
丛生盔形珊瑚光合生理及共生真菌群落分析
2022-03-31 01:02朱文涛夏景全刘相波尹洪洋朱铭任瑜潇谢敏睿黄建中李秀保
热带海洋学报 2022年2期
朱文涛, 夏景全, 刘相波, 尹洪洋, 朱铭, 任瑜潇, 谢敏睿,黄建中, 李秀保
1. 海南大学生态与环境学院, 海南 海口570228;
2. 海南大学海洋学院, 海南 海口570228;
3. 海南大学南海海洋资源利用国家重点实验室, 海南 海口 570228
珊瑚礁生态系统大多数位于寡营养盐的热带海洋环境中, 却是地球上最多样化和生产力最高的生态系统之一。其中造礁石珊瑚的礁石建造能力和形成的复杂物理结构能为珊瑚礁中丰富的生物提供栖息地, 并支持广泛的生态系统服务功能和为人类提供重要的经济服务价值(Roberts et al, 2002)。珊瑚礁生态系统覆盖不到海洋表面的1%面积, 但它们却在维护海洋生物多样性、缓解全球气候变化中起着十分重要的作用, 是对人类及其他生物不可或缺的海洋资源(Spalding et al, 2015)。造礁石珊瑚是珊瑚礁生态系统生物多样性的基础和关键物种, 对气候变化和人类活动的干扰非常敏感(Putnam et al, 2017)。在进入人类世时代后, 大规模白化事件的发生频率和严重程度逐渐增加(Hughes et al, 2018a), 世界上各区域的珊瑚礁正在经历广泛的白化事件, 这对当前珊瑚礁构成了最大的威胁, 因此人类世的热带珊瑚礁的覆盖面积正在迅速减少(Hughes et al, 2017)。如果白化事件持续时间较长, 还会改变珊瑚礁群落组成和结构的复杂性(Hussain et al, 2020), 继而导致生物多样性下降和生态系统服务功能衰退, 对生态、经济和社会产生长期和广泛的影响(Hughes et al,2018b)。
珊瑚宿主与共生藻类、细菌、病毒和真菌等多种微生物构成了珊瑚共生功能体(Thompson et al,2015), 其中珊瑚动物宿主与其共生虫黄藻之间存在互惠互利的紧密关系, 使得珊瑚能对养分的高效吸收和营养物质的再循环利用(Rädecker et al, 2015)。虫黄藻通过光合作用固定的绝大多数碳和氮化合物能被珊瑚宿主获取, 这可以满足动物宿主高达95%的能量需求和日常代谢(Yellowlees et al, 2008), 而珊瑚宿主则为共生藻提供了包括二氧化碳和铵盐等营养物质(Wooldridge, 2010)。超过珊瑚耐受范围的热胁迫是造成珊瑚白化的主要因素, 而且由于人类活动引起的压力也会引起珊瑚褪色(Quimpo et al,2020)。先前的研究表明, 白化会严重破坏珊瑚的生理特征, 若珊瑚宿主与虫黄藻之间的共生关系崩溃,珊瑚还会发生不可逆转的死亡现象(Baker, 2003)。进一步研究表明, 珊瑚对热漂白的敏感性和抵抗力与虫黄藻密度有关(Wooldridge, 2014), 共生的虫黄藻密度是评估珊瑚耐热性和健康状况的关键指标(Qin et al, 2019)。虫黄藻的光合生理变化会影响珊瑚的健康发育(黄玲英 等, 2011), 因此珊瑚共生藻光合生理研究可以揭示不同胁迫下珊瑚光合特性和白化机制(周洁 等, 2011)。同时, 珊瑚白化的敏感性以及共生关系的功能取决于共生虫黄藻密度和光合能力,所以开展不同珊瑚种类和不同野外条件下的珊瑚光合生理研究非常重要(Xu et al, 2017a)。
