
基于OC-CCI 数据的南海高叶绿素a 浓度水域面积的时空变化研究*
2022-03-31 01:02:32李傲冯洋王云涛薛惠洁
热带海洋学报 2022年2期
李傲, 冯洋, 王云涛, 薛惠洁
1. 热带海洋环境国家重点实验室(中国科学院南海海洋研究所), 广东 广州 510301;
2. 中国科学院大学, 北京 100049;
3. 南方海洋科学与工程广东省实验室(广州), 广东 广州 511458;
4. 自然资源部第二海洋研究所, 浙江 杭州 310000
叶绿素a浓度是表征浮游植物生物量的重要指标, 是生态系统变化的指示器, 也是目前用来研究海洋动力过程环境效应的重要变量之一。在海洋生态系统中, 叶绿素a浓度的变化对区域生态系统的稳定, 海气界面二氧化碳通量的交换以及海洋水产资源的分布有着重要的影响(赵辉 等, 2005)。因此清晰地认识海洋叶绿素a浓度的时空变化对了解全球的碳循环、海区的环境质量, 以及渔业资源利用有着重要的意义。
南海是西太平洋最大的热带陆架边缘海, 位于亚洲与大洋洲、太平洋与印度洋的“十字路口”, 是典型的季风区(图1)。强劲的季风配合着复杂的地形使南海具有显著的季节性环流系统与时空多变的中尺度过程。除了局地过程的影响, 大气强迫与外源输入(黑潮)也使得南海的物理环境场存在着不同的周期性振荡。诸多因素交织, 使得南海叶绿素a浓度的时空变化在复杂的物理环境场与丰富的生物地球化学作用下更显庞杂。
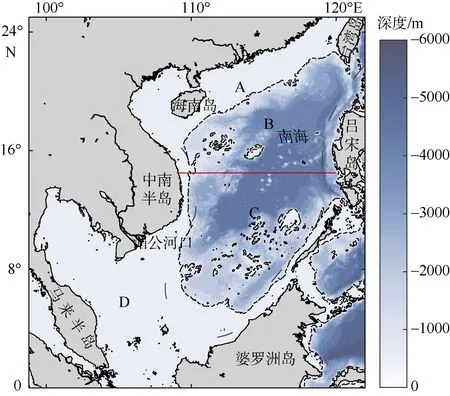
图1 南海地形分布该图基于国家测绘地理信息局标准地图服务网站审图号为GS(2020)4392 的标准地图制作。图中黑色虚线为500m 等深线,红色实线为划分南海南北的14°30′纬线。从北向南将南海划分为4 个区域, 其中A 区域为北部陆架区、B 区域为北部海盆区、C区域为南部海盆区、D 区域为南部陆架区Fig. 1 Topographic map of South China Sea. The black dotted line shows the 500 m isobath. The solid red line(14°30′N) separates the South China Sea into north and south parts
陈兴群等(1989)通过1983 年9 月—1984 年12月4 个航次数据对南海中部叶绿素a的时空分布、光合作用以及环境因子的相互关系进行了讨论, 指出叶绿素a浓度的变化与温度和盐度跃层、营养盐及光的分布有关。Ning 等(2004)通过1998 年春季与冬季的航次数据来探究南海物理生物耦合过程对浮游植物的影响, 指出浮游植物的季节分布与东亚季风驱动密切相关。其中夏季西南季风引发的上升流是湄公河口东部沿岸、海南岛东南区域、巽他陆架北侧叶绿素a浓度激增的原因, 上升流带来大量的营养盐为浮游植物的生长提供支撑。
近些年来, 随着遥感科学的快速兴起, 对南海叶绿素a浓度的时空变化研究成为可能。遥感技术凭借着大面积覆盖、快速、实时、周期性等优势成为南海资源环境监测的重要手段之一。Tang 等(1999,2003, 2004, 2006)、Chen 等(2006)先后利用不同平台遥感叶绿素a浓度资料围绕着南海近海区域叶绿素进行了研究, 指出了区域叶绿素的空间分布与变化特征。Palacz 等(2011)、Tang 等(2011, 2014, 2020)、Yu 等(2019)通过使用长时间序列南海叶绿素数据研究南海叶绿素的时空变化, 发现南海叶绿素a浓度可能存在季节内、季节间、年际以及年代际周期的振荡, 并指出这种振荡可能是由于气候变化、风应力、黑潮等因素的影响所导致。随着数值模式的发展, Xiu 等(2011, 2019)、Guo 等(2015, 2017b)、Guo等(2017a)使用遥感数据结合数值模式均从不同的侧重点研究了南海叶绿素的时空分布变化, 并通过合理的动力机制解释其成因与发展。
