
横道河子地区大林姬鼠肠道菌群组成的性别差异
2022-04-14 02:13周思宇杨文建于成文姜明明金志民
湖北畜牧兽医 2022年2期
苏 航,周思宇,杨文建,于成文,姜明明,金志民
(1.牡丹江师范学院生命科学与技术学院,黑龙江 牡丹江 157011;2.黑龙江农业经济职业学院,黑龙江 牡丹江 157041)
大林姬鼠为哺乳纲啮齿目鼠科姬鼠属动物,体型细长,多栖息于林区、灌木丛、林区空地和林区边缘的农田之中,多以阔叶树的种子和果实为食[1]。在中国主要分布于西南、西北、华北和东北等地区,是中国农林地区较为常见的小型啮齿类哺乳动物[2]。关于大林姬鼠生态与生理生化方面的研究较多[3-6],但对大林姬鼠肠道微生物的研究尚不充足。肠道微生物对宿主的营养消化吸收、免疫、代谢以及维持宿主肠道内环境稳定起着重要的作用[7]。相关研究证明,肠道菌群是肠道微生物中占比最大的组成部分。影响肠道菌群结构及多样性的因素较多,如食性、年龄、季节、饲养方式及海拔等[8]。性别因素对动物的生理和行为有明显影响,但关于性别因素对肠道菌群组成的影响难以形成明确的结论。本研究采用16S rDNA 高通量测序技术,对采自横道河子地区的大林姬鼠进行研究,分析其肠道菌群的组成及性别因素对组成的影响。
1 材料与方法
1.1 样品的采集与保存
样品取自黑龙江省牡丹江市横道河子国家自然保护区(海拔460~600 m,44°44′—44°55′N,129°06′—129°15′E)。采用笼捕法,共采集 13 只样本,选取体型相似、解剖后无明显患病症状的健康成年鼠雌雄各5 只,其中雌鼠均未怀孕。对采集的样本进行编号,如表1 所示。在实验室无菌条件下,将鼠脱颈处死,并进行解剖,取出鼠的盲肠立即放入装有液氮的研钵中速冻,装入无菌冻存管,标记好放入-80 ℃冰箱。

表1 大林姬鼠个体信息
1.2 肠道微生物DNA 的提取
采用CTAB/SDS 方法,对肠道组织进行总DNA的提取,经过1.8%琼脂糖凝胶电泳20 min 检测有明亮条带为提取成功。在每个样本的通用引物两端加上不同的 barcode。 使用通用引物 515F-806R(515F:5′GTGCCAGCMGCCGCGGTAA-3′,806R:5′-GGACTACHVGGGTWTCTAAT-3′)对 16S rDNA 基因V3-V4 区进行扩增。纯化产物送至北京百迈克生物科技有限公司用HiSeq2500 PE250 平台进行双端测序。
1.3 数据处理
获得所有样本的测序数据后,去除引物接头序列,使用 FLASH v1.2.11 软件,通过overlap 对每个样品的reads 进行拼接,得到的拼接序列即原始Tags数据(Raw Tags),使用Trimmomatic v0.33 软件,对拼接得到的Raw Tags 进行过滤,得到高质量的Tags数据,使用UCHIME v4.2 软件,鉴定并去除嵌合体序列,得到各样本的有效数据。
2 结果与分析
2.1 样品中有效序列和OTUs
通过高通量测序技术对10 份大林姬鼠的盲肠样本进行16S rDNA 测序。共获得有效序列633 165条,平均每个样品中含有63 316 条有效序列,其中雌性含有316 011 条,雄性含有317 154 条。有效序列数量最高的是DL8,有76 856 条有效序列,最低的是DL6,有54 513 条有效序列,二者相差22 343条。基于相似度>97%的原则将获得的有效Tags 序列进行聚类,共获得899 个OTUs。韦恩图中重叠部分为雌性样品与雄性样品共有的OUTs,由图1 可知雌雄两组中共有738 个OTUs。
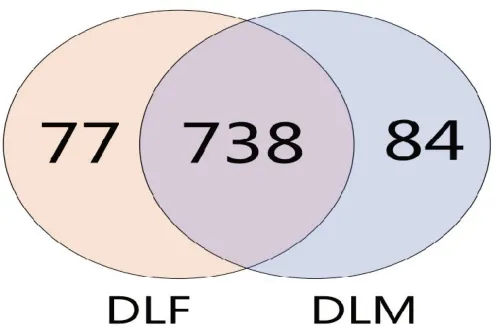
图1 大林姬鼠雌性和雄性OTUs
2.2 Alpha 多样性分析
通过对10 份样品的Alpha 多样性分析指数进行丰富度和多样性比较。从图2 可以看出,随着测试深度的不断加深,稀释曲线从不断上升,直至趋于平缓,说明样品的测序深度比较合理。使用SPSS19.0软件对大林姬鼠组和黑线姬鼠组的Alpha 多样性指数进行差异性分析,结果如表2 所示,两组的Ace、Chao1、Shannon、Simpon 4 个指数均差异不显著(P>0.05),说明不同性别的大林姬鼠肠道菌群丰富度无显著差异。
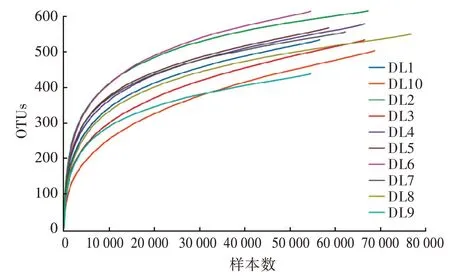
图2 雌性和雄性样品稀释曲线