珊瑚微生物组在生物地球化学循环、物质转化和维护珊瑚健康等方面发挥着重要作用(Biagi et al,2020), 它们通过有效地捕获、保留和循环养分以帮助珊瑚礁在寡营养的热带水域中生存和生长(Vanwonterghem et al, 2020)。目前, 珊瑚相关微生物的研究主要集中栖息在珊瑚宿主组织中的共生虫黄藻和细菌群落, 而对其他真核微生物(例如真菌、原生生物)的物种多样性和功能贡献的了解有限(Ainsworth et al, 2017), 特别是作为珊瑚微生物组最重要组成部分之一的真菌在生物地球化学循环、物质转化和维护珊瑚健康等方面发挥着重要作用。人们对珊瑚宿主与共生细菌之间的相互作用了解得相对较多(Amend et al, 2012), 但对共生真菌的研究相对较少, 特别是对珊瑚共生真菌的物种多样性及其在维持珊瑚的健康中发挥的生态功能知之更少(Góes-Neto et al, 2020)。
丛生盔形珊瑚Galaxea fascicularis隶属于真叶珊瑚科(Euphylliidae)盔形珊瑚属(Galaxea), 广泛分布在红海、印度洋—太平洋的大部分海域, 也是我国近岸珊瑚礁以及南海海域包括海南岛的重要生态和优势种之一。目前, 对丛生盔形珊瑚的分子相关研究主要包括基于核糖体DNA 第二内部转录间隔区(ITS2)序列来探索共生藻科的多样性组成(Zhou et al, 2017)、在不同胁迫下(例如富营养化、热胁迫漂白时、pH 极端变化)的转录组反应等(Lin et al, 2017,2018; Hou et al, 2018)。另外, 还有通过传统的稀释分离法、培养基等方法来研究丛生盔形珊瑚的共附生真菌, 这些文献主要关注丛生盔形珊瑚共附生可培养真菌多样性(徐佳 等, 2011a)、分离及其抗菌活性的筛选(徐佳 等, 2011b)、真菌的鉴定及生理学特性(王荻潇 等, 2018)等方面, 但基于高通量测序方法来探究健康和白化丛生盔形珊瑚的真菌组成和多样性未见报道。在2020 年9 月热胁迫期间, 本研究在调查海南岛西北部近岸海域理化环境要素的基础上, 比较分析了优势丛生盔形珊瑚在健康和白化状态下的虫黄藻密度、光合生理指标和共生真菌群落的差异。
1 材料与方法
1.1 研究区域
海南岛西北部海域珊瑚礁是南海珊瑚礁典型的岸礁, 属于印度洋—太平洋区系珊瑚礁的重要组成部分。近年来随着海南岛的建设和发展, 其西北部海域被过度开发利用, 造礁石珊瑚群落受到破坏退化严重, 生态系统多样性和覆盖率急剧下降(黄晖等, 2012)。2020 年8 月, 海南岛西北部海域水温持续高温, 出现了罕见的大规模珊瑚白化现象, 而且这些珊瑚中的多数种类为高温耐受性物种。本研究是2020 年9 月在海南岛西北部的美夏(Meixia, MX)、黄龙(Huanglong, HL)、排浦(Paipu, PP) 3 个海域进行的。使用GPS 记录调查采集样品站位的经纬度。使用浊度仪(Aqualogger210)、便携式 pH/电导率仪(420C-01A Orion Star, Thermo Fisher Scientific, 美国)原位分别记录了海水的温度、盐度和pH。使用采水器收集底层接近珊瑚礁基质附近深度为1m 的海水1L, 在经过孔径0.45μm 的混合纤维素酯微孔滤膜过滤后, 收集100mL 滤液到试剂瓶中, 使用营养盐全自动分析仪(德国SEAL, AA3)检测水样的无机氮、无机硅酸盐、磷酸盐等溶解无机营养盐含量。所有采样站位的海水温度均超过30℃, 超过了珊瑚生存最适范围的上限(表1)。美夏区域的海水温度均值最高, 介于31.75~34.07℃之间。3 个研究区域的pH 和盐度相差不大, 都处于珊瑚正常生长的范围内。根据海水水质标准(GB3097-1997), MX 和HL 海域的溶解态无机氮(DIN)、磷酸盐(DIP)平均浓度在第二类海水和第三类海水标准范围之间, PP 海域的DIN、DIP 属于第一类海水。

