
金佛山方竹开花特性及花器官发育特征1)
2022-03-28 02:33杨蒙张玉丁雨龙姚文静林树燕
东北林业大学学报 2022年1期
杨蒙 张玉 丁雨龙 姚文静 林树燕
(南京林业大学,南京,210037)
竹类植物是非常重要的森林资源,属于禾本科竹亚科植物,主要分布于热带、亚热带和暖温带地区,具有生长快、分布广、用途多等特点,具有较高的生态和经济价值,被誉为“绿色的金矿”。世界上竹类植物约1 500多种,中国竹子资源有750多种[1]。虽然我国林业产业资源总体上表现为森林资源缺乏,但却拥有世界上最为丰富的竹林资源[2],因此,有效进行竹林资源培育以及扶持竹产业发展可为我国林业产业发展开辟新的道路。
竹类植物有其独特的开花生物学特性,开花周期长,有些数十年甚至上百年开一次花。部分竹类植物一生只开一次花,开花后竹秆整株死亡,部分竹种结实。由于竹亚科植物开花周期长、花期难以预测,使得竹子开花这一正常的现象成为研究的热点和难点,与竹子开花的相关研究已经成为竹类植物研究工作者们感兴趣的方向之一。目前,不仅对紫竹、雷竹、孝顺竹、淡竹、笔竹、翠竹、黄条金刚竹等竹类植物进行了开花生物学特性方面的观察和研究[3-9],也对竹类植物解剖结构方面进行了系统的研究。黄坚钦等[10]对雷竹的小孢子发生和雄配子体形成的研究发现,雷竹的花药壁在整个发育过程中始终无药室内壁层的形成和分化,而表皮层的发育起到药室内壁的作用;李洁[11]对孝顺竹的胚胎学进行了研究,并分析了导致孝顺竹结实率低的原因;林树燕等[12]在对‘霞早’绿竹花形态特征及花药发育的组织学观察中明确了‘霞早’绿竹小穗中各小花的发育进程、开花特性以及不结实的主要原因。
金佛山方竹(Chimonobambusautilis(Keng) Keng)属于禾本科竹亚科寒竹属,是一种优良的笋用以及观赏用竹种,复轴混生型小径竹,节间圆筒形或略为四棱形。金佛山方竹竹笋味美可口,竹材可用作造纸原料[13]。近年来,金佛山方竹在贵州省遵义市桐梓县陆续出现零星开花现象,这为研究金佛山方竹开花特性以及花器官发育提供了机遇。本文对金佛山方竹的开花物候、花器构造、大小孢子及雌雄配子体发育等进行系统而详细的研究,为了解金佛山方竹开花特性和花器官发育等奠定基础,也为其他竹类植物开花的相关研究提供参考。
1 研究区概况
