
基于UPLC-QE-MS探究褐变反应对酸乳中代谢物的影响
2022-03-30 02:13:20宁雪楠康佳欣廖敏和任皓威
食品科学 2022年6期
李 萌,宁雪楠,康佳欣,廖敏和,任皓威,刘 宁
(东北农业大学食品学院,乳品科学教育部重点实验室,黑龙江 哈尔滨 150030)
酸乳是以生牛(羊)乳或乳粉为原料,经杀菌后添加乳酸菌发酵而成的乳制品。因其口感独特、营养丰富、易于人体消化而备受消费者喜爱[1]。除了营养特性以外,风味和颜色也被作为乳制品选择的关键因素之一。在这种情况下,市场上出现了一种新兴的酸乳产品,即褐色酸乳[2]。褐色酸乳又称熟酸乳、烤酸乳、炭烧酸乳[3]。与普通酸乳相比,褐色酸乳具有咖啡般的颜色和焦糖般的风味,这种特殊的颜色和风味主要由褐变反应引起。褐变反应包括酶促褐变和非酶促褐变,非酶促褐变一般包括美拉德反应、抗坏血酸氧化、多酚氧化缩合、焦糖化反应[4],其中美拉德反应被认为是褐色酸乳风味和颜色变化的主要原因[5],其反应机理是还原糖的羰基与蛋白质、氨基酸的氨基发生非酶促褐变反应[6-7]。
目前,关于褐色酸乳的研究主要集中于褐变工艺。张俊山[8]探究了不同还原糖对美拉德反应程度的影响,通过色度值分析发现葡萄糖的褐变效果最好。王晓倩等[9]对褐色酸乳的褐变工艺进行优化,最终发现在葡萄糖添加量8%、褐变温度98 ℃、褐变时间150 min条件下生产出的褐色酸乳颜色及口感最好。关于褐色酸乳在代谢组学方面的信息鲜见报道,褐变反应会对酸乳的风味及颜色有所影响。代谢组学作为一种高通量的检测方法,主要对样品中的小分子物质进行全面定量和定性技术[10],现已广泛用于乳制品研究中,Caboni等[11]采用气相色谱质谱-非靶向代谢组学研究了热处理对奶酪代谢物的影响,结果表明经热处理后羊奶发酵生产出的奶酪半乳糖、葡萄糖酸等物质的相对含量显著上调,α-羟基丁酸的相对含量显著下调。Rubert等[12]采用超高效液相色谱-质谱-非靶向代谢组学研究了热加工对油莎草(Cyperus esculentusL.)乳成分的影响,结果表明新鲜油莎草乳中生物素和精氨酸的相对含量高于热加工的油莎草乳,这些研究说明可以用代谢组学的方法分析乳制品中小分子代谢物的变化。目前,代谢组学中关于褐色酸乳的研究较少,褐变反应是影响褐色酸乳风味和颜色变化的主要原因,因此本研究以普通酸乳和褐色酸乳为研究对象,利用超高效液相色谱-四极杆静电场轨道阱质谱(ultra-high performance liquid chromatographyquadrupole electrostatic field orbitrap mass spectrometry,UPLC-QE-MS)对2 种酸乳进行非靶向代谢组学分析,建立2 种酸乳的代谢图谱,同时探究褐变反应对酸乳中小分子代谢物的影响。