表1 调查站位的地理坐标及水体环境参数Tab. 1 Geographical locations of the sampling stations
1.2 珊瑚采样
选择耐热珊瑚物种丛生盔形珊瑚为研究目标,分别在3 个研究区域采集健康珊瑚和白化珊瑚, 其中, 健康组珊瑚是指从视觉上确保珊瑚的健康状况良好的珊瑚。在每个地点分别选择3 个重复采样点,利用锤子和凿子收集健康和白化的珊瑚样品后, 将其暂养在装有海水的泡沫箱中, 以待测量光合生理指标。回到实验室立即用过滤的海水冲洗部分珊瑚碎片表面, 并将每个珊瑚分成实验所需大小的样品,暂放在–20℃冰箱中, 其中微生物样品装入灭菌的离心管中保存在–80℃超低温冰箱以待分子测序。
1.3 光合生理参数和虫黄藻密度
将采集的珊瑚暂养并黑暗适应半个小时后, 使用叶绿素调制荧光仪MINI-PAM-Ⅱ测量珊瑚共生虫黄藻的光合生理参数最大光化学量子产量(Fv/Fm)。在测量过程中PAM 的光纤探头与珊瑚表面的固定距离保持在1cm 左右, 随机选择每个珊瑚样品的3~5 个不同位置进行重复测定, 并计算平均值。使用洁碧洗牙器和0.45μm 滤膜过滤的海水冲洗珊瑚表面, 量取10mL冲洗后的溶液, 在显微镜下通过血细胞计数板计数来测定虫黄藻密度。根据不同已知表面积大小的铝箔纸进行称重, 建立面积和重量线性关系式; 然后用铝箔纸包裹珊瑚表面后称重, 根据上述关系式, 通过重量来间接估算珊瑚表面积。最终结果使用单位珊瑚表面积的虫黄藻密度(单位: 个∙cm–2)表示。
1.4 高通量测序
将保存好的珊瑚样品在经过研钵磨碎成匀浆溶液后转移到试管中, 按照海洋动物基因组DNA 提取试剂盒制造商的步骤提取珊瑚共生体基因组DNA。使用 NanoDrop ND-1000 型分光光度计测定(NanoDrop Technologies, Wilmington, USA) DNA 的浓度和纯度, 在1%琼脂糖凝胶电泳验证DNA 的质量。使用带有条形码的通用引物ITS3 (5′-GCATCGA TGAAGAACGCAGC-3′)和 ITS4 (5′-TCCTCCGCTT ATTGATATGC-3′)进行扩增。PCR 反应总体积为30μL,扩增的条件为在98℃下预变性1min, 然后依次进行30 个循环: 98℃变性10s, 50℃退火30s, 72℃延伸60s,最后72℃ 5min。利用2%琼脂糖凝胶电泳检测PCR产品量化和鉴定后, 将PCR 产物以等比例混合纯化后, 根据NEB Next®Ultra™ DNA Library Prep Kit for Illumina (NEB, USA)制造商的步骤生成测序文库。在经过 Qubit@2.0 Fluorometer (Thermo Scientific)和Agilent Bioanalyzer 2100 system 评估library 质量后,在Illumina MiSeq 平台进行测序。所得的测序数据经去除冗余和无效序列后, 选取操作分类单元(以97%的序列相似性聚类)的代表性序列, 使用 RDP(ribosomal database project)分类器确定可操作分类单元(operational taxonomic units, OTUs)的注释分类信息(Wang et al, 2007)。按照最低测序深度26542 抽平处理后用于后续分析。不同样品真菌的功能预测信息是通过FUNGuild 数据库(http://www. funguild.org/)得到(Nguyen et al, 2016)。