以贵州省遵义市桐梓县狮溪镇箐坝村开花的金佛山方竹竹林为研究区,该地气候类型属于中亚热带高原季风湿润型气候,年均气温14.6 ℃,最冷月份气温-5 ℃,最热月份气温24.5 ℃;海拔高,山峰多,气候垂直变化明显,年平均降水量1 038.8 mm;夏季降水量最多,冬季降水量最少,呈冬干夏湿特点。
2 研究方法
金佛山方竹开花竹调查:在开花的金佛山方竹林中选定3个5 m×5 m样方,调查并记录分析竹林内开花与未开花竹株的株数,年龄组成,胸径。
金佛山方竹花序形态特征:2019年3月2日在处于盛花期的竹林中,随机选取3株,观察并记录金佛山方竹小穗、小花、外稃、内稃、浆片、雌雄蕊等各部分形态特征,每个部分观测样品50个,并记录小花开花动态,在LEICA DFC295体式显微镜下进行形态观察并拍照。
金佛山方竹花器官发育特征:2018年8月花芽分化至2019年3月盛花期期间,标记3株开花竹株,按花芽及小穗发育程度每周定期采集开花竹子的小穗,取材后立即放入体积分数为70%的 FAA固定液中固定待用。取出固定好的小穗,转移至体积分数为70%乙醇中,体视镜下解剖小花,再将小花的内外稃片依情况剥离,按照小花的雄蕊长度分级保存在盛有10 mL体积分数为70%乙醇溶液的玻璃瓶中,常规石蜡法制片,切片厚度为8 μm,番红-固绿着色,中性树脂胶封片。在LEICA DM2500显微镜下拍照观察,Photoshop进行显微图像处理。
3 结果与分析
3.1 开花竹林林相
由图1可知,金佛山方竹开花初期竹株为有叶开花型,开花过程跨越两个年度,开花竹株在2018年8月中下旬开始进入花芽分化期,9月上旬分化出小穗,9月中旬分化出小花,此时小穗长为10~20 mm,花药长2~4 mm;10月中旬小穗长20~30 mm,花药长5~6 mm;11月中旬小穗长30~40 mm时,花药长7~8 mm;12月中旬小穗长50~60 mm,花药长为9~10 mm,2019年2月中旬进入开花期,盛花期时间较短,依天气情况,晴天时约为7 d,阴天时约为10 d,盛花期后竹株进入结实期,2019年4月中下旬果实陆续成熟,成熟后即脱落。

a/b.花芽分化期(2018年8月);c/d.竹林及花枝(2018年9月);e/f.竹林及花枝(2018年10月);g/h.盛花期竹林及花枝(2019年3月);i/j.竹林及花枝(2019年3月底);k/l.开花竹林及果实(2019年4月15日)。
3.2 开花竹林中开花与未开花竹株的分布