2.3 物种分类分析
2.3.1 门水平菌群组成差异 由图3 可知,在门水平上,雌性和雄性大林姬鼠含量最高的5 个菌门是相同的,分别为厚壁菌门( Firmicutes) 、拟杆菌门(Bacteroidetes)、变形菌门(Proteobacteria)、Epsilonbacteraeota、脱铁杆菌门(Deferribacteres)。其中,各菌门的含量分别为厚壁菌门(雌52.70%,雄44.94%)、拟杆菌门(雌30.24%,雄27.47%)、变形菌门(雌 7.07%,雄 17.51%)、Epsilonbacteraeota(雌7.01%,雄6.98%)、脱铁杆菌门(雌2.23%,雄1.35%)

图3 大林姬鼠门水平下雌性和雄性菌群相对丰度
2.3.2 属水平菌群组成差异 由图4 可知,在属水平上,雌、雄大林姬鼠肠道内各菌群存在比例略有差异,雌性大林姬鼠肠道内占比最高的5 个优势菌属依次为 Unidentified-Muribaculaceae(25.42%)、Unidentified-Lachnospiraceae(20.79%)、螺杆菌属(Helicobacter)(7.01%)、乳杆菌属(Lactobacillus)(6.55%)、螺 菌 科 NK4A136 组Lachnospiraceae-NK4A136-group(4.42%)。雄性大林姬鼠肠道内占比最高的5个优势菌属依次为Unidentified-Muribaculaceae(18.31%)、Unidentified-Lachnospiraceae(15.94%)、假单胞菌属(Pseudomonas)(10.57%)、螺杆菌属(Helicobacter)(6.98%)、螺菌科 NK4A136 组Lachnospiraceae-NK4A136-group(6.89%)。
2.4 Anosim 分析
Anosim 分析是非参数检验,基于Bray-Curtis 距离值的秩次进行每两组间差异显著性检验,用来判断组间差异是否大于组内差异。R-value 在(-1,1)之间,R-value >0 说明组间差异大于组内差异;R-value < 0 说明组内差异大于组间差异;P>0.05 说明组间差异不显著,0.01<P<0.05 说明组间差异显著,P<0.01 说明组间差异极显著。大林姬鼠雌雄两组间R=0.068,说明组间差异大于组内差异,P=0.338(P>0.05),说明大林姬鼠雌雄两组肠道菌群差异不显著。
3 小结与讨论
通过16S rDNA 高通量测序技术对横道河子地区大林姬鼠肠道菌群的性别差异进行了分析。结果表明,不同性别的大林姬鼠肠道菌群组成没有显著差异。关于性别对鼠类肠道菌群组成的影响研究结果并不一致。杨静等[9]对甘肃鼢鼠肠道菌群进行研究后发现,肠道菌群组成和结构在雌雄间差异不显著。Org 等[10]对具有特定遗传背景的小鼠进行检测时发现性别差异对肠道菌群的组成有明显影响。也有学者认为性别差异对肠道菌群结构的影响不显著,原因可能是性别差异被其他因素,如宿主遗传和环境因素所掩盖[11]。本研究的样品虽然是在同一地点,同一时期采集的,但不能控制其遗传背景。因此可以推测本研究中大林姬鼠的肠道菌群雌雄差异并不显著,可能是由于无法准确的控制变量因素。动物肠道菌群受性别影响的机理还尚未明确。Yurkovetskiy 等[12]对小鼠研究表明,小鼠肠道菌群组成的性别差异直到青春期后才出现,且在雄性小鼠阉割后这一差异消失。所以性别因素造成肠道菌群的差异有可能是由于性激素对机体作用所造成的影响。
总的来说,性别可能是影响动物肠道菌群的因素,但由于野生大林姬鼠的其他变量无法控制,导致大林姬鼠雌雄个体肠道菌群的差异不显著。性激素水平也可能是影响微生物组成的部分因素,但机制尚未明确,需要进一步分析。
猜你喜欢
中老年保健(2022年2期)2022-08-24
小学生优秀作文(低年级)(2021年10期)2021-11-24
湖北农业科学(2021年2期)2021-02-07
科学(2020年4期)2020-11-26
湖北农业科学(2019年20期)2019-12-11
贵州农业科学(2019年7期)2019-08-03
湖北农业科学(2019年23期)2019-01-11
作文大王·低年级(2017年1期)2017-02-16
动物营养学报(2015年10期)2015-12-01
爆笑show(2015年3期)2015-05-08