1.5 统计分析
在差异分析之前, 对数据进行正态性假设检验。使用t检验分析健康珊瑚和白化珊瑚的差异, 如果数据不符合正态分布, 考虑非参数的检验方法Wilcox 秩和检验分析。结果表示为平均值±标准误差,p<0.05 认为是差异显著。在OTUs 水平上计算了真菌的Shannon 指数、覆盖率指数, 并使用方差分析或者非参数检验比较多样性指数在不同分组的差异。基于Bray-Curtis 相异矩阵的非度量多维尺度分析(nonmetric multidimensional scaling, NMDS)来分析样本间的真菌群落构成差异, 并使用Permutational ANOVA and MANOVA (PERMA NOVA)来检验各组的群落差异是否显著。使用pheatma 包绘制真菌预测功能的丰度热图, 以上的数据分析和图形的可视化均于R 软件(R4.0.3 版本)中完成的。最后使用在线工具(http://huttenhower.sph.harvard.edu/galaxy/root?tool_id=lefse_upload)进行线性判别差异分析(Linear discriminant analysis Effect Size, LEfSe), 以期找出对各组产生显著性差异影响的指示物种。
2 结果
2.1 虫黄藻与光合生理
如图1a 所示, 健康组丛生盔形珊瑚的共生虫黄藻平均密度为(1.85±0.09)×106个∙cm–2, 介于(1.61~2.32)×106个∙cm–2之间; 白化组珊瑚的共生虫黄藻平均密度为(0.87±0.11)×106个∙cm–2, 介于(0.11~0.87)×106个∙cm–2之间。如图1b 可知, 健康组珊瑚的Fv/Fm均值为0.68, 介于0.63~0.69 之间。白化组珊瑚的Fv/Fm维持在较高水平, 均值为0.57,介于0.41~0.68 之间。使用t检验分析表明珊瑚健康状态和白化状态下虫黄藻密度与光合生理参数存在显著性差异(p<0.05), 白化珊瑚的最大光化学量子产量Fv/Fm、虫黄藻密度比健康珊瑚中相应的指标参数显著降低了16%和75%。

图1 健康组和白化组丛生盔形珊瑚中虫黄藻密度(a)和最大光化学量子产量(Fv/Fm, b)分析图中*表示两组之间具有显著性差异(p<0.05); 图b 中虚线表示0.5 的阈值Fig. 1 Analysis of zooxanthellae density (a) and maximum photochemical quantum yield (Fv/Fm, b) between healthy and bleaching Galaxea fascicularis
2.2 群落组成
通过高通量测序方法对18 个丛生盔形珊瑚样品的共生真菌群落组成进行分子测序, 经过一系列过滤筛选过程后, 留下与真菌组相关的序列进行进一步分析; 在对最小读数进行二次抽样后,按照 97%的相似阈值对有效序列进行聚类, 并与数据库UNITE (https://unite.ut.ee)比对进行物种注释。结果表明, 在不同的分类水平上, 共检测到4门、25 纲、63 目、157 科、302 属、472 种。