虽然对南海叶绿素的时空演变已有着大量的研究, 但由于船测数据成本昂贵且时间上不连续、水色遥感因云遮挡而使南海叶绿素a浓度数据缺值和插值方法等限制, 大多数研究指标仅聚焦于叶绿素a浓度, 南海表层高叶绿素a浓度水域面积的变化鲜有研究。对南海高叶绿素a浓度水域面积的时空变化分布的定量描述及其调控因子仍不明确。准确分析南海高高叶绿素a浓度水域面积对于估算浮游植物生物量变化、维持区域生态系统稳定、监测海洋渔业资源等方面都有着重要的意义。因此本文选取高叶绿素a浓度水域面积为研究指标, 参考Palacz 等(2011)根据每年冬季(最具生产力的季节)平均叶绿素a浓度峰值0.2mg·m–3来划分海表叶绿素a浓度的高低, 基于多年遥感数据资料(1999 年1 月至2019 年12 月), 对南海叶绿素a浓度高值区面积的时空变化分布进行定量描述, 同时估算出叶绿素高值区浮游植物生物量的变化, 分析了高浓度叶绿素a面积的变化特征与其调控因子(海面温度、风速等)之间的关系。
1 数据和方法
1.1 数据来源
本文使用的叶绿素资料为1999 年1 月—2019年12 月欧洲航天局(European Space Agency)海洋颜色气候变化倡议(OC-CCI) 5.0 版本的海表月平均叶绿素a数据(https://www.oceancolour.org/)。该数据结合了海洋观察宽视场传感器(Sea-viewing Wide Field of View Sensor, SeaWiFS)、中分辨率成像光谱仪(moderate-resolution imaging spectroradiometer,MODIS)、中分辨率成像频谱仪(medium resolution imaging spectroradiometer, MERIS)、可见光红外成像辐射计(visible infrared imaging radiometer, VIIRS)在内的4 种传感器测量结果。该资料空间分辨率为1/24°×1/24°。OC-CCI 对多传感器数据融合采用的大气校正算法对薄云和气溶胶显示出很好的适应效果。为了制图效果明显, 本文中叶绿素a浓度值均取以10 为底的对数。此外, 本研究还使用了1999年1 月—2019 年4 月的平均温度数据与风场数据(http://apps.ecmwf.int/datasets/)。Dee 等(2011)的研究表明欧洲中期天气预报中心(European Centre for Medium-Range Weather Forecasts, ECMWF)资料在年代际尺度变化上有着更接近实测资料的优势。选用的该资料空间分辨率为0.125°×0.125°, 本研究选取的时间范围是1999 年1 月至2019 年4 月。
研究中使用的月平均混合层深度数据为日本海洋-地球科技研究所(Japan Agency for Marine-Earth Science and Technology, JAMSTEC)开发的地球模拟器海洋环流模型数据集(Dataset of Ocean General Circulation Model for the Earth Simulator, OFES)数据(http://apdrc.soest.hawaii.edu/), 其空间分辨率为0.1°, 垂直方向为54 层, 每层的厚度参考实际海洋温跃层厚度, 其表层5m, 最底层330m, 其水深随深度的增加而逐渐变大, 由于OFES 模型混合层深度数据在时间上只到2017 年12 月, 因而2018 年与2019 年用气候态月平均混合层深度进行了补充。