1 材料与方法
1.1 材料与试剂
生牛乳 市购;乳糖酶 沧州夏盛酶生物技术有限公司;白砂糖 乐陵市厨大妈食品有限公司;直投式发酵菌粉YO-MIX883 丹尼斯克(中国)有限公司;甲醇、乙腈、乙酸铵、氨水(均为分析纯) 德国CNW公司;超纯水 香港屈臣氏集团有限公司;无水乙醇湖北福星生物科技有限公司。
1.2 仪器与设备
Vanquish超高效液相色谱、Q Exactive HFX高分辨质谱仪、Heraeus Fresco17离心机 美国Thermo Fisher Scientific公司;JXFSTPRP-24研磨仪 上海净信科技有限公司;PS-60AL超声仪 深圳市雷德邦电子有限公司;SPL-150生化培养箱、ZQPW-70全温培养振荡器天津市莱玻特瑞仪器设备有限公司;HH-4数显恒温搅拌水浴锅 常州赛普实验仪器厂。
1.3 方法
1.3.1 发酵乳的制备
分别向2 个相同的生牛乳样品中加入0.814%的乳糖酶并置于52 ℃培养箱中,50 min后将牛乳取出并置于96 ℃水浴锅中15 min,使乳糖酶失活,将其中一个牛乳样品放在94.4 ℃水浴锅中使其转变为褐色牛乳,212 min后将其取出,另一个作为普通牛乳;将普通牛乳和褐色牛乳接种YO-MIX883发酵剂,此发酵剂的菌种为保加利亚乳杆菌和嗜热链球菌,接种量为万分之四,然后在42 ℃条件下恒温培养,待pH值到达4.6时将酸乳取出,4 ℃后熟18 h。
1.3.2 样品前处理
代谢物的提取参考Cai Yuping等[13]提出的方法并稍作改动,将2 种酸乳样品真空冷冻干燥,取20 mg研磨成粉后的样品并加入1 000 μL萃取溶液(甲醇-乙腈-水,2∶2∶1,V/V,带有同位素标记的内标混合物),涡旋混匀4 min,随后在冰水浴中超声5 min。匀浆及超声处理重复3 次。将样品在-40 ℃孵育1 h,然后在4 ℃、12 000 r/min离心15 min,将得到的上清液转移新瓶中进行上机分析,并通过等分混合2 种样品的上清液制备质量控制样品,每个样本的生物学重复为6 个。
1.3.3 UPLC-QE-MS检测
UPLC条件:ACQUITY UPLC BEH Amide色谱柱(2.1 mm×100 mm,1.7 μm);流动相:A为水相(含25 mmol/L乙酸铵和25 mmol/L氨水),B为乙腈;样品盘温度4 ℃,进样体积3 μL。