由表1可知,通过调查开花金佛山方竹林地竹株的生长情况,发现林地内新生竹(1年生和2年生)的株数很少(约占总数的12%),多数为3~4年生竹株(约占总量的88%),由于竹林进入开花期,表现出衰退的林相,老竹开花,出笋减少,导致1~2年生的竹株减少。从平均胸径来看,开花竹株的胸径普遍小于未开花竹的胸径,未开花竹虽然胸径差异不显著,其1~2年生竹株的胸径也小于3~4年生的竹株,说明金佛山方竹一旦开花,林分成衰退趋势,竹株数量和秆茎粗度均受到影响。

表1 开花与未开花竹的生长参数
3.3 方竹开花的生物学性状
由图2可知,金佛山方竹的花序属于混合花序(图2a),小穗是花序的基本组成单位,小穗分为顶生小穗和侧生小穗两种,小穗形状为披针形、紫色、体扁、无毛、有柄,有退化的苞片,每个小穗着生5~11朵小花,小花在小穗轴上互生,顶端小穗不发育。金佛山方竹的成熟小穗有柄,柄长5.0~20.5 mm。花为颖花,雌雄同花,小花有外稃1枚,内稃1枚,均为紫色,外稃环抱内稃,内稃包裹浆片以及雄雌蕊,外稃披针形、紫色、膜质、无毛;内稃披针形、紫色、有2脊;每个小花含3个雄蕊,花药紫色,散粉时顶端开裂,丁着药,花丝白色,雄蕊长9.0~10.0 mm;每朵小花有浆片3枚,尖端紫色,底端透明,浆片大小不等,环绕雌雄蕊,肺形,浆片长3.0~4.0 mm,宽1.0~3.0 mm,较小的一枚包裹在内稃内,较大的两枚包裹在外稃内,开花时底端膨大;雌蕊由子房、花柱、柱头3部分组成,子房椭圆形,花柱长0.4~1.0 mm,柱头羽毛状、白色、长2.0~5.0 mm、3裂;果实浆果状颖果。

a.金佛山方竹的花枝;b/c.小穗;d.小穗轴;e.开放的小花;f/g.雌蕊和雄蕊;h.外稃;i.外稃脉;j.内稃;k.浆片;l.果实。
3.4 方竹花药的发育过程
金佛山方竹的花在一天中都可以开花,金佛山方竹小花开放,首先内外稃微张,紫色花药初露,伴随内外稃张大,浆片膨大,花丝伸长,花药先是半露,再是全露,伸出内外稃外,最后花药完全下垂,开裂并散粉。花药一般从内外稃微张,到完全露出稃片外需2~6 h,从花药完全露出到花药散粉需3~5 h。金佛山方竹的小花含有3枚花药,花药的发育过程要经过孢原细胞、造孢细胞、小孢子母细胞、小孢子母细胞减数分裂这几个发育时期形成小孢子,小孢子进一步发育形成雄配子体。因此,花药的发育过程分为小孢子发生时期和雄配子体发育这两个阶段(见图3)。
