
象草全基因组bHLH 转录因子家族鉴定及表达分析
2022-03-26 02:27高莉娟张正社文裕宗西方闫启卢丽燕易显凤张吉宇
草业学报 2022年3期
高莉娟,张正社,文裕,宗西方,闫启,卢丽燕,易显凤,张吉宇*
(1. 兰州大学草地农业生态系统国家重点实验室,兰州大学农业农村部草牧业创新重点实验室,兰州大学草地农业科技学院,甘肃 兰州 730020;2. 广西农业职业技术学院,广西 南宁 530007;3. 广西畜牧研究所,广西 南宁 530001)
象草(Cenchrus purpureus)为禾本科蒺藜草属多年生的C4草本植物,起源于非洲,目前在全世界热带和亚热带地区均有栽培种植[1-2]。20 世纪40 年代由印度、缅甸等低纬度地区引入我国[3]。现已在江西、福建、湖南、台湾等地区广泛栽培种植[4-5]。象草茎叶中粗蛋白含量可达10.8%,营养价值高,生物量大,抗逆性强,再生能力强,已成为世界上公认的高产优质牧草[6-9]。近年来,随着生活水平的提高,人们愈加重视食物种类和安全问题,在肉类的选择上更倾向于草食动物产品,因此,进一步提高牧草产量显得至关重要[10]。
bHLH(basic-helix-loop-helix)类转录因子广泛存在于真核生物中,该类蛋白因具有bHLH 结构域而得名[11-12]。该结构域大约由60 个氨基酸残基构成,按照其功能可分为碱性区域和HLH 区域[13]。碱性区域由15~20 个氨基酸组成,分布在多肽链N 端,含有大量碱性氨基酸,能与DNA 结合,其保守氨基酸可识别E-box(5'-CANNTG-3')[14]。HLH 区域大约由40 个氨基酸组成,分布在C 端,依赖疏水氨基酸的互作可以促使蛋白质形成同源或异源二聚体[15-16],这些二聚体可以与靶基因启动子的不同部分相结合,在真核生物转录调控中起着十分重要的作用[17-19]。
bHLH 转录因子在植物形态发育过程中发挥着重要作用。据报道,水稻(Oryza sativa)的稀穗突变体(LAX PANICLE,LAX)是属于bHLH家族的转录因子,是控制植株顶端分生组织的主要调节因子[20]。研究表明,bHLH 转录因子在花色素生物合成过程中起着重要的调控作用。在水稻中,OSB1 和OSB2 能够参与调控花青素的生物合成[21]。随后在矮牵牛(Petunia hybrida)中获得的AN1基因被证明可以参与花青素的合成[22]。