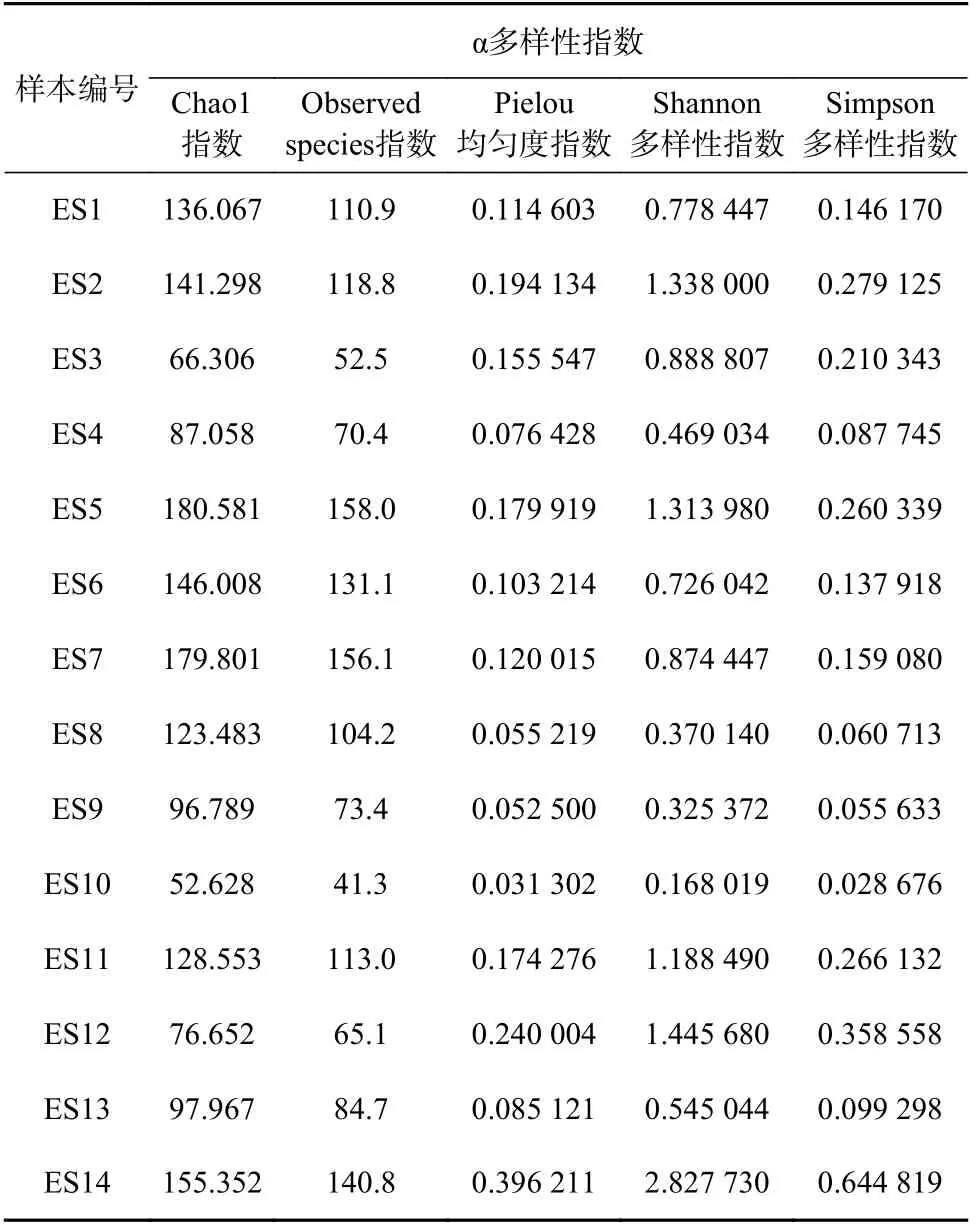
表3 东海鱼类物种相对丰度的α多样性指数Table 3 Alpha diversity index of relative abundance of fish species in the East China Sea

续表2
3.4 样本间鱼类物种差异分析
利用R软件根据获得的样本OTU统计表绘制了14个采样站点样本共有OTU的花瓣图,各样本中共有和特有OTU比例如图5a所示。由图可见,14个采样站点共有OTU所占比例较低,且ES14、ES5、ES6、ES2和ES7这5个采样点具有较高比例的特有OTU。为了比较不同海域鱼类物种的组成情况,按照离岸远近,将研究区域从空间上分为近岸、近海和远海3个部分,每个部分随机选取相同数量的站点进行分析,即ES12、ES13、ES14为近岸组,ES7、ES8、ES9为近海组,ES1、ES2、ES3分为远海组,3部分OTU比例如图5b所示,近海海域的特有OTU数量最多,近岸海域次之,远海海域最少。图6展示了每一优势鱼种在不同采样站点的相对丰度,其中,OTU检出频率较高的鱼类为蓝点马鲛、日本鲐、鲻和远东拟沙丁鱼,在14个采样站点中的检出频率均为0.7以上。值得注意的是,OTU相对丰度最高的物种赤鼻棱鳀在采样站点中的出现位置最为集中,为ES12站点,该站点位于福建、浙江沿海。
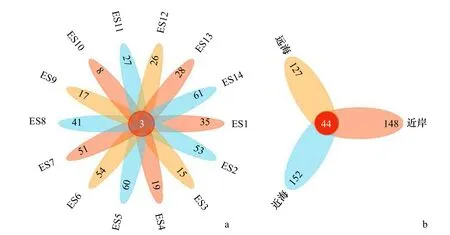
图5 东海水域不同采样站点共有OTU的花瓣图Fig. 5 Petal map of OTU shared by different sampling stations in the East China Sea
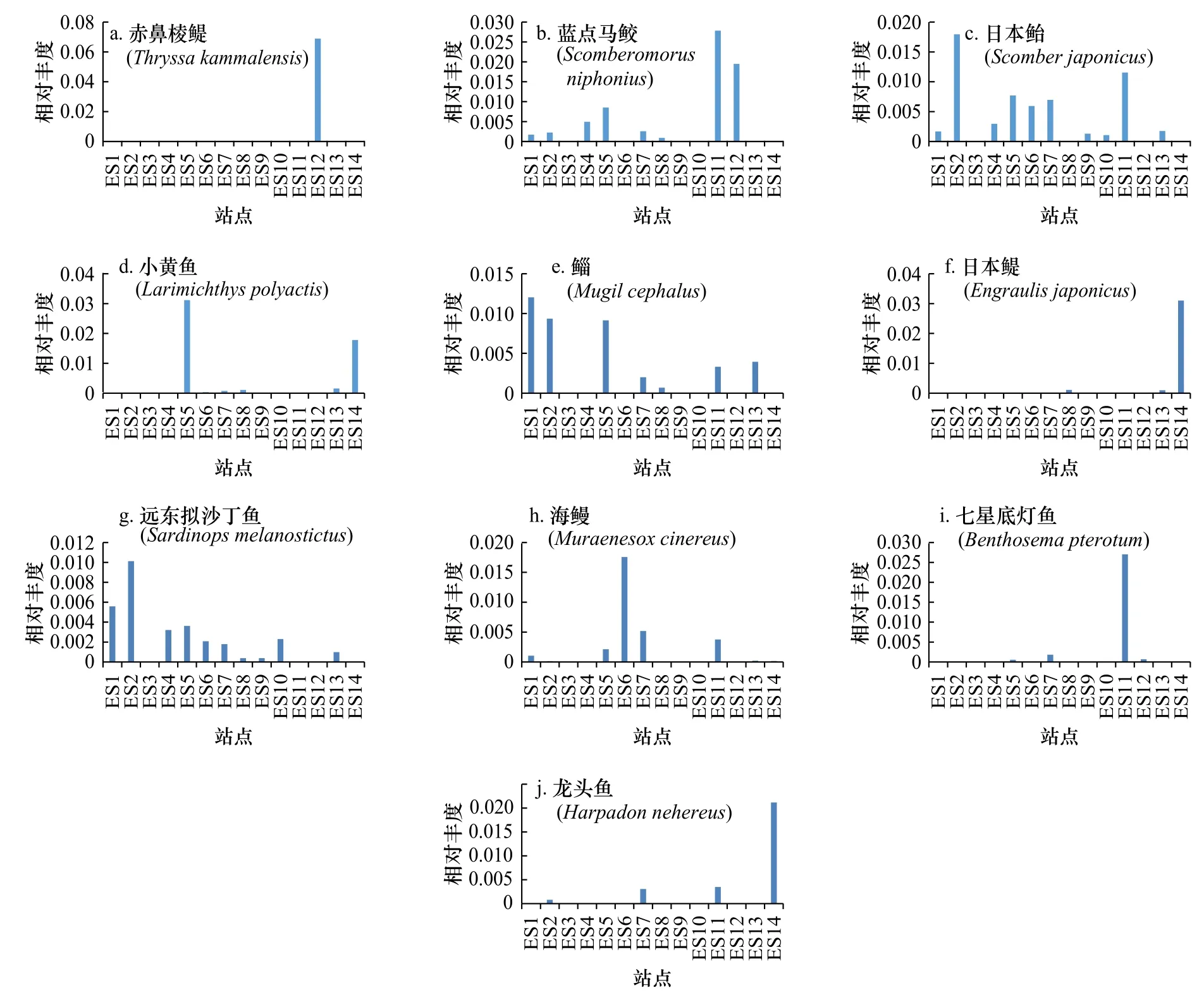
