
Cob color, an indicator of grain dehydration and agronomic traits in maize hybrids
2022-03-30 08:52:24XiaoGuiLiangZhenGaoSiShenXueZhaoLiZhangShunLiZhou
The Crop Journal 2022年2期
Xiao-Gui Liang, Zhen Gao, Si Shen, Xue Zhao, Li Zhang, Shun-Li Zhou,c,d,*
a Key Laboratory of Crop Physiology, Ecology and Genetic Breeding, Ministry of Education/Jiangxi Province, Jiangxi Agricultural University, Nanchang 330045, Jiangxi, China
b College of Agronomy and Biotechnology, China Agricultural University, Beijing 100193, China
c Scientific Observation and Experimental Station of Crop High-Efficiency Use of Water in Wuqiao, Ministry of Agriculture and Rural Affairs, Wuqiao 061802, Hebei, China
d Innovation Center of Agricultural Technology for Lowland Plain of Hebei, Wuqiao 061802, Hebei, China
Keywords:Maize hybrids Cob color Grain dehydration Agronomic traits Artificial selection
ABSTRACT Screening for agronomic traits associated with grain moisture is important for mechanical grainharvesting of maize.Cob color as a visual indicator has received limited attention,though it has been subjected to artificial selection, and may have some association with threshability. To investigate the relationships between cob color and grain moisture and other agronomic traits, field experiments were conducted during 2016–2017 using 23 commercial hybrids with red or white cobs. Kernels of red-cob hybrids dehydrated faster, showing lower moisture content at harvest than white-cob hybrids. A cob color index(CCI)was established as a quantitative measure of cob color in the hybrid panel.Ranging from 0 (whitest) to 17.98 (reddest), CCI correlated well with grain dehydration and other agronomic traits associated with growth ontogeny, plant morphology, and plant N content. Strong selection of red cob for recently released hybrids suitable for mechanical grain-harvesting indirectly validated the observed link between cob color and grain dehydration.We propose that cob color and CCI could be used in future selection of maize cultivars bred for mechanical grain-harvesting.
1. Introduction
As one of the world’s most important crops, maize provides human food, livestock feed, and industrial materials. In China,scientists are promoting mechanical grain-harvesting to replace the traditional ear-harvesting and low-profit maize production[1–3]. One of the major obstacles is the high grain moisture at harvest of conventional maize hybrids [4–6]. Identifying hybrids with low grain moisture at harvest and related agronomic traits would facilitate mechanical grain-harvesting[7,8]. Rapid grain dehydration could be associated with ear traits such as early maturity, husk length and numbers, grain size, pericarp thickness, endosperm color, and cob diameter[8–11]. As an ear trait, cob color shares the same locus (Pericarp Color 1, P1) but different alleles with pericarp and endosperm color [12–14]. It is not known whether cob color is correlated with grain dehydration.
Cob color is generally classified as red(colored)and white(colorless). It has been proposed [15] that the P1 allele of red cob has been artificially selected since domestication for currently unidentified improvements in modern temperate maize lines. Empirical observations of experienced maize scientists[5,14,15]suggest that red cob is correlated with mold resistance,early maturity,and cob hardness, making it suitable for mechanical grain-harvesting. This experience has been obtained in part from the three most popular maize hybrids in China during the last 20 years. Nongda 108 and Zhengdan 958 (with white cob) are late-maturing and stay green at harvest, while Xianyu 335 (with red cob) is often criticized for early senescence. Early maturity may lead to low grain moisture at harvest and affect the migration of plant C and N [5,16–18].However, there are no reliable reports of the correlation between cob color and early maturity, grain dehydration, or other traits.In addition, cob color is often graded. on a binary basis, as red or white. This practice would obscure the depth and dimension of the color, hiding the potential association between cob color and agronomic traits.
The development of cob color as an indicator for maize selection has not been well studied, in particular in commercial hybrids in relevant field environments.We hypothesized that maize cob color would be associated with grain dehydration and other agronomic traits,and set out to quantify cob color with the goal of developing it as a visual selection tool.Our experimental approach was to verify the connection between early maturity and red cob and to create a cob color index that would allow prediction of grain dehydration and other agronomic traits.