1.2 研究方法
1.2.1 经验正交函数分解插值方法
本研究使用经验正交函数分解插值方法(Data Interpolating Empirical Orthogonal Functions,INEOF)对遥感观测的缺失数据进行补充, 这种方法通过确定主要的时间和空间模态对缺失数据进行填补。该方法由Beckers 等(2003)提出, 具有自适性, 该方法可以通过数据本身获取插值的必要参数。由于局部少量像元数据的改变对低阶EOF 模态几乎没有影响, 该方法首先对其中的缺失数据赋予初始预测值, 通过EOF 分解可以从大量数据中提取反映数据变化的空间模态和时间系数, 选取最佳空间模态和时间系数对数据进行重构, 完成缺失数据的插补(图2)。
本研究重构南海整体叶绿素步骤如下(图2b):
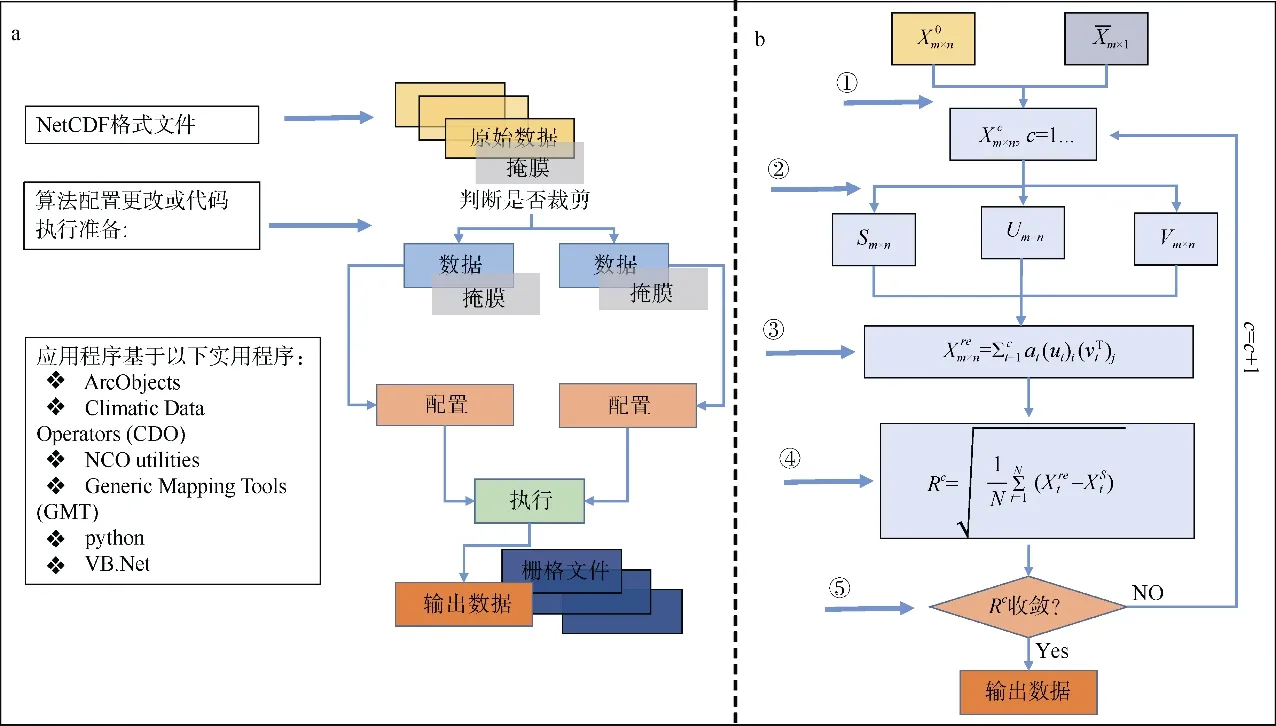
图2 经验正交函数分解插值方法(DINEOF)流程图a. DINEOF 程序运行中的参数配置与流程; b. DINEOF 方法步骤分解Fig. 2 DINEOF flow chart. (a) parameter configuration and operational procedure of DINEOF; (b) DINEOF steps

2) 对矩阵Xm×n进行奇异值分解, 分解后对应的有特征模态(Um×n)、奇异值矩阵(Sm×n)和时间特征模态(Vm×n)。

为确保DINEOF 方法能够获得有效的重构结果, 重构前, Liu 等(2019)建议将影像中高于一定缺失比例(该缺失比例设定为95%)的无法提供有用信息的单幅影像去除, 使影像集的总体缺失比例能够低于 40%; 而该方法对影像的时间连续性没有要求, 即使影像时间上分布稀疏, 该方法也能获得较好的结果。本研究中使用的是OC-CCI 月平均数据, 由于是多源遥感数据的融合产品, 这套数据在时间上连续, 且单幅影像最大缺失率小于30%, 数据缺失在空间上主要集中在南海北部与南海南部巽他陆架区域, 在时间上每年冬季的遥感数据缺失相对较高(图3)。
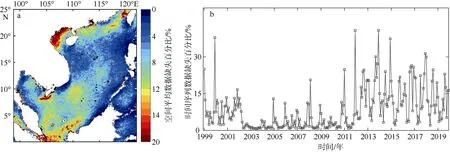
图3 OC-CCI 数据缺失百分比a. 原始数据在空间上缺失百分比; b. 原始数据在时间上缺失百分比。图 a 基于国家测绘地理信息局标准地图服务网站审图号为GS(2020)4392 的标准地图制作Fig. 3 Percentage of OC-CCI missing data. (a) the percentage of the original data missing in space, and (b) the percentage of the original data missing in time
在本次数据重构中, 随机选取了500164 个像元(有效数据总量的 1%)作为交叉检验数据(预留数据), 同时将对应位置的像元设置为缺失数据。首先保留一个EOF 模态重构缺值数据, 并计算预留数据与重构数据的均方根误差(root mean squared error, RMSE), 然后逐个增加EOF 模态保留数并不断的重构缺失数据并计算相应模态保留数对应的预留与预测数据的 RMSE。当连续两次EOF 重构的RMSE 值小于10–2时, 则认为迭代的EOF 重构收敛; 同时将EOF 模态最大值设为30,即考察保留 1~30 个模态的优劣性, 得到最优保留数。本次数据重构通过交叉检验发现最优模态为 15, 最优的迭代次数是 147, 其对应的 RMSE为0.2885 (图4)。