图6 各优势鱼种在不同采样站点的相对丰度Fig. 6 Relative abundance of dominant fish species at different sampling stations
利用R软件绘制基于种水平的鱼类组成热图(图7)。如图所示,不同物种的组成在各个站点(样本)间存在差异,从样本聚类上看,地理上相邻的站点ES9和ES10被最先聚在一起,而与ES9和ES10物种组成差异最大的为站点ES2,并不是地理上与ES9和ES10相距最远的站点。从物种聚类上看,闪电烛光鱼(Polyipnus stereope)和焦氏舌鳎(Cynoglossus joyneri)最先聚在一起,除了在站点ES2有较高的相对丰度,在其余站点的相对丰度均较低;鲻、日本鲐和蓝点马鲛均为相对丰度前10的优势物种且在各站点的组成较为相似。整体看来,多样性和丰富度的趋势与3.3节所述α多样性指数结果一致,其中ES5、ES2和ES14站点均有较高的多样性和丰富度。
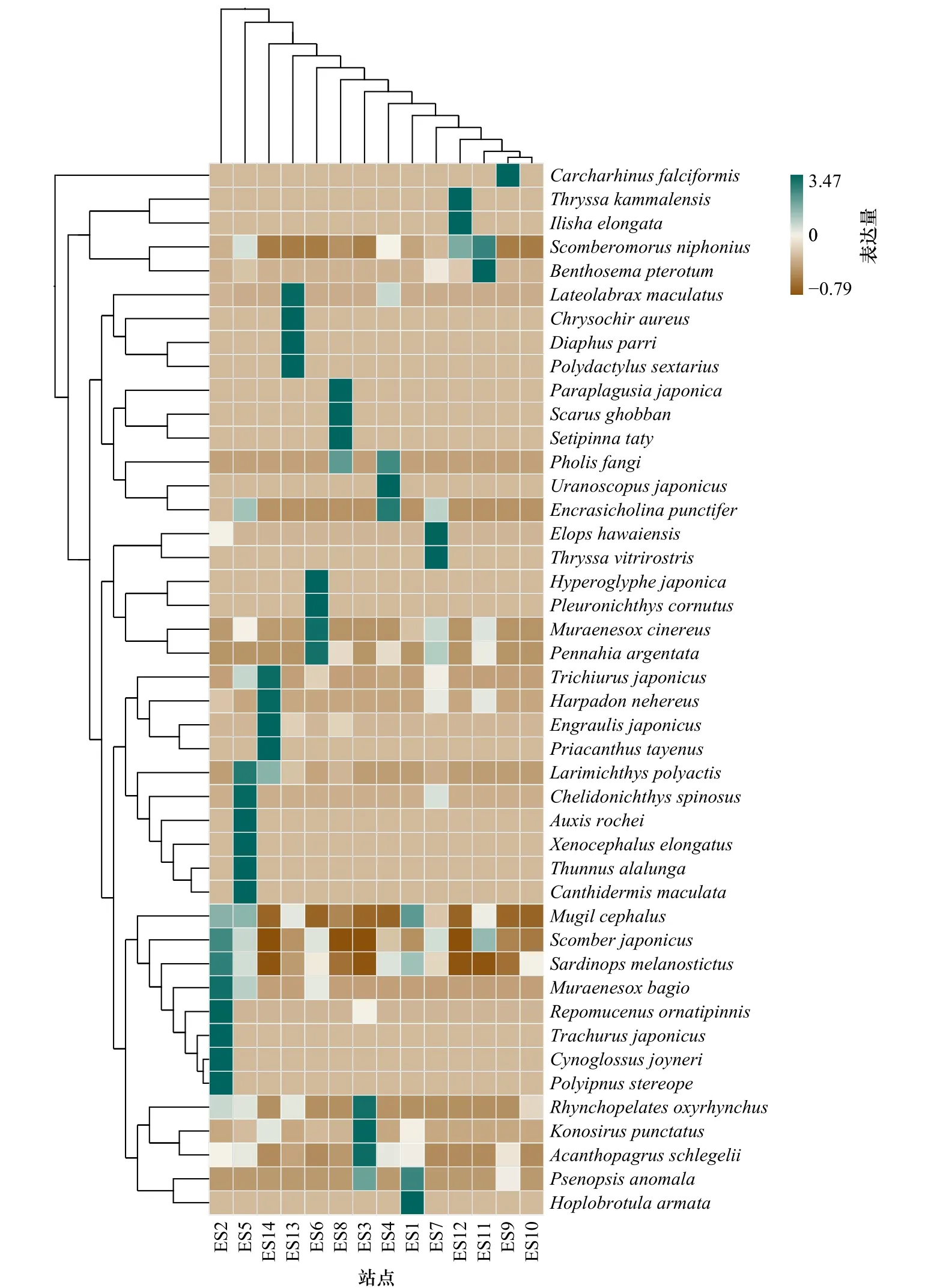
图7 环境DNA检出的东海鱼类物种组成热图Fig. 7 Heat map of fish species composition in the East China Sea detected by eDNA
4 讨论
东海海域海岸线曲折,浅滩面积广阔,渔业资源丰富,但近年来,由于人类活动(主要为过度捕捞)的影响,其传统渔业资源遭到严重破坏,如曾属于四大经济鱼类之一的大黄鱼(Larimichthys crocea)早已濒临灭绝,各大渔场的主要渔获物逐渐被年龄结构简单、经济价值低、个体小和营养级层次低的种类所替代[28]。因此,开展东海海域鱼类群落结构和多样性研究,可以为相关鱼类物种的保护和合理利用提供参考资料,进一步实现东海海域生态系统的科学管理。然而,东海传统的渔业资源调查主要为底拖网调查,不仅耗费大量人力物力,还会破坏海洋底质,进而破坏海底生态系统的稳定性,此外,渔获物的鉴定比较依赖鱼类专家,研究周期较长。