MS条件:选择QE HFX质谱仪,采用电喷雾电离源,在正离子和负离子模式下采集数据。正离子模式下喷雾电压为3 600 kV;负离子模式下喷雾电压为-3 200 kV。鞘气体流量为30 arb;辅助气体流量为25 arb;毛细管温度为350 ℃;全MS分辨率为60 000,MS/MS分辨率为7 500;轰击能量为10/30/60 eV。
1.4 数据处理
原始数据经ProteoWizard软件转成mzXML格式后,使用XCMS进行峰识别、峰提取、峰对齐和积分等处理,然后与BiotreeDB(V2.1)二级质谱数据库匹配进行物质注释,算法打分的Cutoff值设为0.3。采用SIMCA软件(V16.0.2, Sartorius Stedim Data Analytics AB, Umea,Sweden)对UPLC-QE-MS的数据进行多元统计分析,主要包括无监督的主成分分析(principal components analysis,PCA)和有监督的正交偏最小二乘判别分析(orthogonal partial least squares-discriminant analysis,OPLS-DA)。
PCA主要研究样本分布、偏差特征和共同趋势,OPLS-DA主要是对样本进行分类,并对大多数判别变量进行识别,并基于拟合优度和预测优度验证分类和预测能力,通过置换实验(n=200)对模型进行过拟合检验,所得截距(Q2)的负值表明模型的稳健性。变量投影重要度(variable importance in the projection,VIP)得分显示每个变量对模型的贡献,对预测成分中的VIP得分进行分析,只有VIP值大于1的代谢物才被视为类别之间的鉴别[14]。最后将得到的代谢物信息根据VIP值、Studentt检验的P值和差异倍数(fold change,FC)以2为底的对数值进行筛选,从而得到具有显著差异的代谢物。
2 结果与分析
2.1 UPLC-QE-MS代谢图谱分析
基于UPLC-QE-MS分别在正负离子模式下对普通酸乳和褐色酸乳进行检测,得到2 种酸乳样品的总离子流图(total ion current,TIC)。由图1可知,普通酸乳和褐色酸乳的基线稳定,说明仪器数据采集稳定性很好。正负离子模式下TIC出峰的保留时间、峰面积以及峰的数量有所不同,因此需要在后续对原始数据进行处理分析。

图1 普通酸乳和褐色酸乳随机单个样品的TICFig. 1 Total ion current chromatograms of random single samples of ordinary yogurt and brown yogurt