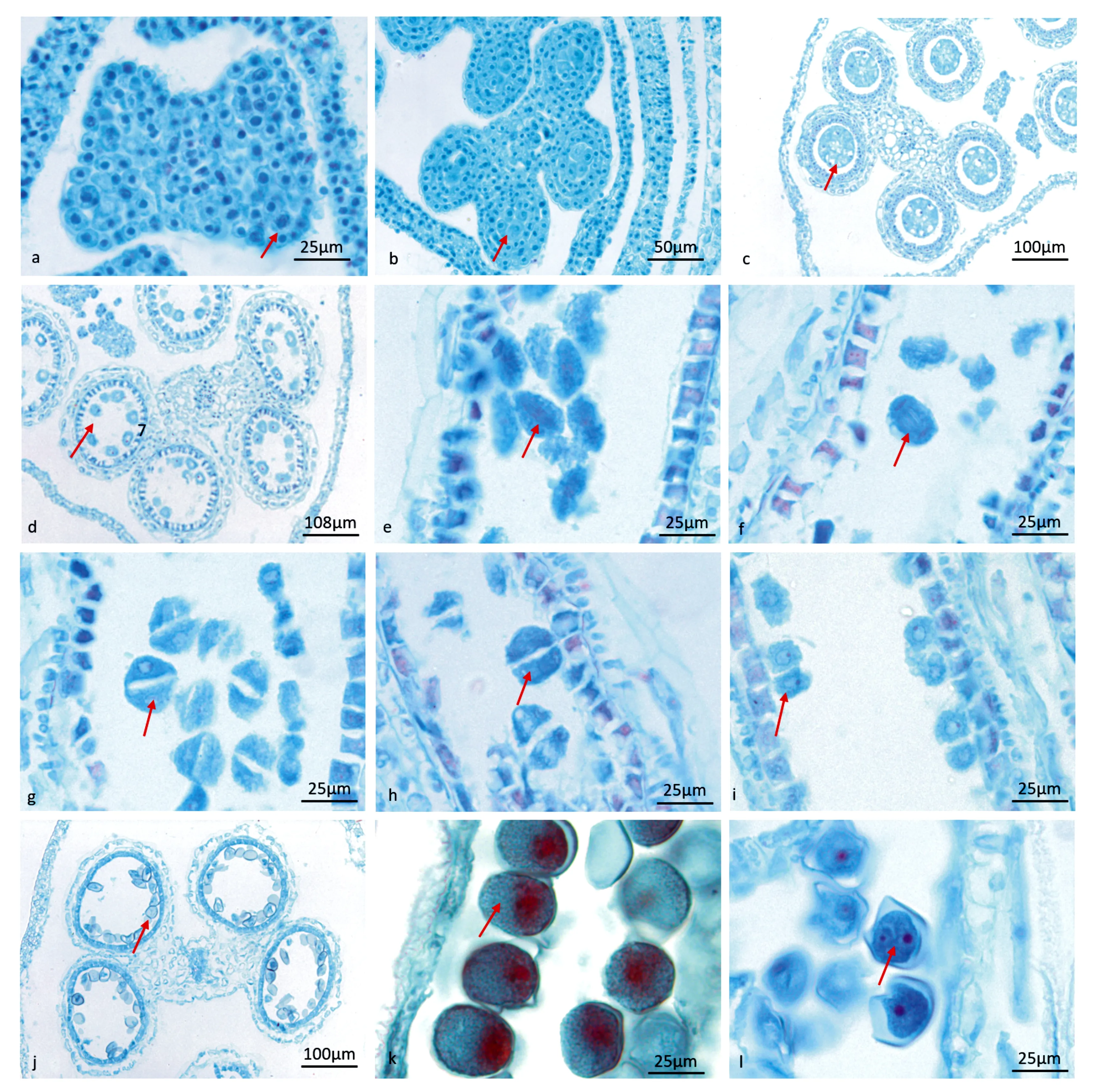
a.初生造孢细胞;b.次生造孢细胞;c.小孢子母细胞;d.小孢子减数分裂;e.减数分裂中期Ⅰ;f.减数分裂后期Ⅰ;g,减数分裂前期Ⅱ;h.四分体;i.四分体刚释放出的小孢子; j、k.单核靠边期;l.二核花粉粒。
小孢子发生过程:花药最初由表皮细胞和分生细胞发育而来,位于四个角隅处的细胞先分裂,在这四个角隅处表皮层下方分化出孢原细胞,孢原细胞体积较大,具有极强的分裂能力,此时花药长度为0~1 mm,经过一次平周分裂,多次垂周分裂产生初生壁细胞和初生造孢细胞;当花药长度为1~2 mm时,由初生造孢细胞再次分裂形成次生造孢细胞;当花药长度为3 mm时,次生造孢细胞再发育形成核大,且无明显液泡的小孢子母细胞;当花药长度为4 mm时,小孢子母细胞发育变为椭圆,随即进入减数分裂时期,这一减数分裂过程与大多数被子植物的分裂过程一致,经过第一次减数分裂(前期、中期、后期、末期)后,母细胞产生分割壁,形成两个子细胞(即二分体),两个子细胞进入第二次减数分裂阶段,经过第二次减数分裂(前期、中期、后期、末期)四个核之间形成细胞壁(即四分体),四分体左右对称,小孢子母细胞胞质分裂的方式为连续性。小孢子母细胞减数分裂的过程非常短暂,在同一药室同一时间小孢子的核在减数分裂过程中发育进度也会出现不一致的情况,发现小孢子第二次减数分裂时会同时出现中期Ⅱ和末期Ⅱ。
雄配子体发育过程:金佛山方竹小孢子四分体维持的时间较短,四分体胼胝质壁发生溶解,小孢子随即分开,成为游离的单核花粉粒,此时花药长度为5 mm;刚释放的小孢子细胞质浓厚,无液泡,细胞壁薄,细胞核位于中央,为单核居中期;小孢子进一步发育,细胞壁加厚,体积增大,细胞质液泡化,随着中央大液泡的扩大,细胞核被推向一侧,为单核靠边期,此时花药长度为6~7 mm,液泡随后消失,细胞质充满细胞;当花药长度为7~8 mm时细胞核进行第一次有丝分裂,形成营养核和生殖核,胞质不均等分裂,在两核之间形成细胞板,小孢子分成两个细胞,靠近花粉壁的是生殖细胞,位于细胞中央的为营养细胞,称为2-细胞型花粉粒,少数生殖细胞有丝分裂,形成两个精细胞,由两个精细胞和一个营养细胞构成雄配子体(3-细胞型花粉);当花药长度为9~10 mm时,花粉粒成熟散粉。