图4 最优重构参数a. 最优保留模态数; b. 最优迭代次数Fig. 4 Optimal reconstruction parameters. (a) the number of optimal retained modes, and (b) optimal number of iterations
1.2.2 经验正交函数分解插值方法的时空误差
将上述方法重构影像与原始资料相减并取绝对值, 获得绝对误差数据集, 求取该数据集绝对误差在时间与空间上的均值, 以考量其误差的时空分布特征(图5)。在空间维上(图5a), 影像重构误差范围在0.01~2mg·m–3之间变化, 在中央海盆误差最小, 并依次从中央海盆到沿岸呈递增趋势。大于0.5mg·m–3的值主要集中在20m 等深线以内。在时间维上(图5b),不同年份与季节之间误差波动较大, 总体看来, 秋冬季节数据重构误差明显高于春夏季节, 除了冬季月份以外, 其他时间的重构误差空间平均皆小于0.14mg·m–3, 总体误差平均值小于0.0907mg·m–3。重构误差范围主要集中在0.06~0.15mg·m–3之间, 其中2012 年冬季数据重构误差相比其他年份异常显著,通过数据检查发现这种原因可能是由于2012 年冬季数据缺失较多原因所导致的。同时为了分析重构误差对总体数据的影响情况,我们绘制了重构误差占原叶绿素a浓度数据百分比(简称重构误差百分比, 如图6), 可以发现除了南海北部区域(台湾海峡、北部陆架、北部湾)、婆罗洲岛西北出现重构误差百分比高值外, 其他区域重构误差百分比相对较小。南海平均重构误差百分比为1.46%, 重构误差百分比大于5%的面积仅占总体区域的3.47%, 因此使用DINEOF 方法插值南海缺值叶绿素数据来估算南海高叶绿素a浓度水域面积的方法具有可行性。
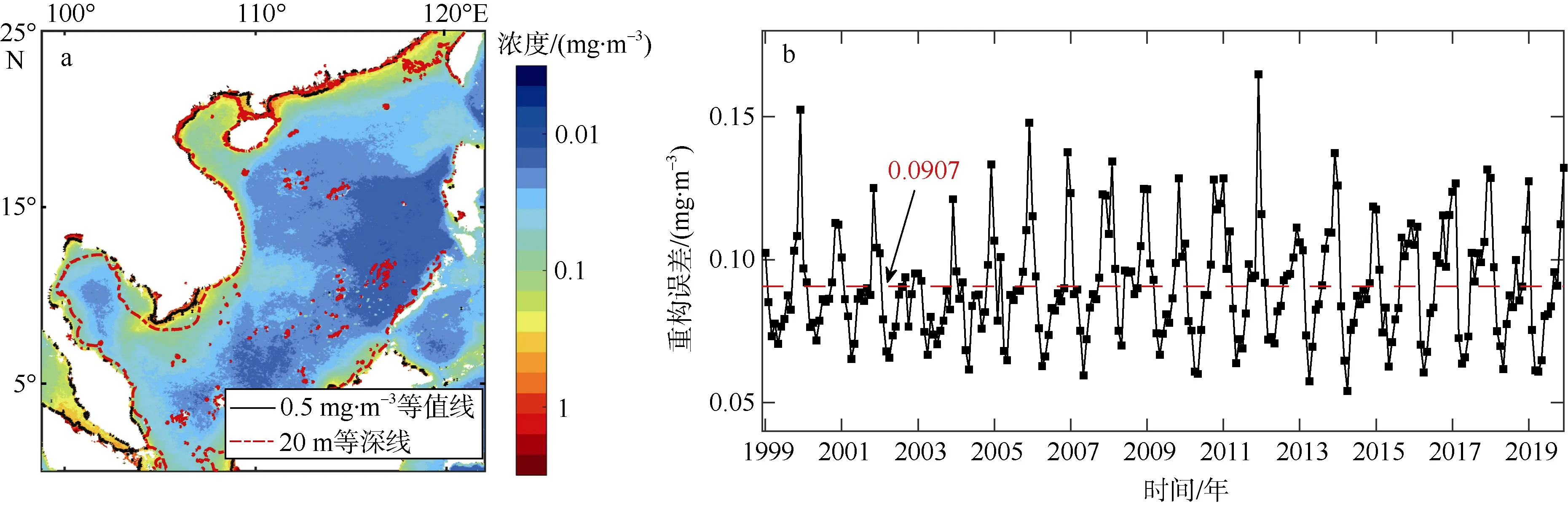
图5 重构误差的时空分布a. 重构误差的空间分布; b. 重构误差的时间分布, 图中红色虚线表示总体误差平均值。图a 基于国家测绘地理信息局标准地图服务网站审图号为GS(2020)4392 的标准地图制作Fig. 5 Spatial and temporal distribution of reconstruction errors. (a) spatial distribution of reconstruction errors; and (b)temporal distribution of reconstruction errors
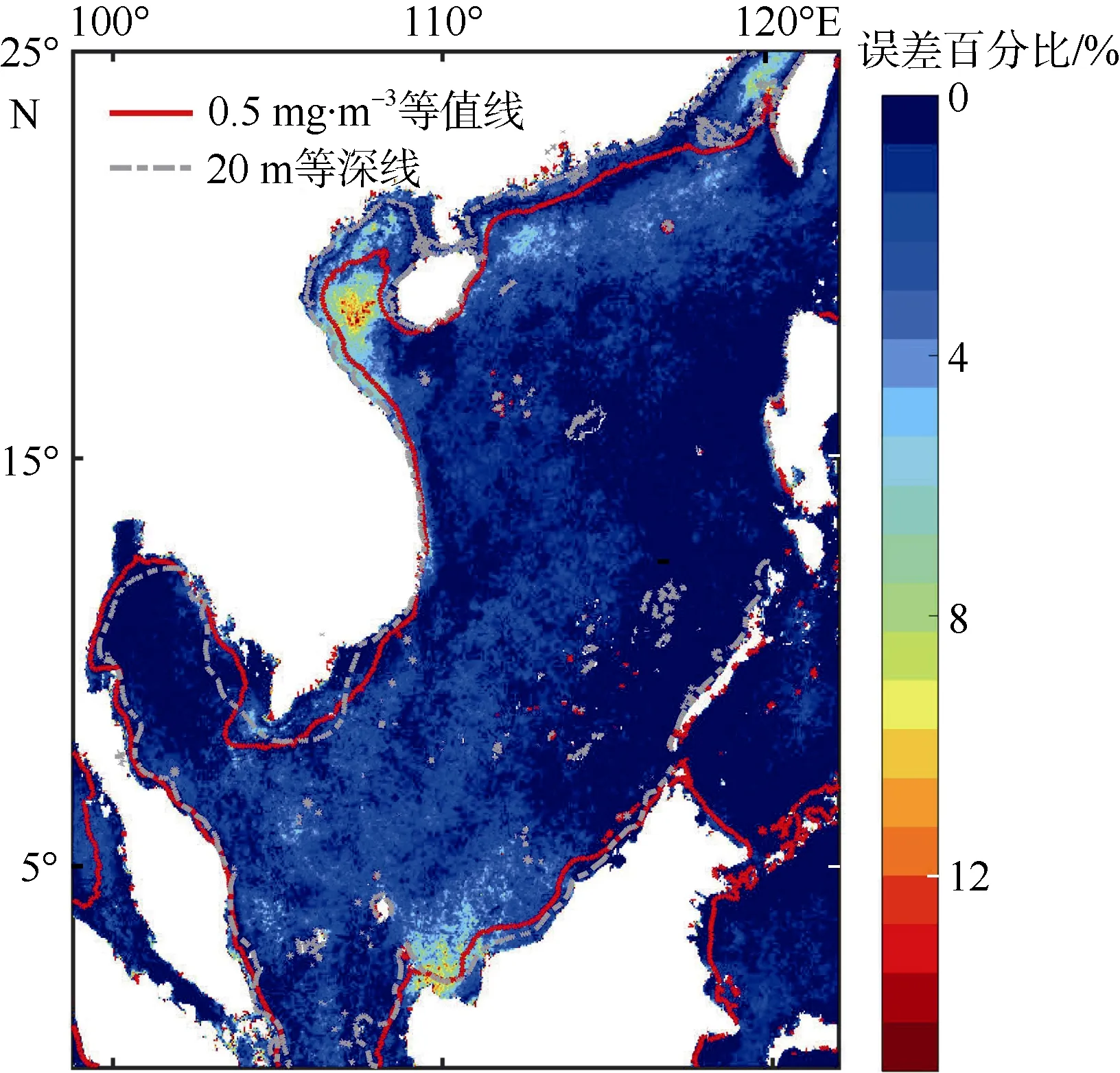
图6 重构误差占原始数据百分比Fig. 6 Percentage of reconstruction error in original data
2 结果
2.1 高叶绿素a 浓度水域空间分布特征
同Yu 等(2019)的研究结果一致, 从叶绿素的总体平均分布来看, 高浓度的叶绿素a普遍存在于南海近岸区域。图7 是南海高叶绿素a浓度水域的季节变化, 结果表明本研究中南海海表高叶绿素a浓度水域与前人研究(Tang et al, 2014; Loisel et al,2017; Huynh et al, 2020)具有相似的区域特征与季节变化, 其中高叶绿素a浓度区域普遍存在沿海地带,特别是在中国沿海、越南沿岸、泰国湾以及婆罗洲岛附近。在南海中部深水海盆区域, 叶绿素a浓度水平普遍较低(赵辉 等, 2005)。