本研究使用eDNA技术进行东海鱼类多样性研究,仅需采集少量海水样本即可,对生物较为友好,且对专业人员比如鱼类专家的要求不高,一定程度上降低了研究成本并减少了研究时间[29-30]。此外,对于一些密度较低的物种,eDNA技术与传统的研究方法相比更为灵敏和准确[12],其结果可作为传统方法的补充。
本研究在14个站点的海水样本中共检测出2纲,23目,29科,42属,44种海水鱼类(表2),大部分种类在东海传统渔业资源调查结果中均有出现。前人对东海海域进行底拖网渔业资源调查[7-8,31],发现多年来各季节主要的优势鱼类物种较为固定,9月份的优势物种主要为带鱼、小黄鱼、刺鲳(Psenopsis anomala)、日本鲐、龙头鱼(Harpadon nehereus)等,这些物种在本次取样的分析结果中也均有出现。此外,与东海传统渔业资源调查相比,存在一些未曾报道或报道较少的鱼类,如尖突吻䱨(Rhynchopelates oxyrhynchus)、黄吻棱鳀(Thryssa vitrirostris)、银灰半棱鳀(Encrasicholina punctifer)、尖头黄鳍牙鱛(Chrysochir aureus)、双 鳍 舵 鲣(Auxis rochei)、疣 鳞 鲀(Canthidermis maculata)等。其中一些鱼类通过传统的形态学方法进行鉴定或区分会存在一定的偏差,比如同属于棱鳀属(Thryssa)的黄吻棱鳀和赤鼻棱鳀,形态学上主要根据上颌骨末端的长度来区分[32],但长期生活在同一环境中可能会使得这一骨骼特征表现出高度的相似性[33],因此,仅根据形态学方法来区分近缘种有一定的局限性。