bHLH 类转录因子参与植物对逆境响应的研究备受关注,该家族通过激活相关基因的表达,进而提高了植物对干旱、低温、缺素等恶劣环境的耐受性[23]。SPATULA基因首次在拟南芥(Arabidopsis thaliana)中被发现,是一个多功能基因,主要调节拟南芥花形态建成和角果发育等多种发育过程的bHLH 蛋白。此外,SPATULA也参与控制茎尖分生组织的周围区以及叶片、花瓣、柱头和根的特定组织的发育[24-25]。随着分子生物学的快速发展,bHLH家族的多样性已被揭示,且植物bHLH 转录因子已经在拟南芥[26]、水稻[27]、杨树(Populussp.)[28]、玉米(Zea mays)[14]以及其他物种中得到鉴定,但象草bHLH家族成员分析还未见报道。
本研究通过全基因组数据库搜索鉴定,共鉴定出229 个CpbHLH基因,并对其进行了系统发育、染色体分布、保守基序生物信息学分析,并结合本课题组前期转录组测序(未发表数据),分析了CpbHLHs基因在赤霉素(gibberellin,GA3)和多效唑(paclobutrazol,PAC)处理之后的表达情况。这将为深入研究象草CpbHLH基因家族生物学功能提供科学依据,并有助于bHLH基因在其他植物物种中的功能研究。
1 材料与方法
1.1 材料
供试象草的种茎由广西壮族自治区畜牧研究所提供,于2019 年5 月在兰州大学榆中校区温室内种植,在种植后的30 d(节间发生最早期)[29],各选取27 株长势较为一致的象草,以喷雾的方式分别均匀喷施200 mg·L-1的赤霉素(GA3)和500 mg·L-1的多效唑(PAC),喷施蒸馏水作为对照(CK),处理时间为8:00 am,在整个喷施过程中,将相邻的植株用黑色塑料袋罩起来。在处理后的0,1,48 h 分别取各处理的茎尖,每个时间点至少取9 个单株,液氮速冻后-80 ℃保存备用。
1.2 象草bHLH 转录因子家族的鉴定
从PlnTFDB(3.0)(http://plntfdb.bio.uni-potsdam.de/v3.0/)下载得到水稻和拟南芥bHLH 蛋白序列。象草全基因组数据来自本课题组[30],编码序列(coding sequence,CDS)、蛋白质序列、基因序列均下载自国家生物信息中心(National Genomics Data Center,https://bigd.big.ac.cn/)。