冬季, 南海高叶绿素a浓度水域面积为所有季节中最大, 高值区显著出现在南海北部、南海南部陆架区域。春季, 南海高叶绿素a浓度叶绿素水体面积最小, 主要局限在南海近岸附近的窄带内。夏季, 高叶绿素a浓度水域面积比春季略有增加, 但海盆区域的叶绿素a浓度仍处于较低水平。与其他季节不同的是, 越南南部海域高浓度叶绿素a水体在夏季覆盖了大片海域, 可以清晰地看出从湄公河口东部向东延伸至海盆的一个高叶绿素a浓度急流正在形成。秋季, 高叶绿素a浓度水体面积相比夏季呈上升趋势, 特别是中国沿海和吕宋岛西北部地区。
2.2 高叶绿素浓度水域面积的时间变化趋势
南海整体叶绿素a月平均浓度时间序列变化为双峰结构, 在夏季处于低浓度水平, 在秋季略微抬升, 于冬季到达顶峰(Palacz et al, 2011), 这种变化趋势与南海海表高叶绿素a浓度水域面积有着非常相似的特性。图8 为南海海域(0—25°N, 98°30′—122°00′E, 其中不包含苏禄海, 苏拉威西海与马六甲海峡)内高叶绿素a浓度水域面积变化时间序列,可以看出南海高叶绿素a浓度水域面积有着非常显著的季节变化。双峰形状的变化趋势在整个时间序列都有体现, 高叶绿素a浓度水域面积在每年冬季附近到峰值, 在每年春季末与秋季末达到谷值。
前人研究发现, 南海大多区域叶绿素a浓度的特征为: 表层呈现低值, 次表层存在一个叶绿素a浓度最大值深度(Subsurface Chlorophyll Maximum,SCM), 底层浓度极低(陈兴群 等, 1989; Chen, 2005;倪晓波 等, 2006; Xing et al, 2019)。南海以热带海洋性气候为主, 温跃层随季节变化小, 大约在 50~100m 之间, 常年存在的SCM 多集中在75m 左右,基本位于真光层底部(倪晓波 等, 2006)。Xing 等(2019)使用BGC-Argo 浮标在南海南部和北部海域进行长时间序列的观测, 浮标数据表明SCM 层基本都在混合层以下, 同时资料显示出南海四季混合层内叶绿素a浓度较为均一。基于上述例证, 本文采用表层叶绿素a浓度来进行混合层浮游植物生物量的遥感估算, 通过地球模拟器海洋环流模型数据集(Dataset of Ocean General Circulation Model for the Earth Simulator, OFES)获取混合层深度资料估算区域总体生物量(图9a), 结果表明海表高叶绿素水域面积变化与该区域整体生物量有着较好的一致性。但通过与之对应的区域海表叶绿素a平均浓度(图9b)的计算发现, 冬季, 高叶绿素a浓度水域整体浮游植物生物量处于全年最大, 但其区域内叶绿素平均浓度到达全年低值; 夏季, 高叶绿素a浓度水域整体浮游植物生物量为全年最小, 其区域内叶绿素a平均浓度为全年高值。这种现象存在着季节尺度的振荡, 并伴随着显著的负相关关系(图9c)。冬季浮游植物生物量取得最大值, 这与混合作用有着密切的关系。且近岸叶绿素a浓度并不低于夏季, 但在海表高叶绿素a浓度水域面积与其对应区域总生物量都较大的时候, 海表叶绿素a浓度却很低, 表明可能是由于动力过程造成的稀释作用促使海表叶绿素重新分布。
2.3 南海不同海域高叶绿素a 浓度水域面积变化特征
南海受大陆与相连海域影响, 其不同区域水团性质在不同季节有着显著的变化, 在流场的驱动下,不同水团的相互作用与迁移直接影响着上层海洋的热力与动力结构, 从而对高叶绿素a浓度水体的空间分布产生着影响。由于南海冬季风速较强, 且参考Quan (2016)计算的南海冬季气候态Sverdrup 流函数, 发现南海中部约14°30′N 处有一零流函数等值线贯穿南海, 因此为了更好的研究不同区域高叶绿素a浓度水体的变化趋势, 我们以南海500m 等深线与北纬14°30′经线为界限, 将南海划分为4 个区域(图1), 从上到下依次分为: 北部陆架区(A)、北部海盆区(B)、南部海盆区(C)、南部陆架区(D)。