本次采样时间为2020年9月,环境DNA分析结果中的优势物种组成(图3,图6)与近20年传统调查的结果有所不同,比如2000年9月丰富度最高的带鱼以及丰富度第3的刺鲳[34]在本次结果中丰富度相对较低,未被列入优势物种,而本次环境DNA分析结果中丰富度前10的鱼类中赤鼻棱鳀和鲻在以往的东海传统渔业资源调查中丰富度较低或报道较少。这些差异的产生可能与目标物种的生活习性以及环境DNA在特定环境中的降解速率有关,比如传统调查中的优势物种带鱼,属暖水性中下层鱼类,具有昼夜垂直移动的习性,且昼沉夜浮[35],这一生活习性可能会影响到样品的采集质量,即白天采集的表层水样中所含有的带鱼环境DNA可能会相对较少,这需要在样品采集中适当增加采样深度或增加夜间采样量。而本次14个站点采样检测出的相对丰度最高的物种赤鼻棱鳀,仅存在于ES12站点,一方面说明采样季节ES12站点即福建、浙江沿海有丰富的赤鼻棱鳀资源,另一方面也说明单次采样所得到的数据并不全面,对于面积较大的研究海域,需要设计多个站点且同一季节多次采样,并将结果进行综合分析,从而减少个别样品差异对整个实验结果的影响。
本研究采用α多样性指数(表3)来表征东海海域的鱼类物种多样性,α多样性指数数值大小很大程度上由群落自身特点所决定,该指数的主要优势在于可反映多样性空间变化规律和时间尺度的变化趋势[36],包括丰富度、多样性和均匀度几个指标。Chao1和Observed species指数表征丰富度,14个站点中该指数在ES5和ES7站点数值较高,表明相比于其他站点具有较高的鱼类物种丰富度;Shannon和Simpson多样性指数表征多样性,14个站点中该指数在ES12和ES14站点数值较高,表明相比于其他站点具有较高的鱼类物种多样性,且这两个站点均离岸较近,大陆架浅滩有江河淡水汇入,初级生产力高,利于海洋鱼类的繁衍聚集,所以生物多样性高。