在金佛山方竹花药的发育过程中,发现也有少量小孢子在收缩期时,无法正常液泡化,小孢子核不能进行正常的分裂过程,导致花粉败育。
3.5 方竹花药壁发育
由图4可知,金佛山方竹花药壁发育属于单子叶型,孢原细胞进行第一次平周分裂,向外形成初生周缘层,初生周缘层的细胞进行垂周分裂形成两层次生周缘层,处于外层的细胞直接发育为药室内壁,在里层的细胞进行一次平周分裂形成中层和绒毡层,花药处于小孢子母细胞时期时,花药壁分化形成四层花药壁细胞。花药壁由初生周缘层细胞发育而来,在次生造孢细胞时期,花药壁的发育尚未明显分化,到小孢子母细胞后期,花药壁才分化完全,花药壁的结构从外向内依次是表皮、药室内壁、中层、绒毡层。金佛山方竹在花药发育的整个过程中,处于最外层的表皮细胞一直只有一层,在横切面上,大小为9.4~12.5 μm。表皮细胞只进行垂周分裂增加细胞的数目以适应花粉囊的扩大生长,通常花药横切面上表皮细胞有26~35个。药室内壁的细胞呈现出长方矩形,在花药的发育过程中,它的体积不断增大,在小孢子母细胞时期,药室内壁细胞不断发生径向方向的伸长,在横切面上,径向长度为20.3~25.0 μm。通常花药横切面上药室内壁细胞有15~20个;中层细胞存在花药壁的时间很短,在小孢子母细胞时,它的形状为矩形,在横切面上,径向长为7.1~11.9 μm;当小孢子母细胞减数分裂时期,中层细胞变扁,呈现狭长形,并且渐渐解体;在成熟花粉粒时期,中层细胞已经消失,处于花药壁最内层的绒毡层起初为单核细胞;在小孢子母细胞时期,绒毡层细胞排列紧密,为矩形细胞,具有浓厚的细胞质以及较大的细胞核,在横切面上,径向长为4.8~7.1 μm;在小孢子母细胞进入减数分裂时期时,绒毡层细胞核分裂旺盛,细胞内大多数为两核;当单核小孢子形成后,绒毡层细胞出现消解现象,细胞内容物不断减少,细胞之间间隙变大,细胞界限模糊;随着小孢子继续发育至后期,绒毡层已经基本消解,当成熟花粉粒形成时,绒毡层细胞完全消失不见。
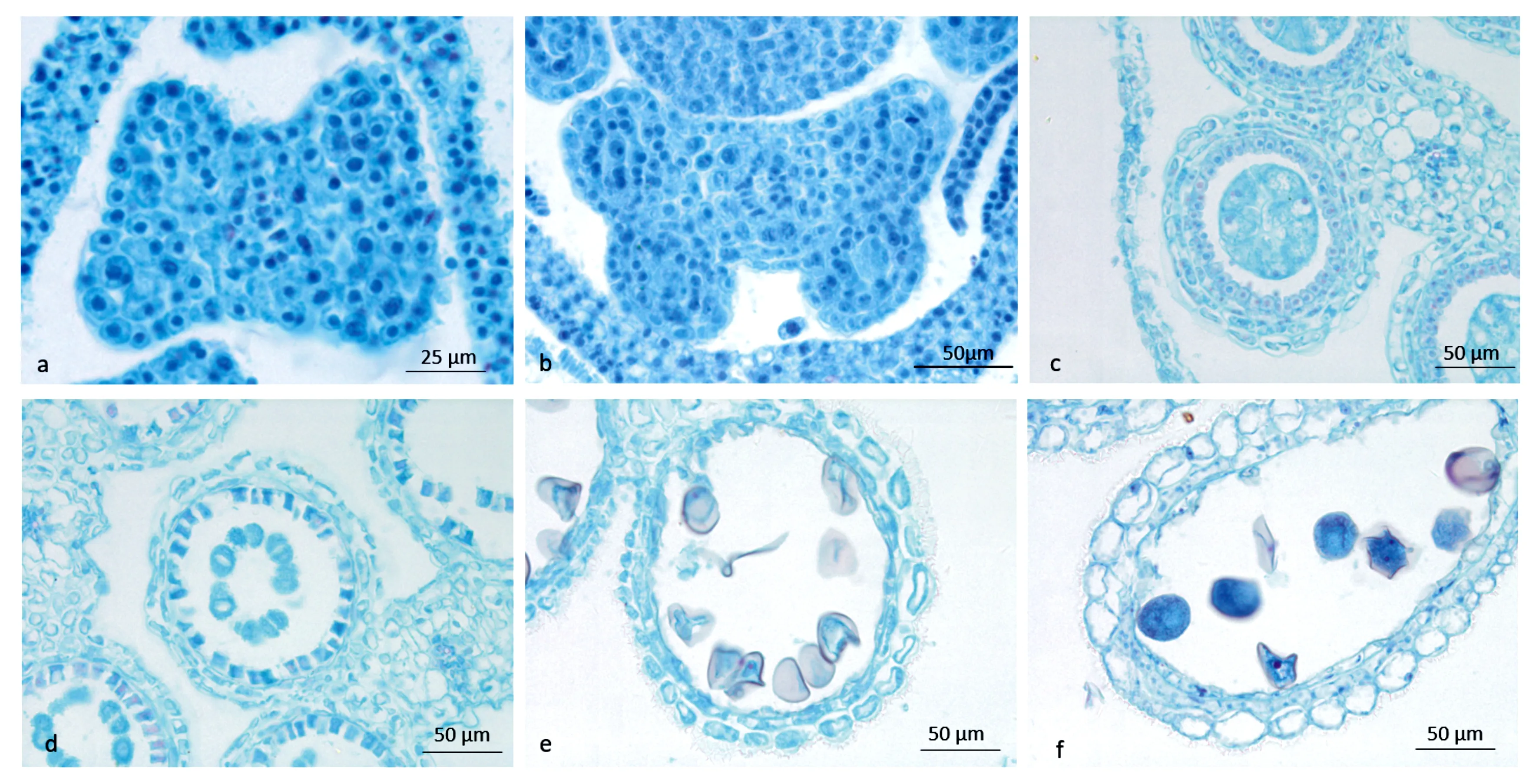
a. 初生造孢时期的花药壁;b. 次生造孢时期的花药壁;c. 小孢子母细胞时期花药壁;d. 二分体时期花药壁;e. 单核花粉粒时期花药壁;f. 成熟花粉粒时期花药壁。
3.6 大孢子发生及雌配子体发育