通过对不同区域高叶绿素a浓度水体面积季节时间序列变化比对(图10)可以发现, 各个区域相比较, 南部海盆区域高叶绿素a浓度水体面积最小,南部陆架区域高叶绿素a浓度水体面积最大。除了北部陆架(A)变化较为平稳外, 其他3 个区域(B、C、D)变化大致相当, 在不同年份略有波动。4 个区域的高叶绿素a浓度面积变化在冬季相对显著。自1999年起, 高叶绿素a浓度水体面积有着一个上升的趋势, 并在2007 年附近到达顶峰, 之后呈下降趋势一直持续到2018 年。

图10 南海海表不同区域不同季节高生物量水域面积的年际变化a. 北部陆架; b. 北部海盆; c. 南部海盆; d. 南部陆架Fig. 10 Interannual variation of water area with high chlorophyll a concentration in different seasons in the South China Sea
海表高叶绿素a浓度水域面积的年际变化会在海表浮游植物生物量空间分布上给予体现, 由于冬季高叶绿素浓度水域面积变化剧烈, 所以我们绘制了1999—2018 年冬季高浓度叶绿素a的空间分布(图11), 可以看出, 空间上主要分布在南海沿岸地区与南北部陆架区域, 但在巽他陆架与南部海盆区域有着较为显著的年际变化特征。在非厄尔尼诺年(如2005 年、2006 年、2010 年等)巽他陆架大面积区域被高浓度叶绿素a所裹挟, 湄公河口东部有一高浓度叶绿素a饼状区域向中央海盆扩散; 在厄尔尼诺年这些现象基本消失殆尽。这些面积变化显著的区域也是主导冬季南海总体高叶绿素a浓度水域面积呈年际变化的核心区域。

图11 南海冬季高叶绿素a 浓度水域的空间分布该图基于国家测绘地理信息局标准地图服务网站审图号为GS(2020)4392 的标准地图制作Fig. 11 Spatial distribution of water with high chlorophyll a concentration in the South China Sea in winter
3 讨论
南海高叶绿素a浓度水体面积变化受多种因素影响, 时空变化复杂。主要影响因素有: 1) 受台风等强天气过程引起的叶绿素a浓度短时间内急剧增加, 从而扩张高叶绿素a浓度水域面积(Shih et al,2020); 2) 南海冬季以东北季风主导, 夏季转变为西南风。受季风影响, 冬季上层海洋通常为气旋式环流, 而夏季环流特征为在海盆南部是反气旋式环流,海盆北部则是弱气旋式环流(Zu et al, 2019, 2020)。季风和环流直接影响上层海洋的物理环境场, 从而影响浮游植物的生长与分布, 进而影响高叶绿素a浓度水域面积; 3) 受全球气候变化而引起的浮游植物生物量年际以及年代际的振荡(Tang et al, 2011)。在高浓度叶绿素a水体空间分布上, 南海丰富的锋面、涡旋、上升流等中尺度过程对南海水团性质有着显著的影响(Wang et al, 2012), 控制着浮游植物的群落与分布, 从而改变着叶绿素分布的空间格局。Yu 等(2019)在研究南海叶绿素变化与驱动因子关系时指出, 尽管多种因子相互作用, 共同促使着叶绿素的时空格局演变, 但风场与温度场仍为影响南海叶绿素时空变化的主要影响因子。故以下对南海冬季高生物量水域面积变化分析将主要结合多年的风场、温度场来讨论。