以水稻和拟南芥bHLH序列作为查询序列,利用BLAST 鉴定出象草基因组中的候选bHLH基因。然后,从PFAM 数据库(http://pfam.xfam.org/)中下载bHLH(PF00010)家族基因的隐马尔科夫模型(hidden markov model,HMM),最后利用Pfam,NCBI conserved Domains(http://www. ncbi.nlm.nih.gov/Structure/cdd/ wrpsb.cgi)和SMART(http://smart.embl-heidelberg.de/)数据库核对以确定bHLH 结构域(PF00010)的存在。
1.3 染色体定位分析
提取所有bHLH基因在象草基因组中的位置信息,利用在线工具MapGene2Choromosomev2(http:/mg2c.iask.in/mg2c_v2.0/)绘制CpbHLH家族基因的染色体定位图谱。
1.4 系统进化分析
利用MEGA 7 软件,使用邻接法(neighbor-joining,NJ)对象草和水稻的蛋白序列进行系统进化树的构建,其中校验参数Bootstrap=1000,其他参数均使用系统默认值。
1.5 基因结构、保守基序分析
利用MEME(http://meme-suite.org/tools/meme;最佳匹配长度为6~50,最大基序数为10)在线工具对提取到的bHLH 蛋白序列进行保守基序(motif)分析。使用基因结构显示系统GSDS(http://gsds.cbi.pku. edu.cn/index.php)绘制基因结构示意图。
1.6 象草bHLH 基因在赤霉素和多效唑处理后的表达分析
采用Trizol 试剂(北京天根生化科技有限公司)提取象草茎尖分生组织的总RNA,质量检测合格后,用于象草cDNA 文库的构建和Illumina 上机测序,该过程均由北京诺禾致源科技股份有限公司(http:// www.novogene.com/)完成。测序完成之后,利用获得的转录组数据(未发表)中bHLH基因在GA3和PAC 处理之后的茎尖分生组织的表达量(fragments per kilobase million,FPKM),绘制热图。