在白化组中的优势菌门为子囊菌门(Ascomycota)、担子菌门(Basidiomycota)、壶菌门(Chytridiomycota)、毛霉门(Mucoromycota), 相对丰度分别为80.43%、16.78%、2.31%、0.43%; 在健康组中优势菌门为Ascomycota、Basidiomycota、Chytridiomycota、Mucoromycota, 相对丰度分别为86.09%、11.93%、1.74%、0.23%。在门水平上(图 2a), 健康组和白化组中均无现显著性差异(p>0.05)。在属水平上(图2b), 真菌前10 个优势属的相对丰度从高到低,分别为Gloeotinia、曲霉属(Aspergillus)、枝孢菌属(Cladosporium)、 丝孢子菌属(Sporothrix)、Kazachstania、 链 格 孢 霉 属 (Alternaria) 、Solicoccozyma、侧 齿 霉 属(Engyodontium)、晶杯菌属(Hyaloscypha) 、Juxtiphoma。 其中,Gloeotinia、Aspergillus两个属在健康组和白化组中的平均相对丰度均在 10%以上, 分别为23.94%、21.04%和10.48%、13.15%。但是这些优势属在白化组和健康组中的平均相对丰度均无显著差异(p>0.05), 除了Solicoccozyma在白化组(5.66%)中显著高于健康组(2.42%)。
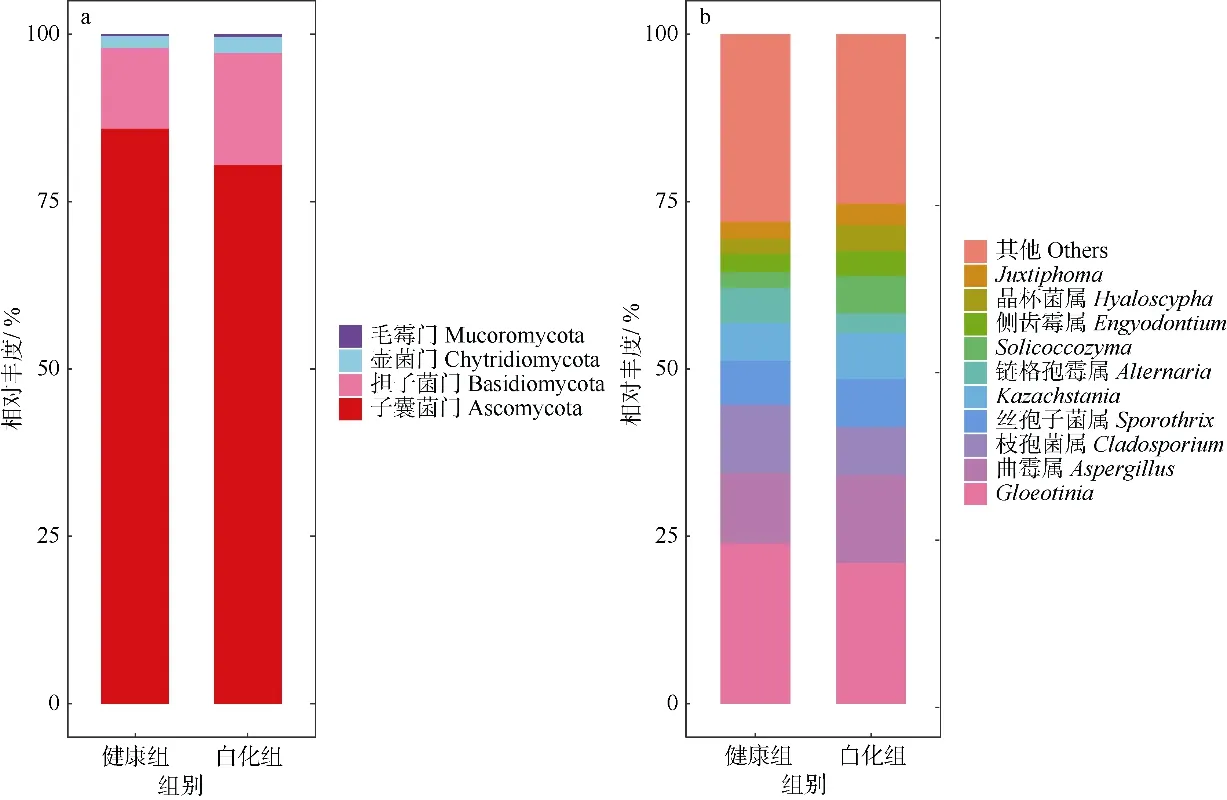
图2 健康组和白化组丛生盔形珊瑚在门水平(a)和属水平(b)的真菌相对丰度图Fig. 2 Relative abundance of fungi in Galaxea fascicularis at phylum (a) and genus (b) level
2.3 真菌多样性
根据在每个单独的样本中显示观察到的OTUs的数量和相对于总识别的序列的数量来构建稀释曲线(图3a), 除了样品P5 有更低程度的稀薄形状外,本次测序的绝大部分样品的稀释曲线在测序深度为10000 左右时趋于平缓状态, 另外16 个测序样品的覆盖度指数均到达了0.999, 这些表明共生真菌的测序深度足够进行可靠的α多样性分析以及保证了测序结果的合理性。健康组和白化组丛生盔形珊瑚中共生真菌Shannon 多样性指数如图3b 所示。其中健康组的Shannon 多样性指数平均值为3.78±0.24,变化范围分别为2.28~4.66 之间; 白化组中共生真菌的Shannon 多样性指数平均值为3.94±0.24, 变化范围介于2.92~5.24 之间。尽管共生真菌的Shannon多样性指数平均值在健康组中低于白化组, 但两者之间并无显著性差异。
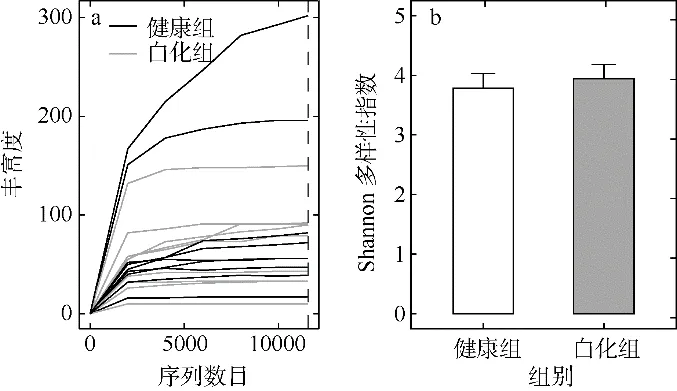
图3 健康组和白化组丛生盔形珊瑚共生真菌稀释曲线(a)和Shannon 多样性指数(b)图a 中每一条曲线代表一个样本, 竖直虚线表示样本测序深度情况Fig. 3 Dilution curve (a) and Shannon diversity index (b)of symbiotic fungi in healthy and bleaching Galaxea fascicularis
2.4 差异群落