浮游植物的生长离不开营养盐的支撑, 近岸水域水体受淡水径流补充浮游植物生长状况正常, 而南海海盆区域水体层结稳定, 真光层内因氮、磷的限制而呈现寡营养状态。Tang 等(2020)通过计算冬季浮游植物生物量、风场和混合层深度的变化趋势表明, 风驱动的混合和营养盐供应的变化可能是浮游植物生物量波动的主导因素。风场影响着表层水团输运与水体混合, 影响着营养盐的分布最终决定着浮游植物的群落与分布, 从而影响着高生物量水域的变化。我们使用了Z分数标准化方法, 观察了从1999 年至2018 年冬季南海高生物量水域面积与风速的变化关系(图12a), 发现高叶绿素a浓度水域面积与风速有着正相关关系。参照Palacz 等(2011)以气候态年平均风速7m·s–1等值线将海表风速分为高风速区与低风速区, 我们绘制了3 个年份7m·s–1风速等值线(图12b), 观察风区是否存在显著迁移。从图12b 中可以看出, 和1999 年相比, 2006 年冬季高风速区有增大趋势, 而2018 年高风速区又存在减小的势头, 与之对应的3 年(图11)南海高叶绿素a浓度面积从大到小依次顺序是 2006 年>1999 年>2018 年, 可见风速的减小以及风区的迁移可能对冬季南海表层高叶绿素a浓度水域面积变化有着重要的影响。
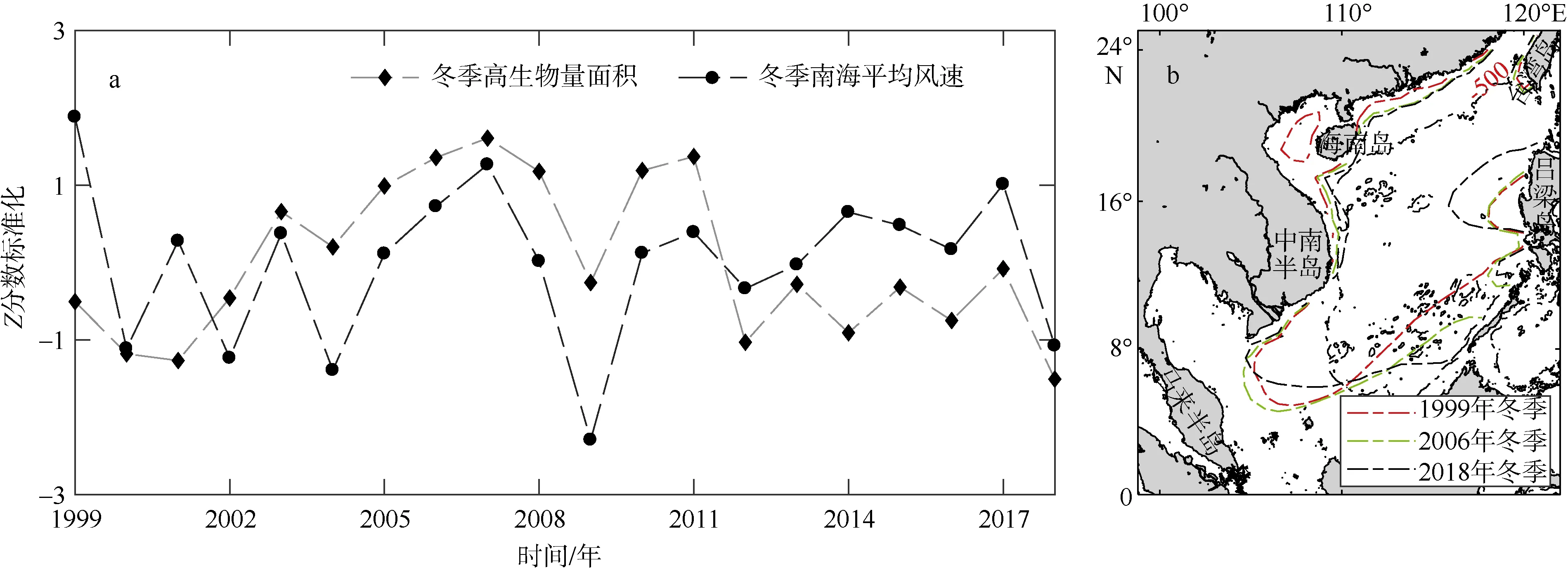
图12 南海冬季高生物量水域面积与风场的标准化时间序列(a)和不同年份海表7m·s–1 风速等值线(b)图b 基于国家测绘地理信息局标准地图服务网站审图号为GS(2020)4392 的标准地图制作Fig. 12 Standardized time series of water area with high chlorophyll a concentration and wind field in the South China Sea in winter (a); contours of sea-surface wind speed (7 m·s–1) in different years (b)
南海处于热带与亚热带的交汇处。我们通过研究不同年份南海冬季海表温度的空间分布(图13)发现冬季南海海温从南到北呈递减趋势, 在湄公河口东部附近有一冷水随南海西边界流从南海北部入侵到南海南部的现象, 称为北部冷水入侵, 且在不同年份时有出现。北部冷水入侵多出现在非EI Niño年(例如2005 年、2006 年、2010 年等)冬季, 同时与之对应的是该年份巽他陆架大面积高质量浓度叶绿素水体的出现与湄公河口东一饼状高叶绿素a浓度水域生成(图11)。为了更加直观比较厄尔尼诺年与非厄尔尼诺年的差异, 我们分别将不同年份冬季的南海高叶绿素a浓度水域(图14a、14b)与水温(图 14c、14d)进行合成分析, 同样发现在厄尔尼诺年北部冷水入侵的消失伴随着越南东南附近高叶绿素a浓度水域的消失, 在非厄尔尼诺年北部冷水入侵的出现伴随着越南东南附近高叶绿素a浓度水域的生成。

图13 南海冬季海表温度空间分布该图基于国家测绘地理信息局标准地图服务网站审图号为GS(2020)4392 的标准地图制作Fig. 13 Spatial distribution of winter SST in the South China Sea
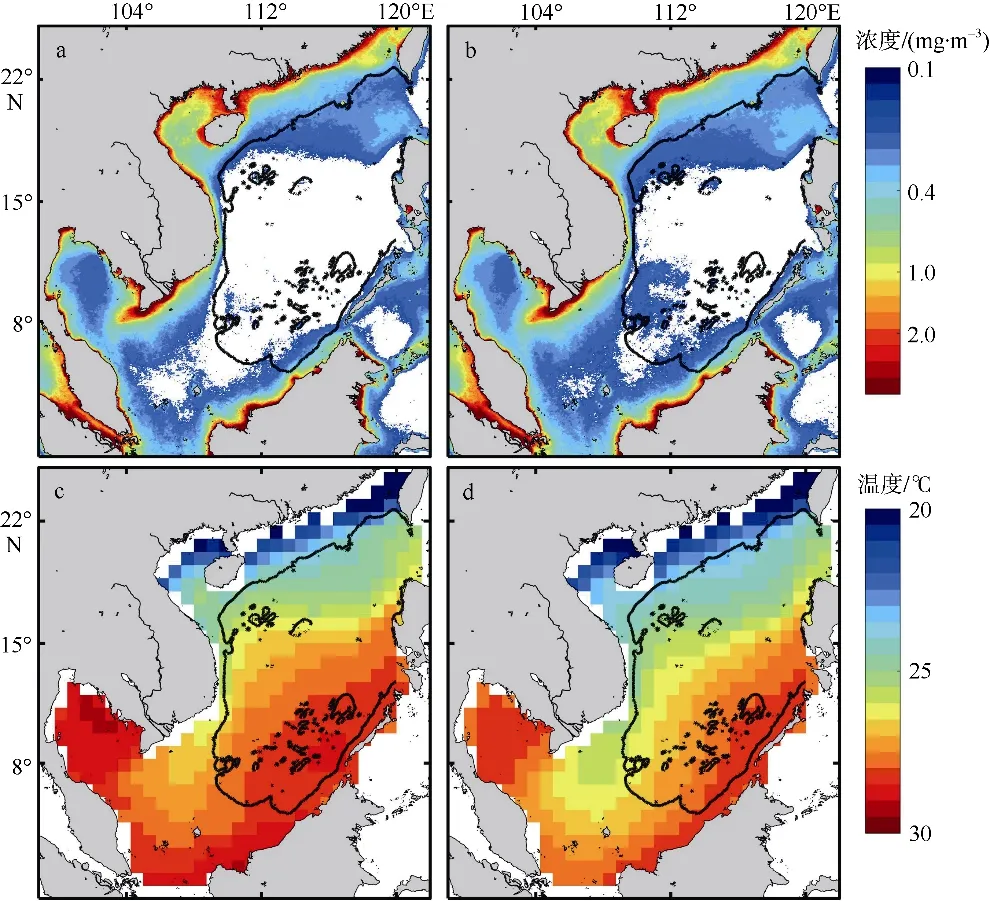
图14 南海冬季叶绿素a 浓度(a、b)和温度(c、d)的合成分析a 和c 为厄尔尼诺年; b 和d 为非厄尔尼诺年。图中黑色实线为500m 等深线。该图基于国家测绘地理信息局标准地图服务网站审图号为GS(2020)4392 的标准地图制作Fig. 14 Synthesis analysis of winter SST and chlorophyll a in the South China Sea