采用qRT-PCR 分析CpbHLH家族9 个基因在象草经GA3和PAC 处理之后的表达模式。引物序列见表1,选用象草CpEE1a为内参基因[31],PCR 反应程序为:95 ℃30 s;95 ℃5 s,60 ℃30 s,共40 个循环。每组试验设3 个生物学重复,应用2-ΔΔCT方法计算各基因的相对表达量[32]。

表1 实时定量PCR 引物序列Table 1 Primer sequences used in real-time quantitative PCR
1.7 象草bHLH 基因的功能分析
为了进一步明确CpbHLHs基因的功能及属性,本研究利用百迈客云在线数据分析平台(www.biocloud.net)对鉴定得到的229 个CpbHLHs基因进行GO 分类注释。
2 结果与分析
2.1 CpbHLH 转录因子家族成员的鉴定及其染色体定位分析
在象草基因组中总共鉴定得到229 个bHLH基因,根据它们在染色体上的位置分布,依次命名为CpbHLH001~CpbHLH229。
CpbHLH基因染色体定位(图1)分析表明,229 个CpbHLH基因分布在象草基因组上的14 条染色体上,其中B01 号染色体上分布最多,有23 个基因,占总的CpbHLH基因的10.0%。其次为B03 号和A07 号,分别占总个数的9.2%(21)、8.7%(20)。B07 号染色体上分布的基因最少,只有5 个基因,占总个数的2.2%。在象草A03 号、A04 号、A05 号、B06 号染色体的 底端,以及A02 号、A06 号、B04 号、B05 号 染色体顶端 发现了CpbHLH基因的基因簇。
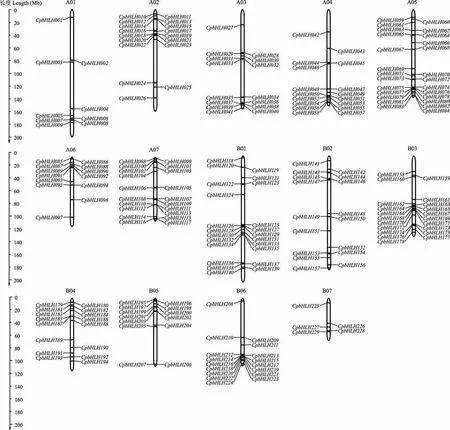
图1 象草CpbHLH 基因在染色体上的定位Fig.1 Chromosomal locations of C. purpureus bHLH gene family
2.2 CpbHLH 基因家族基因结构及保守基序分析
通过对CpbHLH 蛋白序列进行系统进化树分析发现,可以分为11 个亚族(图2a)。比较每个CpbHLH基因的内含子/外显子结构,对其结构进行分析(图2b)显示,CpbHLH基因中外显子数目为1(CpbHLH046,CpbHLH003等)~12(CpbHLH051)个不等,大多数基因(90%)含有1~6 个外显子,长度在300 kb 以上。内含子的数目差异较大,数目最多的是CpbHLH129和CpbHLH018,为11 个,有11 个CpbHLH基因不存在内含子。