2. Materials and methods
2.1. Experimental site, materials, and design
The experiment was conducted at the Wuqiao Experimental Station of China Agricultural University in Hebei in 2016–2017.Weather data for the two summer maize seasons from June to October are shown in Fig. S1. Soil conditions and fertilization are described in Table S1. Twenty-three commercially released hybrids, including 12 white- and 11 red-cob hybrids, which are currently or have been historically widespread in China,were used to collect two years of experimental data. The hybrids were planted in a randomized complete block design in three replications in the field (Table S2; Fig. S2). Each plot was planted in six rows in an area of 32.4 m2at a density of 67,500 plants ha-1.According to local practice, maize was planted on June 15, 2016 and June 16, 2017, and the regular harvest was taken on October 3, 2016 and October 2, 2017. Field management followed local farmer practices.
2.2. Agronomic traits and yield determination
Shoot dry weight (DW) and leaf area index (LAI) were measured at the stages of 6th leaf expanded (6-LS), 13th leaf expanded (13-LS), silking (SS), and 15, 28, 42, and 53 days after pollination (DAP) in both seasons. Plant height and ear height were measured after pollination. Three plants from each plot were used for measurement of green leaf area using the following formula: leaf length × width × 0.75, and LAI was calculated as the total green leaf area per unit area of soil [19]. Samples were separated into leaves, stems, and grain (or vegetative and reproductive tissues), dried in an oven at 70 °C, weighed, and ground to fine powder for N concentration determination by the Kjeldahl method. N accumulation amount (Naccu) was calculated for tissues and for whole plant. Silking date and physiological maturity date were determined by visible silk and black layer attained in 50% of ears, respectively. Anthesis silking interval (ASI) was calculated. For yield and yield traits, approximately six m2(2 rows × 5 m) of ears were harvested at maturity. Ear length, ear rows, and kernels per row were measured. Grain was then dried at 70 °C to determine % moisture and thousand-kernel weight. Yield was standardized to 14% moisture.Based on the above measurements, the following C and N efficiency were calculated following Chen et al. [20]:

Potential grain-filling rate was measured in 2017.One hundred kernels from central parts of three cobs from each plot were sampled. Data of fresh and dry weight and grain volume were measured at 14 and 28 DAP. The relative potential grain-filling ratio(RPGR) for each hybrid was calculated as:
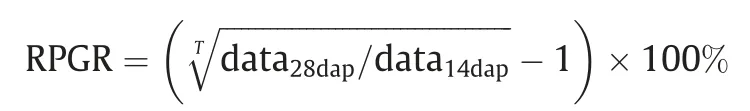
where, T is the interval in days between samples [20,21]. Grain moisture was measured at 14 and 28 DAP,and at 0,10,and 20 days after regular harvest time (DARH). Dynamics of grain dehydration were fitted by piece-linear and nonlinear curves. Thirty kernels from the central parts of three cobs from each plot were sampled to measure grain hardness at 0,10,and 20 DARH.Grain shear forces for a kernel position of the top, lateral and venter of single kernel were determined with a microcomputer-controlled electronic universal testing machine (RGM-6005, REGER, Shenzhen, Guangdong,China) as described previously [22].
2.3. Quantification of cob color
To quantify cob color,three cobs from each plot were ground to approximately 50 mesh, placed in wells of a 24-well plate to greater than 2 mm depth and scanned in a scanner(HP LaserJet M227, 600 bpi, HP, Palo Alto, CA, USA). Adobe Photoshop CS6(Adobe Systems, San Jose, CA, USA) was used to assign values to cob color by the L*a*b*model according to experience with cotton color [23,24]. L*, a* and b* indicate the color from black to white,from green to red, and from blue to yellow, respectively. The cob color index (CCI) was calculated as:
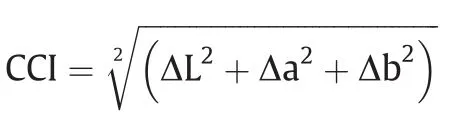
where Δ is the difference between sample and control. The maize hybrid ZD-958 was closest to white,with the lowest summed value of L*, a* and b*, and was used as the control in this study.
2.4. Statistical analysis
Microsoft Excel 2019(Microsoft,Redmond,WA,USA)was used for data standardization and visualization. IBM SPSS 20 (IBM,Armonk, NY, USA) was used for calculating ANOVA and Pearson correlations.
3. Results and discussion
3.1. Differences and potential driving factors in grain dehydration between red- and white-cob hybrids with similar yields
Among the yield traits,there were significant differences in ear diameter and harvest index between the red- and the white-cob hybrids in both seasons (Fig. S3A, B). However, final yields were similar between reds and whites (Fig. S3C). For grain dehydration,grain moisture content decreased nonlinearly with ongoing ontogeny, and could be divided into two phases: rapid decline during fast grain filling and slow dehydration later (Fig. S3D). The averaged grain moisture of the red-cob hybrids was lower (P <0.001)than the whites at 53, 63, and 73 DAP, i.e., 0, 10 and 20 DARH.No significant difference was found at 14 and 28 DAP (Fig. S3D).Thus,we identified a tight connection between cob color and grain dehydration.