2.2 PCA结果
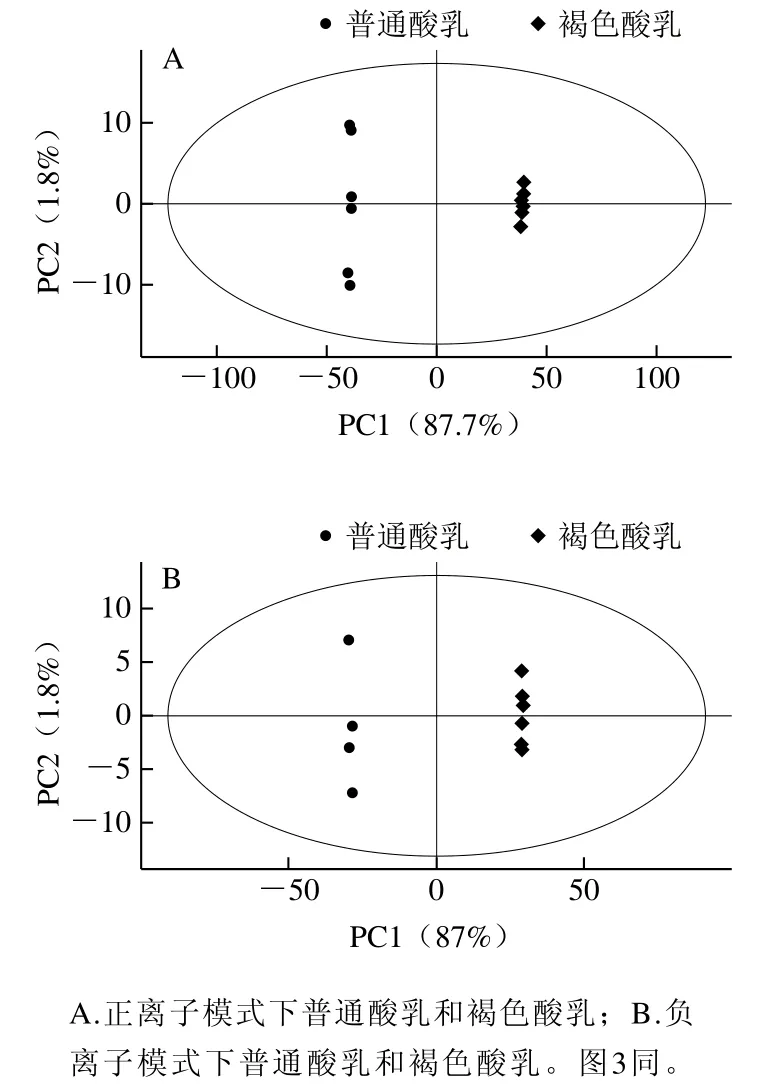
图2 普通酸乳和褐色酸乳PCA模型得分图Fig. 2 PCA score plots of ordinary yogurt and brown yogurt
由图2可以很明显地看出,在正离子和负离子模式下2 种酸乳样品有明显分离趋势,正离子模式下PCA模型的负离子模式下PCA模型的PCA模型的代表模型对X变量的解释性,当它的值大于0.4时,说明此模型可靠,能够用于解释普通酸乳和褐色酸乳之间的代谢物差异[15]。
2.3 OPLS-DA结果

图3 普通酸乳和褐色酸乳的OPLS-DA模型图及置换检验图Fig. 3 OPLS-DA model plot and permutation test plot of ordinary yogurt and brown yogurt
为了更好地对数据进行可视化及后续分析,采用OPLS-DA对数据进一步分析,可以过滤掉代谢物中与分类变量不相关的正交变量,并分别对正交变量和非正交变量分析,从而更加准确地识别代谢物的差异[16]。如图3A、B所示,2 种酸乳样本能够明显地分开,所有样本均处在95%置信区间的椭圆内。正离子模式下模型Q2=0.993;负离子模式下模型代表模型对Y变量的解释性,Q2代表模型的可预测性,非常接近1,说明建立的模型符合样本数据的真实情况。原模型Q2非常接近1,说明如果有新样本加入模型,会得到近似的分布情况,因此该模型可以很好地解释两组样本之间的差异[17]。为了防止模型出现过度拟合,对该模型进行200 次置换检验,最终结果如图3C、D所示,正离子模式下置换检验模型Q2=-0.96;负离子模式下Q2=-0.98。置换检验随机模型的Q2值均小于原模型的Q2值,说明原模型具有良好的稳健性,不存在过拟合现象。
2.4 差异性代谢物的筛选与鉴定
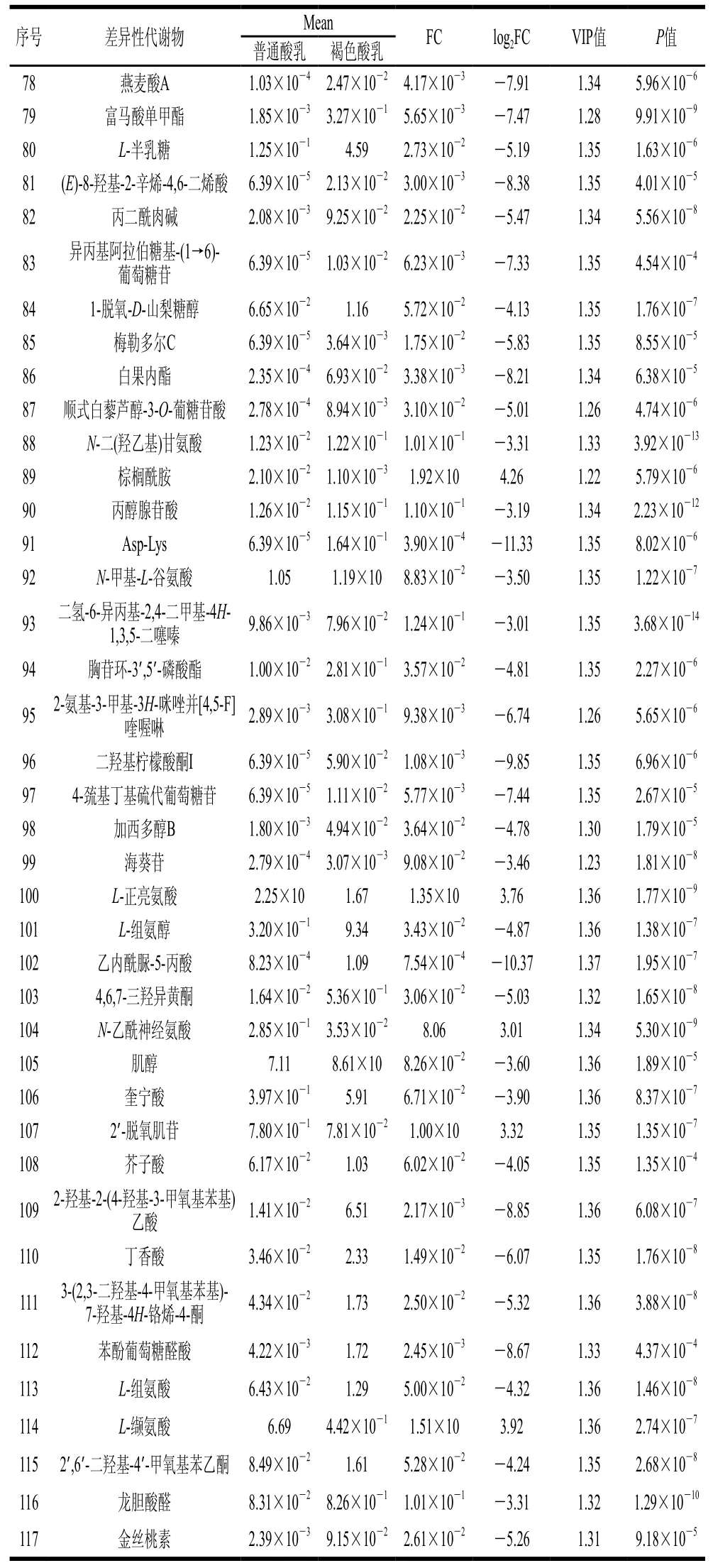
为了在数百个变量中找到比较重要的差异性代谢物,本研究根据VIP值大于1.2、Studentt检验的P值小于0.001、log2FC的绝对值大于3的筛选原则进行进一步筛选,最终共得到117 种代谢物,这些代谢物的具体信息如表1所示,主要包括氨基酸、肽、糖类和风味化合物等,2 种酸乳的主要差异是颜色与风味,因此本研究主要是对酸乳颜色及风味有影响的代谢物进行讨论。

表1 普通酸乳和褐色酸乳差异性代谢物Table 1 Differential metabolites between ordinary yogurt and brown yogurt