基于Bray-Curtis 距离矩阵进一步利用非度量多维尺度分析(nonmetric multidimensional scaling,NMDS)分析了样本群落即β-多样性的差异性(图4a),当应力值小于0.2 时, 表示分析结果具有一定的可靠性。本研究中的应力值为0.066, 说明数据可用于NMDS 分析。如图4 所示, 无论是健康组还是白化组中, 大部分样本之间的分布距离较远且较为分散,这说明珊瑚真菌群落结构的组内差异性大于组间差异性, 珊瑚的真菌群落存在很强的个体异质性。使用PERMANOVA 方法进行检验分析发现, 健康组和白化组的丛生盔形珊瑚中共生真菌群落结构较为稳定,β-多样性并不存在显著性差异(p>0.05)。通过线性判别分析(LEfSe)发现(图4b), 健康组和白化组之间在丰度上存在显著差异的真菌物种类群仅有7 类。其中, 在健康组中富集的类群有4 个, 包括微球黑粉菌纲(Microbotryomycetes)、锁掷孢酵母科(Sporidiobolaceae)、锁掷酵母目(Sporidiobolales)、红酵母属(Rhodotorula)。在白化组丰度较为丰富的仅有线黑粉菌目(Filobasidiales)、Piskurozymaceae科、产油菌属(Solicoccozyma)共3 个类群。
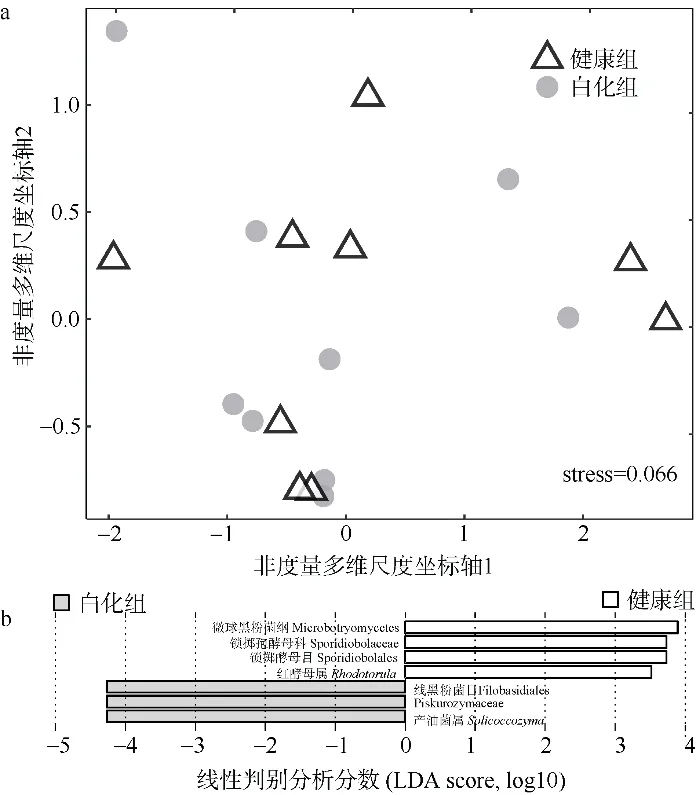
图4 健康组和白化组丛生盔形珊瑚共生真菌群落组成的非度量多维尺度(a)和线性判别分析(b)Fig. 4 NMDS analysis (a) and LEfSe analysis (b) of Fungi community composition in healthyand bleaching Galaxea fascicularis
2.5 预测功能
基于真菌的物种分类数据, 利用FUNGuild 数据库预测了丛生盔形珊瑚共生真菌群落的营养型。如图5 所示, 丛生盔形珊瑚真菌有280 个OTUs, 可被划分为7 个功能类群, 包括3 类营养型和4 类互有交叉营养型功能菌群, 分别是共生菌群(Symbiotroph, 3.53%) 、 腐生菌群(Saprotroph,11.10%)、病理寄生菌群(Pathotroph, 19.55%)、腐生-共生过渡型(Saprotroph-Symbiotroph, 0.38%)、病理寄生-共生菌群(Pathotroph-Symbiotroph, 9.24%)、病原菌- 腐生- 共生菌群(Pathotroph-Saprotroph-Symbiotroph, 11.67%) 、 病理寄生- 腐生菌群(Pathotroph-Saprotroph, 5.70%), 剩余的是未定义的营养型(38.83%)。但是这些真菌功能类群在健康组和白化组中的相对丰度均无显著差异(p>0.05)。
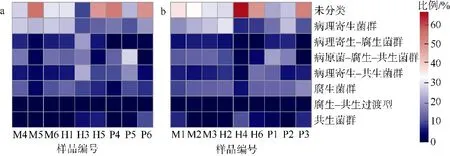
图5 健康组(a)和白化组(b)丛生盔形珊瑚共生真菌的FunGuild 功能预测相对丰度热图Fig. 5 Function predict relative abundance heat map based on FunGuild
3 讨论
珊瑚因环境胁迫而引起共生藻丧失、光合色素减少进而褪色的现象被称为白化。人们普遍认为块状珊瑚比分枝状珊瑚具有更高的热应力承受能力(Wooldridge, 2014)。研究发现, 含高密度共生虫黄藻的块状珊瑚遭受热白化胁迫后比低密度的物种存活率更高(Qin et al, 2019)。这可能与高密度的虫黄藻会导致更有效的自我遮蔽能保护珊瑚免受高强度光的伤害有关(Xu et al, 2020)。本研究中健康丛生盔形珊瑚的共生虫黄藻密度为(1.85±0.09)×106个∙cm–2,该数值低于南海北部三亚湾和大亚湾(李淑 等,2007)、南海黄岩岛(潘子良 等, 2019) 等海域的块状珊瑚共生虫黄藻密度。