对于这种北部冷水入侵强度差异的南海西边界现象, Quan 等(2016)也指出受到盛行季风以及出、入流的影响, 西边界流的结构和强度有显著的年际变化特征, 在大部分厄尔尼诺年的冬季, 南海西边界流变得较深、较窄、较弱。厄尔尼诺通过调控南海季风的年际变化来调控南海西边界流年际变化是这种空间格局形成的主要因素(Zu et al, 2019)。因此,北部冷水入侵的年际变化可能是引起不同年份冬季局地浮游植物生物量增减的重要原因之一。
4 结论
本文基于OC-CCI 数据, 运用DINEOF 插值方法重构了南海叶绿素场, 基于面积指标建立了估算南海浮游植物生物量的方法体系, 结合海表温度、海面风场等环境要素的时空分布, 从时间和空间尺度分析了南海高叶绿素a浓度水域面积的变化特征。
总体来说, 在空间分布上, 离海岸越近, 重构误差值就越大。从时间分布上看, 每年除冬季外, 其他月份重构误差的空间平均值皆小于0.14mg·m–3,总体误差平均值为0.0907mg·m–3。除了南海北部部分区域(台湾海峡、北部陆架、北部湾)、婆罗洲岛西北出现重构误差占叶绿素a浓度百分比出现高值外, 其他区域重构误差百分比占比相对较小, 且重构误差百分比大于5%的面积仅占总体区域的3.4%,因此可以认为本次重构误差结果控制在一个较好的水平。
在时间序列上, 重构的高叶绿素a浓度水域面积有着显著的季节性特点。在秋、冬季节, 面积到达双峰结构高值, 其中秋季的峰值相对冬季较小;在夏季到达全年面积的最小值。季节变化主要影响因素是风场, 风速的减小以及风区的迁移可能对冬季南海表层高叶绿素a浓度水域面积变化有着重要的影响。对于冬季海表高叶绿素a浓度水域面积大而浓度低, 夏季海表高叶绿素a浓度水域面积小而浓度大的主要可能原因是风驱动海表动力过程促使海表叶绿素重新分布。
在空间分布上, 高叶绿素a浓度水体常年存在于海岸附近, 特别是在中国沿海、越南沿岸、泰国湾以及婆罗洲岛附近。不同年份(如2003 年、2005年、2006 年、2007 年、2008 年、2010 年、2011年、2017 年)的冬季, 在南海中部深盆区域以及巽他陆架存在着异常变化。这种变化主要可能是由于厄尔尼诺调控下的南海季风出现年际变化所导致的, 季风的年际变化影响着西边界流, 从而影响着冬季南海西边界冷水的侵入, 导致南海南部深水海盆区域以及巽他陆架处高叶绿素a浓度水域的变化。
由于本文是基于由遥感资料得到的叶绿素a浓度来进行分析研究的, 缺乏实测航次资料对比。文中通过地球模拟器海洋环流模型数据集(OFES)混合层深度数据估算南海高生物量水域混合层生物量, 可能会因为不同季节次表层叶绿素最大值深度变化, 使得海表叶绿素a浓度估算南海整体混合层生物量得到的结果可能与实际之间会有一定的差异。本文通过遥感数据初步探讨了南海高叶绿素a浓度水域面积的时空变化以及主要影响因素,但是其时空变化的响应机制及动力过程仍有待于进一步研究与发现。
猜你喜欢
小猕猴智力画刊(2022年4期)2022-05-23 13:48:22
黄河之声(2021年10期)2021-09-18 03:07:18
水上消防(2020年5期)2020-12-14 07:16:14
阅读(科学探秘)(2020年8期)2020-11-06 06:22:48
文史春秋(2019年11期)2020-01-15 00:44:36
中国果业信息(2019年1期)2019-01-05 17:41:42
智能城市(2018年7期)2018-07-10 08:30:00
生物学教学(2017年9期)2017-08-20 13:22:32
军营文化天地(2017年1期)2017-03-06 01:34:23
草原(2016年1期)2016-01-31 21:21:51