续表1
续表1
2.4.1 氨基酸和二肽
如表1所示,与普通酸乳相比,褐色酸乳中异亮氨酸、缬氨酸的相对含量下调,组氨酸的相对含量上调。缬氨酸与异亮氨酸是疏水性氨基酸,大多数疏水性氨基酸都有苦味[18]。除了氨基酸的相对含量发生变化外,普通酸乳和褐色酸乳中共有8 种二肽有显著差异,分别为京都啡肽、焦谷氨酰胺基谷氨酰胺、Asp-Lys、Phe-Ala、Asp-Thr、Pro-Asp、Glu-Pro和N-γ-谷氨酰谷氨酰胺。这8 种二肽在褐色酸乳中的相对含量高于普通酸乳,其中京都啡肽、焦谷氨酰胺基谷氨酰胺在2 组酸乳中相对含量的FC显著高于其他6 种二肽,京都啡肽是一种内源性镇痛神经肽[19],虽然鲜见其在乳制品中生成机理的研究报道,但根据本研究结果可以推测,褐变反应可能对酸乳中京都啡肽的生成具有促进作用。此外,褐色酸乳中京都啡肽的相对含量是普通酸乳中的13 975 倍,它是117 种差异性代谢物中FC最大的代谢物,可作为褐色酸乳潜在的标志性代谢物。焦谷氨酰胺基谷氨酰胺是一种能够赋予食品一定鲜味的味觉化合物,褐色酸乳中焦谷氨酰胺基谷氨酰胺的相对含量是普通酸乳中的3 096 倍,这可能是因为牛乳在加热过程中,焦谷氨酸和游离氨基酸在焦谷氨酰环化酶的作用下生成焦谷氨酰胺基谷氨酰胺[20],相比于普通酸乳,褐色酸乳在发酵前会经过更长时间的热处理,因此褐色酸乳中焦谷氨酰胺基谷氨酰胺的相对含量高于普通酸乳。这些氨基酸和二肽相对含量的变化可能是赋予褐色酸乳特殊风味的原因之一。
2.4.2 糖类
糖类是乳酸菌发酵的主要碳源,可为微生物的生长繁殖提供能量[21]。与普通酸乳相比,褐色酸乳中共有12 种糖类的相对含量显著升高,糖类含量的升高可能是褐变反应和酸乳中微生物发酵代谢所致[22]。由表1可知,褐色酸乳中5-氨基咪唑核苷酸和6-磷酸果糖的相对含量分别是普通酸乳的464 倍和289 倍。5-氨基咪唑核苷酸是乳酸菌核酸代谢途径中重要的中间产物,其经过羧基化最终会形成腺嘌呤核苷酸和鸟嘌呤核苷酸,它们是发酵食品中重要的风味核苷酸,对酸乳的风味和香气有一定贡献[23]。有研究表明,牛乳在热处理过程中会生成半乳糖和吡喃半乳糖基甘露糖[24]。半乳糖会经过Leloir途径最终转化为葡萄糖[25],葡萄糖在糖酵解过程中会生成6-磷酸葡萄糖,并在己糖激酶的作用下转化为6-磷酸果糖[26]。因此褐色酸乳中6-磷酸果糖的相对含量显著升高。这些糖含量的升高有助于褐色酸乳甜味的提升。
2.4.3 风味化合物和其他类物质