Given similar field conditions (temperature and humidity),grain dehydration may in general be driven by two main factors:physiological dehydration, which results from grain filling during maturation,and ecological dehydration,which results mainly from microenvironmental influences caused by plant morphological characteristics such as plant height, ear length and width, and foliar coverage [8,10,25]. The first is the essential cause of grain dehydration and the second influences the former via changes in environmental conditions. Grain dehydration and grain filling are synchronous during grain maturation.In agreement with the similar grain moisture at 14 and 28 DAP, the RPGR was slightly but insignificantly higher for reds than for whites(Fig. S3E). Both vegetative and reproductive phases were shorter for the red-cob hybrids, leading to an approximate average of 2–3 days earlier of maturity in the two seasons (Fig. S4A). The earlier transition from vegetative to reproductive stage in reds may promote grain dehydration. Plant height during grain filling was consistently greater,but ear diameter and LAI consistently lower, in red-cob hybrids than in whites (Fig. S4B–D). These traits may be beneficial for air flow and light incidence below the canopy, ensuring photosynthetic production and promoting grain water loss to the air[26].The lower ear height of red-cob hybrids may also increase lodging resistance [27]. We propose that the slightly higher RPGR,shorter vegetative phase, and in particular the more favorable plant morphological characteristics lead to faster grain dehydration for red-cob hybrids than for whites. Studies [10,11] have shown that husk traits can affect grain dehydration, while the differences between red- and white-cob hybrids deserve further study.
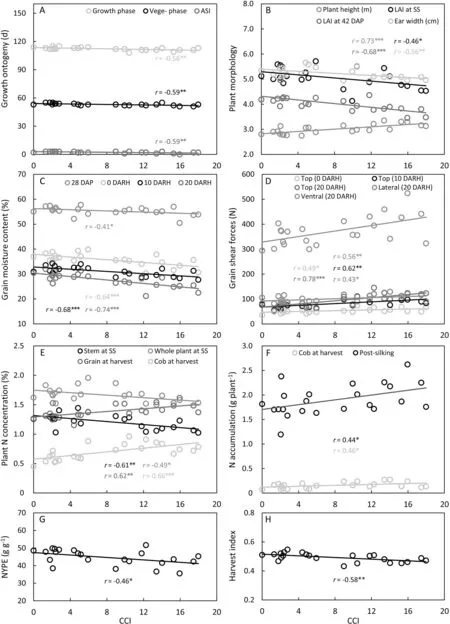
Fig.1. Correlation(n=23)between CCI and growth ontogeny(A),plant morphology(B),grain moisture content(C),grain shear force(D),plant N%(E),plant N accumulation(F),NYPE(G),and harvest index(H).ASI,anthesis and silking interval;CCI,cob color index;DAP,days after pollination;DARH,days after regular harvest;LAI,leaf area index;N%, N concentration based on dry weight; NYPE, N yield production efficiency; SS, silking stage.

Fig.2. Cob color information collected for common maize hybrids released in China.(A)Dynamic changes of cob color from 1991 to 2017(n=549).Shading indicates hybrid number collected in each period.(B)The proportion of each cob color among recently released grain harvest-mechanization hybrids(n=32).Data were collected from China Seed Association (http://www.seedchina.com.cn) and Seed Business (http://www.chinaseed114.com/seed/sspz/). Detailed information for the hybrids is presented in Table S4.
3.2. Establishment of CCI as a quantitative method to evaluate cob color and its correlation with agronomic traits
The binary classification of cob color is simple but rough.If cob color could be measured with greater objectivity and precision,more could be learned about its correlation with agronomic traits and underlying mechanisms. CCI values gradually increased from white to deep red(Fig.S5).The CCI of red-cob hybrids ranged from 8.97 to 17.98, while the CCI of whites varied from 0 to 5.95(Fig. S5). This finding indicated that CCI could quantify and differentiate cob color well. Among the parameters measured, CCI was significantly correlated with agronomic traits involved in growth ontogeny (growth phase, ASI, and vegetative phase), plant architecture(plant height,LAI at SS and 42 DAP,ear width),grain dehydration (grain moisture at 28 DAP and 0, 10, and 20 DARH), grain crushing(top shear force at 0,10,and 20 DARH;lateral and ventral shear force at 20 DARH),plant N(stem/whole plant N%at SS,grain and cob N% at harvest, cob Naccuat harvest and post-silking Naccu,and NYPE)and harvest index(Fig.1).The highest correlation coefficients were found for ‘‘CCI-grain moisture content” and ‘‘CCIgrain shear forces” (Fig. 1C, D). Thus, CCI was established as an effective method for quantification of cob color.