由图5可知,金佛山方竹的子房为一室单子房,柱头羽毛状两裂,在子房早期发育时,内室中央有环状突起,称为胚珠原基。内外珠被开始启动时,靠近珠心先端的表皮下第一层细胞分化出孢原细胞,孢原细胞体积大,细胞质浓厚,细胞核显著,内外珠被细胞不断向上生长,孢原细胞体积增大,发育成大孢子母细胞,胚珠开始倒转;大孢子母细胞进入第一次减数分裂时期,纵向排列分裂成二分体,第二次减数分裂形成纵向排列的四分体,胚珠倒转180°,形成倒生胚珠,四分体中位于合点端的功能大孢子进一步发育,其它三个大孢子退化消失;位于合点端的功能大孢子成熟后,细胞内的液泡不断增大,发育成单核胚囊,进行第一次有丝分裂,形成二核胚囊,两个细胞核分别位于两端各自进行两次有丝分裂,分别形成四核胚囊和八核胚囊,液泡不断扩大,胚囊体积纵向扩大。八核胚囊早期,位于两端的细胞核各有一个移向胚囊中央,组成一个具有两个核的中央细胞,中央细胞高度液泡化,位于合点端的3个细胞发育为3个反足细胞,位于珠孔端的3个细胞其中一个发育为卵细胞,另外两个发育为助细胞,合称为卵器,整个胚囊构成雌配子体。
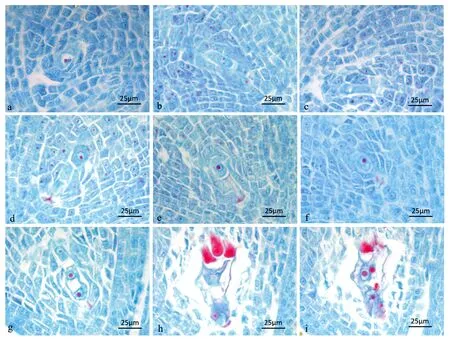
a.由孢原细胞发育而来的大孢子母细胞;b.二分体;c.四分体;d/f. 靠近合点端的功能大孢子;g.二核胚囊;h. 3个反足细胞;i. 成熟胚囊。
3.7 雌蕊、雄蕊发育对应关系
由表2可知,同一花药长度的小花雌蕊和雄蕊的发育进程基本保持一致,小花早期发育时,当花药的孢原细胞分化出造孢细胞,子房内的珠心表皮分化出孢原细胞;当小花花药长2~3 mm时,花药内的造孢细胞分化出小孢子母细胞,雌蕊的子房内大孢子母细胞发育形成;当花药长度为4 mm,子房和花药内的大小孢子母细胞均处于减数分裂时期,减数分裂的过程非常短暂;当花药长度为5 mm时,花药内为刚释放的小孢子,子房内为单核胚囊;当花药长度为6~7 mm时,花药内为单核靠边期,子房内为二核胚囊;当花药长度为7~8 mm时,花药内为小孢子经有丝分裂产生2或3细胞花粉粒,子房内大孢子形成四核胚囊,四核胚囊再进行分裂形成八核胚囊;当花药长度为9~10 mm时,花药内小孢子发育为成熟花粉粒散粉,子房内大孢子也发育为具有多个反足细胞的成熟胚囊。因此,金佛山方竹的花属于雌雄蕊同熟的类型。
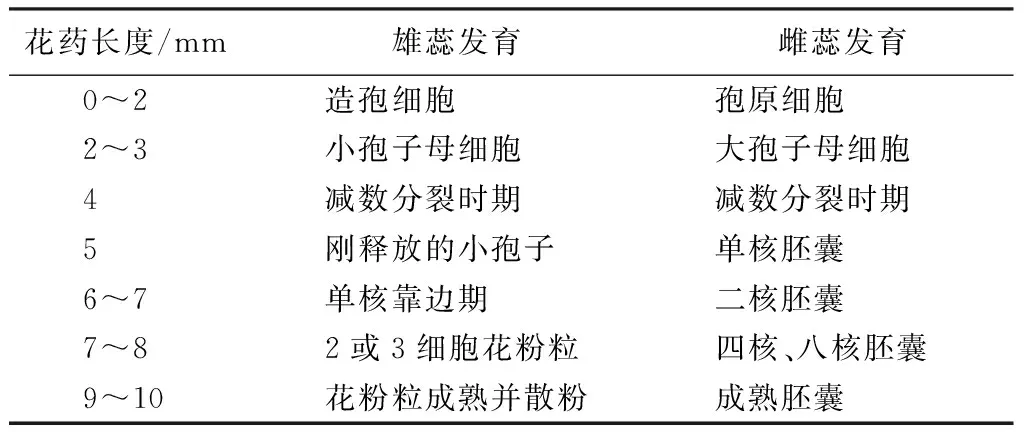
表2 金佛山方竹花药长度与雌雄蕊发育进程的对应关系
4 讨论
4.1 开花竹林林相