从调查的海水温度的结果可以看出, 本次采样期间调查的3 个区域所有站位的海水温度都超过30℃, 并且美国国家海洋和大气管理局(National Oceanic and Atmospheric Administration,NOAA)卫星数据显示调查区域海水温度一直处于较高水平。可以推测, 珊瑚虫黄藻可能受到热应激的威胁而数量显著减少, 但健康珊瑚虫黄藻含量仍在建议的最佳共生体密度 1~3×106个∙cm–2范围之内(Wooldridge, 2020)。在本研究中白化珊瑚的虫黄藻密度比健康珊瑚的显著降低了75%。虫黄藻密度是健康珊瑚和白化珊瑚外观差异最直接和最直观的证据之一, 丛生盔形珊瑚呈现出白化的极端外观表征, 这是由于共生虫黄藻被逐渐排出后密度显著降低到一定阈值的表现(李淑 等, 2011)。
光系统Ⅱ(PSⅡ)最大量子产量(Fv/Fm), 能反映和指示造礁石珊瑚共生虫黄藻的潜在光合能力和生理状态(Gorbunov et al, 2001)。在本研究中白化珊瑚的Fv/Fm比健康珊瑚中的降低了16%, 说明白化状态下虫黄藻的光合作用原初反应过程受到抑制、光合效率降低。Fv/Fm低于0.5 通常被用来指示虫黄藻光系统Ⅱ遭到损害(Bhagooli et al, 2004)。尽管这项研究中白化珊瑚观察到光合产量的显著降低, 但在白化珊瑚Fv/Fm的平均值仍然大于0.5, 表明光合电子传递系统并没有受到破坏, 暗示了丛生盔形珊瑚在高温下光合生理的适应能力较强。
本研究利用高通量测序技术评估了珊瑚中的真菌多样性和组成, 在白化和健康丛生盔形珊瑚样品中鉴定出4 个优势真菌门, 分别为子囊菌门Ascomycota、担子菌门Basidiomycota、壶菌门Chytridiomycota、毛霉门Mucoromycota。这与西南太平洋不同珊瑚属骨骼和珊瑚藻相关的真菌中鉴定的结果一致(Góes-Neto et al,2020)。在菌属分类水平上,Gloeotinia属的平均相对丰度均在20%以上, 这与海口海甸岛荣域温泉(舒为 等,2020)、南大西洋中脊的深海热液喷口站点(Xu et al,2017b)的真菌序列中的报道一致。其中该真菌属的Gloeotinia temulenta是一种内生真菌, 是可在牧草种子中生长的病原体(Alderman, 1998), 但是在本研究中仅仅鉴定到属, 该属在丛生盔形珊瑚中的具体功能仍然不确定。
相对丰度较高的曲霉属Aspergillus、枝孢菌属Cladosporium、链格孢霉属Alternaria等主要为陆生的子囊菌属物种(Góes-Neto et al, 2020), 在丛生盔形珊瑚(徐佳 等, 2011a)和南海3 种珊瑚(张丹 等,2015)中均有发现。这些共附生真菌是重要生物活性物质的生产者之一, 也是珊瑚微生物组的常见成员。杂色曲霉(Aspergillus versicolor)是曲霉属中研究最广泛的珊瑚真菌之一,据报道是加勒比海柳珊瑚(Harvell et al, 2007)和佛罗里达礁群岛柳珊瑚Gorgoniaspp.中的致病病原体(Alker et al, 2001)。尽管有研究在健康的珊瑚和周围海水中也发现了Aspergillus versicolor(Soler-Hurtado et al, 2016), 但在本文中健康组和白化组丛生盔形珊瑚中均未出现该物种。毛霉菌门的物种可能会引起海洋动物疾病,不过在本研究中鉴定到的丰度极低, 在健康和白化珊瑚中均低于0.5%。