3.3. Examination of common hybrids released by China in the past confirmed the artificial selection of red cob
Xie et al. [15] used elite lines to show that cob color has been subjected to artificial selection for improvement in the temperate maize zone. It is not well known how the frequency of red cob has changed in commercial maize hybrids over the years. In 549 common hybrids released in China from 1991 to 2017, the frequency of red cob increased from approximately 50%to more than 80%,including 65%red,9%pink(light red)and 8%purple(dark red)(Fig,2A; Table S3).Recently released hybrids bred for mechanized grain harvesting were also collected (Table S3). The proportion of red cob was nearly 94%, and only one hybrid had a white cob(3.1%, n = 32, Fig. 2B). This finding strongly suggests a close relationship between cob color and the characters needed for mechanical grain-harvesting. Superior grain dehydration and grain hardness may be, at least partly, the driving forces for artificial selection (Fig. 1). Other reasons could include higher mold resistance and lower toxin accumulation for red-cob hybrids.
3.4. Cob color was also correlated with plant N
N absorption,distribution,and remobilization are necessary for plant development and grain protein deposition [16,28]. Plant N concentration was lower in vegetative tissues but higher in grain and cob for red-cob than for white-cob hybrids (Fig. S6A). This character may favor improvement of grain protein quality. However, only N accumulation in the cob reached significance(Fig. S6B). N deposition in ears depends on post-silking uptake and remobilization of pre-silking accumulation. Under the experimental conditions,post-silking N uptake(reproductive N accumulation), was consistently higher in red-cob than in white-cob hybrids in both seasons, and the 14% difference in reproductive N accumulation in 2017 reached significance (Fig. S6C). As to N remobilization,NRE was similar between the two types of hybrids(Fig. S6D). These results suggest that the higher N concentrations in grain and cob in red-cob hybrids may have been due mainly to N absorption and distribution rather than to remobilization during grain filling.The NHI was generally higher but the NYPE lower for the red-cob hybrids in both seasons (Fig. S6D, E). These results suggest the potential effects of selection for red cob on plant N metabolism.
4. Conclusions
We validated artificial selection for red cob using Chinese maize hybrids. We proposed that accelerated grain dehydration could motivation such selection. We developed a method to quantify cob color using the CCI, which could be used for future selection of cultivars for grain-mechanized harvesting. The potential effect of artificial selection for red-cob cultivars on plant N utilization invites further study.
CRediT authorship contribution statement
Xiao-Gui Liang:Conceptualization,Investigation,Methodology,Software,Writing–original draft,Writing–review&editing.Zhen Gao:Investigation, Data curation, Formal analysis.Si Shen:Software, Visualization.Xue Zhao:Investigation.Li Zhang:Investigation.Shun-Li Zhou:Supervision, Methodology, Project administration, Writing – original draft, Writing – review &editing.
Declaration of competing interest
Authors declare that there are no conflicts of interest.
Acknowledgments
This work was supported by the National Key Research and Development Program of China (2016YFD0300301) and the China Agriculture Research System of MOF and MARA, the Education Department Funds (190233) and the Natural Science Funds of Jiangxi Province (20202BABL215004).
Appendix A. Supplementary data
Supplementary data for this article can be found online at https://doi.org/10.1016/j.cj.2021.11.003.

- The Crop Journal的其它文章
- Brief Guide for Authors
- Effects of sgRNA length and number on gene editing efficiency and predicted mutations generated in rice
- Imbalance between nitrogen and potassium fertilization influences potassium deficiency symptoms in winter oilseed rape(Brassica napus L.)leaves
- ZmWRKY104 positively regulates salt tolerance by modulating ZmSOD4 expression in maize
- A novel genomic prediction method combining randomized Haseman-Elston regression with a modified algorithm for Proven and Young for large genomic data
- Identification of genes involved in the formation of soluble dietary fiber in winter rye grain and their expression in cultivars with different viscosities of wholemeal water extract