根据历史文献记载,金佛山方竹曾经历两次开花,1935—1938年间分布在贵州省桐梓县柏芷山、箐坝大山和毗邻的重庆金佛山林区的金佛山方竹全部开花死亡[14],1984年有性种实更新的金佛山方竹林在桐梓县境内又一次大面积开花死亡[15]。通过对贵州省桐梓县的金佛山方竹开花竹林林相的观察发现,金佛山方竹目前属于零星开花。竹类植物盛花期的时间因竹种不同表现出较大差异,金佛山方竹的盛花期在3月份,而异叶苦竹的盛花期在4月上旬、孝顺竹的盛花期为5月份、福建茶秆竹的盛花期为4—5月份、鹅毛竹的盛花期在3月中旬、月月竹的盛花期在3—4月份,酸竹属的一个竹种在10—11月份开花[16]。金佛山方竹开花竹林年龄多为3年生和4年生,为有叶开花型竹子,花芽分化到开花结果期历经两个年度,在当年8月份形成花芽,9月分化出小穗和小花,次年2—3月进入开花期,盛花期在3月份,4—5月份结果。
4.2 竹类的花序特点
竹类植物以小穗为基本单位构成花序,与普通植物学定义不同,竹类植物的无限花序与有限花序的区别在于小穗基部有无芽[17]。无限花序基部保留生长点,可以不断产生休眠芽,生长出新的假小穗。有限花序基部没有休眠芽,即一次形成小穗一次开花。无限花序基部的分生组织具有分裂和分化的能力,能够不断形成新的小花,最早形成的花着生于花轴的基部,其开花方向是朝顶部的方向;有限花序的顶端分生组织只分化出一朵小花,花序轴生长有限,花着生在花轴的顶端,小花从上至下依次开放[18]。竹类植物的花序单次发生、续次发生是划分超族的依据[19],这在竹类植物的分类学上具有重要的意义。林树燕等[20]通过研究霞早绿竹、黄条金刚竹以及翠竹的花序,对竹类植物的花序类型进行了修订,认为竹子的花序是以小穗为单位,竹类植物的花序修订为混合花序,通过查阅以往文献和观察试验发现,不同的竹种在同一小穗上的开花先后顺序不统一,一种是小穗基部先开放,一种是小穗顶部的小花先开放,也有位于小穗中部的小花先开放,开花的先后虽然不同,但是小穗的发育统一。金佛山方竹的花序由顶生小穗和侧生小穗组成,属于混合花序。金佛山方竹同一花序轴上,顶端小穗上的小花先开放,基部小穗上的小花后开放,开放顺序为从花序轴顶端至花序轴基部;在同一小穗上,小穗基部的小花先开放,顺次向顶端开放。
4.3 雄蕊发育特征