其他优势属例如Gloeotinia、丝孢子菌属Sporothrix、Kazachstania、Solicoccozyma、侧齿霉属Engyodontium、 晶杯菌属Hyaloscypha、Juxtiphoma等未在以往丛生盔形珊瑚可培养真菌的报道中发现。这可能与传统的微生物分离培养方法的限制有关, 大约95%的菌落不能被成功分离培养出来。例如有学者(徐佳 等, 2011a)利用5 种培养基在丛生盔形珊瑚中共分离出19 株共附生真菌, 这些真菌基本在本研究的高通量测序结果中被检测到较低的丰度, 这可能与受到PCR 引物偏好性的影响有关(Deiner et al, 2017)。本研究中的所有样品都采集于高温胁迫期间, 这可能对珊瑚共附生真菌种类和丰度产生影响。
基于真菌的物种分类数据和 FUNGuild 数据库, 丛生盔形珊瑚共生真菌群落预测结果共发现3 类营养型和4 类互有交叉营养型功能菌群。但是利用 FUNGuild 数据库得到真菌的功能预测信息是不全面的(Nguyen et al, 2016), 其中本研究中未定义的营养型可达38.826%。在本研究中真菌类群划分到病理寄生菌群和病原菌-腐生-共生菌群的相对丰度分别为19.551%和11.670%。这些预测功能仅是基于真菌群体的摄食习惯, 它们的功能并非完全是致病性的, 也有可能会帮助珊瑚宿主形成稳定的共生关系(Góes-Neto et al, 2020)。本研究结果侧重于真菌多样性检测和功能预测, 然而这些预测有待通过真菌培养和宏基因组技术进一步详细验证, 因此在后续研究中可以结合这些方法来更全面评估和阐明丛生盔形珊瑚共附生真菌的实际生态功能。
研究表明珊瑚在热应激、藻类胁迫等各种压力下, 其共生细菌的α-多样性和β-多样性会响应压力而发生改变(Zaneveld et al, 2016), 这表明珊瑚宿主调节共生微生物组之间相互作用的平衡稳定以及抵抗病原体感染并维持健康微生物组功能的能力降低(Pollock et al, 2019)。当微生物组群落结构改变和功能失衡后(Patel et al, 2020), 珊瑚最终可能由于遭受机会性微生物的侵害而白化和死亡(Glasl et al, 2017)。本研究结果表明, 健康组丛生盔形珊瑚共生真菌的 Shannon 指数的平均值(3.78±0.24)低于白化组的(3.94±0.24), 但差异性不显著。在丰度上有显著差异的真菌物种类群有7 类,平均丰度所占比例较小, 因此丛生盔形珊瑚在健康和白化状态下共生真菌群落组成相对稳定。珊瑚病原体和机会真菌类群没有在白化珊瑚中显著增加, 并且它们也可以在健康珊瑚样本中检测到较小的丰度。本研究采样时间约于白化事件发生的一个月后, 但白化珊瑚中预测的致病功能真菌丰度较少。白化状态的丛生盔形珊瑚中没有观察到珊瑚病原体显著增加, 在热胁迫下都保持与健康状态珊瑚相似的真菌群落结构, 并且白化珊瑚没有发生严重的光系统损伤和能够保持较好的光合生理状态,这暗示共生真菌群落的稳定性可能在热应激反应和珊瑚生存中发挥重要作用。
猜你喜欢
野生动物学报(2022年3期)2022-08-11
野生动物学报(2022年1期)2022-02-24
动漫星空(兴趣百科)(2020年12期)2020-12-12
中国信息化周报(2020年10期)2020-04-07
江河文学(2020年6期)2020-01-04
科普童话·神秘大侦探(2018年9期)2018-10-25
东坡赤壁诗词(2018年3期)2018-07-16
儿童故事画报·发现号趣味百科(2017年4期)2017-06-30
红蜻蜓(2014年3期)2014-07-09
读书(2014年8期)2014-01-19