在另一项不同年份不同季节(2015年冬季)的同类研究[37]中,设立10个站点对东海、黄海的鱼类进行多样性调查,其中Shannon多样性指数范围为1.97~4.22,Pielou均匀度指数范围为0.35~ 0.70,均高于本研究的对应数据,即多样性和均匀度更高,主要原因可能是研究区域的差异,该研究中10个站点分别位于黄海海域和东海海域,各自较为集中,而本研究14个站点仅位于东海海域,且跨度大呈直线分布;而季节性以及年际差异也可能造成一定影响,冬季的东海为一些重要经济鱼类的越冬场所,而本研究采样季节主要是鱼类的产卵及索饵育肥季节,且产卵场和索饵场主要分布在大陆架和近海海区[38],在一项2001年的底拖网调查[39]中也发现,12月份东海中部鱼类群落多样性指数高于9月,伏季休渔结束后对主要经济鱼类物种的捕捞使得东海鱼类群落多样性和均匀度更高。
本研究首次利用eDNA技术分析了东海海域夏季鱼类物种多样性,且采样站点跨度大,研究海域范围广,站点间多样性差异较大,获得了丰富的鱼类物种信息,鉴定所得44种海水鱼类有8种属于鲱形目,7种属于鲭形目。eDNA技术的便捷、经济、高效以及无损性使其在东海这类典型海域的生物多样性研究中有良好的应用前景。不过本研究在采样上仍有不足,首先采样站点在一条东北-西南走向的直线上,缺乏同一经纬度的数据对比;其次采样深度仅为表层,而采样时间随机,这可能会使分析结果缺乏生活水层较深和昼夜活跃度不同的鱼类物种数据。因此,今后在基于eDNA的东海鱼类物种多样性调查中应增加同一经纬度的站点设计,同一站点应尽量分别采集昼夜样本,合理设计采样深度,并进行长期监测。
猜你喜欢
当代水产(2022年7期)2022-09-20
儿童时代·幸福宝宝(2020年9期)2020-09-08
中华诗词(2019年2期)2019-08-27
探索科学(2017年5期)2017-06-21
法制与社会(2016年34期)2017-01-14
河北渔业(2016年12期)2017-01-11
河北渔业(2016年11期)2016-12-16
诗潮(2014年7期)2014-02-28