被子植物通过双受精作用来产生新的个体,有性生殖的方式很大程度维持了物种遗传物质的稳定,同时,基因重组丰富了种群的变异类型。竹类植物大多数通过无性繁殖的方式产生新个体,而通过有性生殖方式繁殖的新个体有利于种群的繁衍发展。金佛山方竹的花药壁的结构从外向内四层结构,依次是表皮、药室内壁、中层、绒毡层,花药壁发育为单子叶型,绒毡层为腺质型,小孢子母细胞进行减数分裂产生4个大小一致左右对称排列的四分体,胞质分裂为连续型,这与林树燕等[21-23]观察的月月竹、鹅毛竹、孝顺竹结果基本一致。而黄坚钦等[10]指出,雷竹的花药壁由4层细胞组成,从外到内依次为表皮、中层和绒毡层,没有药室内壁的存在,与本文的研究结果不同。
4.4 雌蕊发育特征
金佛山方竹由二心皮卷合发育而来,一室子房,胚珠为倒生胚珠,具有双珠被,薄珠心。金佛山方竹的胚囊发育类型与水稻、小麦等禾本科植物的一样,都是蓼型胚囊。金佛山方竹大孢子母细胞减数分裂产生4个线形排列的四分体,靠近珠孔端的3个退化消失,靠近合点端的一个细胞有作用,发育为功能大孢子。发育方式与翠竹[24]、异叶苦竹[25]的的发育方式一致,而鹅毛竹[25]、毛竹[26]、孝顺竹[11]在大孢子母细胞四分体时期位于珠孔端的单胞发育为功能大孢子。
金佛山方竹的成熟胚囊为7胞8核,即3个反足细胞,具两核的中央细胞,两个助细胞、及位于助细胞之间的卵细胞。3个反足细胞具有继续分裂增殖的能力,形成反足细胞团,与鹅毛竹、异叶苦竹、孝顺竹、淡竹、翠竹等[6,11,24-25]形成反足细胞团的结果一致。在禾本科植物中,反足细胞的数目也会增多,体积会增大。小麦反足细胞约为26个[27],多胚水稻ApⅢ的反足细胞大多数是增殖形成4~20多个细胞,其功能一般与胚囊营养的吸入有关[28-29],主要为8核胚囊的发育提供营养,早中期8核胚囊发育的时期表现得比较明显,而在后期8核胚囊发育的时期却表现得不是很明显。
猜你喜欢
医学信息(2022年17期)2022-10-11
传染病信息(2022年3期)2022-07-15
健康护理(2022年3期)2022-05-26
当代水产(2022年1期)2022-04-26
癌症进展(2021年9期)2021-07-20
小学生学习指导(当代教科研)(2021年6期)2021-05-23
家庭医药(2020年5期)2020-05-14
阅读(低年级)(2019年8期)2019-10-10
创新作文(1-2年级)(2017年5期)2017-12-07
科学大众(中学)(2015年9期)2015-10-12