由表1可知,褐色酸乳中2-乙酰基呋喃和5,6-二氢-4-甲氧基-2H-吡喃-2-酮的相对含量显著升高,分别是普通酸乳中的1 555 倍和13 倍,它们都是美拉德反应过程的中间体或产物。2-乙酰基呋喃是许多热处理食品中的风味成分,它的气味阈值很低,是一种具有甜味、杏仁味和香醋味的风味化合物,对食品的感官特性有显著的贡献[27]。5,6-二氢-4-甲氧基-2H-吡喃-2-酮是吡喃酮的衍生物,它能够赋予食物焦糖般的香气。由表1可观察到,褐色酸乳中苯醌相对含量是普通酸乳中的2 113 倍,这可能是牛乳褐变过程中发生了多酚氧化缩合反应,这种反应是以酚类为底物,在高温、潮湿条件下将酚类氧化为苯醌,从而使牛乳的颜色变深[28-29]。因此可以推断,褐色酸乳的颜色除了受美拉德反应影响外,可能在一定程度上也与苯醌有关。同时,苯醌可能会与其他氨基酸发生反应,进而生成一些风味化合物(醛、胺等)[30]。与普通酸乳相比,褐色酸乳中酚类、醇类、酮类、酯类、萜类等物质的相对含量显著上调,它们是酸乳中风味物质的前体,同时还具有一些有益功能,对酸乳的风味及营养功能有很大的贡献。
3 结 论
本研究考察褐变反应对酸乳中小分子代谢物的影响,2 种酸乳代谢物的相对含量有显著差异,这些代谢物主要包括氨基酸、二肽、糖类、风味化合物等。褐色酸乳中糖类、风味化合物等物质的相对含量普遍升高,这可能是引起2 种酸乳风味不同的主要原因。与普通酸乳相比,苯醌在褐色酸乳中的相对含量显著上调,这说明褐色酸乳在生产过程中除了会发生美拉德反应外,还可能发生多酚氧化缩合反应,反应产生的苯醌可能对褐色酸乳的风味及颜色有所影响。酚类、醇类、酮类、萜类等在褐色酸乳中含量较高,对酸乳的风味有一定贡献作用。京都啡肽是117 种差异性代谢物中含量差异最大的代谢物,可作为褐色酸乳潜在的标志性代谢物,本研究结果可以为褐色酸乳的研究与开发提供一定理论依据。
猜你喜欢
食品安全导刊(2021年21期)2021-08-30 08:22:06
北方人(2020年22期)2020-12-02 08:11:02
疯狂英语·新悦读(2020年4期)2020-06-18 05:35:28
自我保健(2020年8期)2020-01-01 21:12:03
阅读(快乐英语中年级)(2019年11期)2019-09-10 07:22:44
儿童时代·幸福宝宝(2017年1期)2017-06-29 22:29:29
中国酿造(2016年12期)2016-03-01 03:08:23
食品工业科技(2014年23期)2014-03-11 18:19:04
食品工业科技(2014年15期)2014-03-11 18:17:29
食品工业科技(2014年15期)2014-03-11 18:17:18