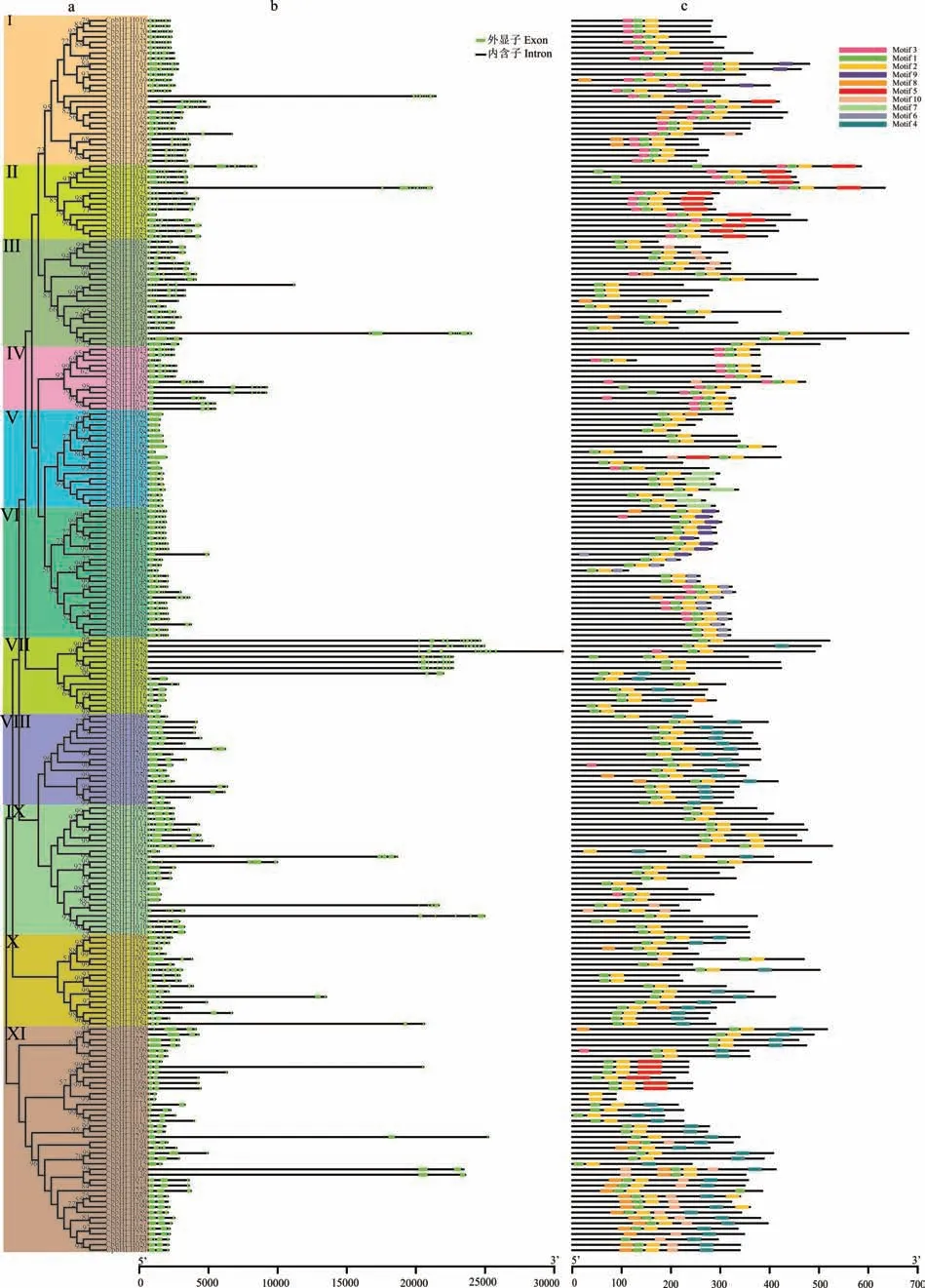
图2 象草CpbHLH 转录因子的保守基序分析Fig.2 The conserved motifs analysis of C. purpureus bHLH transcription factors
为了进一步揭示CpbHLH 蛋白在象草中的功能多样性,使用MEME 软件进行蛋白结构域预分析。结果分析表明:CpbHLH 蛋白结构较为保守,总共鉴定得到10 个比较保守的Motif(Motif1~10),其中Motif1 和Motif2 最为保守,是各组共有基序(图2c)。除共有基序外,各组基序还具有一定特异性,如Motif6 仅在Ⅵ亚组中存在,Motif9 只在I 和Ⅵ亚组中存在。
2.3 CpbHLH 基因家族系统进化分析
利用象草中229 条和水稻中的167 条bHLH 蛋白序列,进行象草和水稻中的系统进化关系分析。结果表明(图3),与水稻的22 个亚家族相比较,象草的229 个CpbHLHs成员分布在18 个亚家族中,其中两个物种都是C 亚类的成员数量最多。其次为B 亚族,含有24 个CpbHLH基因。G 亚族中CpbHLH成员数量较少,仅为2 个。其中Q、H、K、F 亚族中不包含CpbHLH基因。
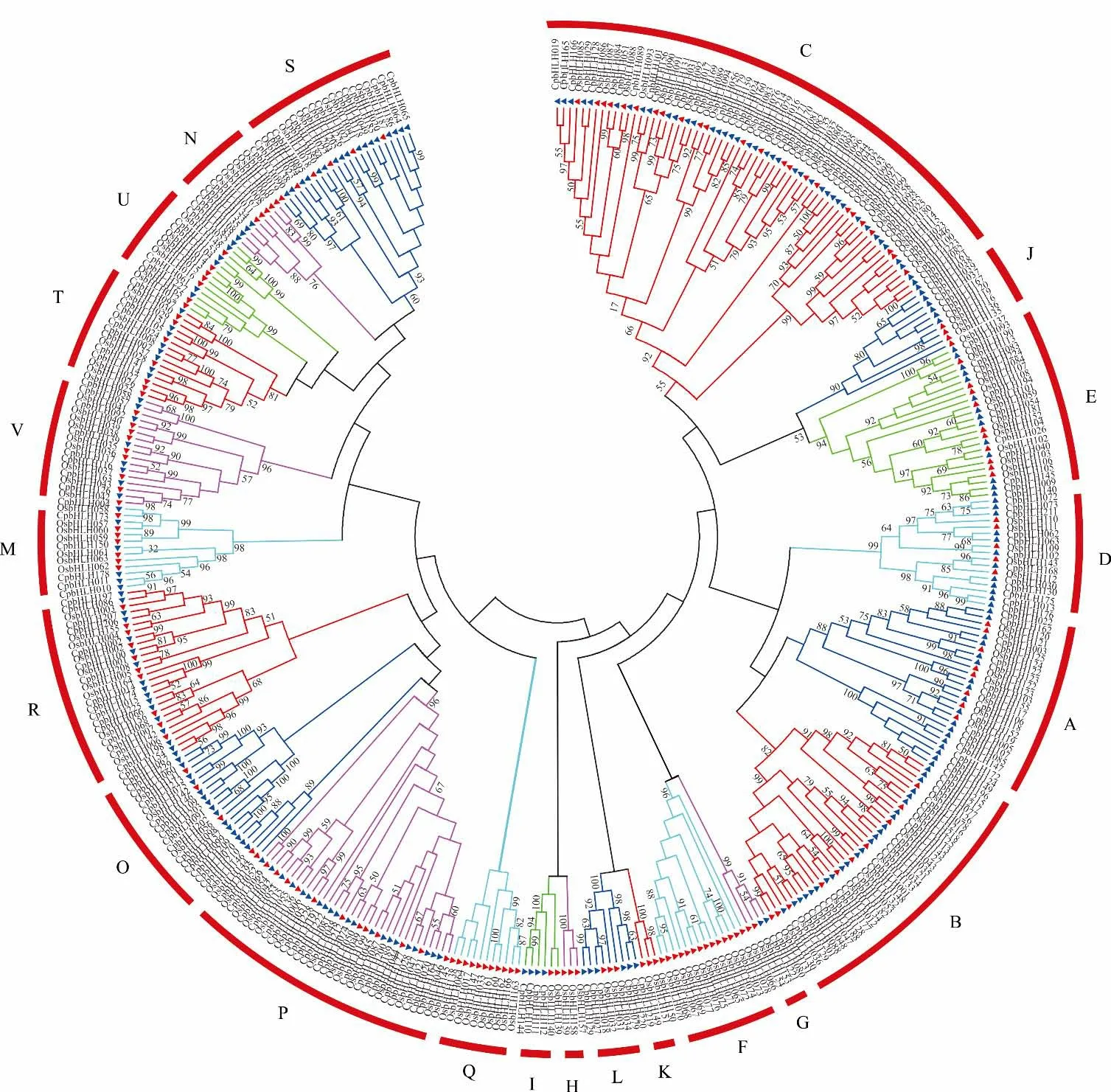
图3 象草与水稻bHLH 转录因子的系统进化分析Fig.3 Phylogenetic analysis of bHLH transcription factors in C. purpureus and O. sativa
2.4 CpbHLH 基因家族的表达分析
通过前期用赤霉素与多效唑处理不同时间后,对象草茎尖的转录组测序,得到RNA-seq 数据,分析得到CpbHLH基因在不同处理下的表达量(图4)。结果显示,赤霉素与多效唑处理后的不同时间点,bHLH基因均显示出高表达水平,因此可以推测在茎尖分生组织中表达的CpbHLH基因,其功能可能具有相似性。CpbHLH在GA3处理1 和48 h 以及PAC 处理1 和48 h 后分别有55、65 和51、56 个CpbHLH基因差异表达。
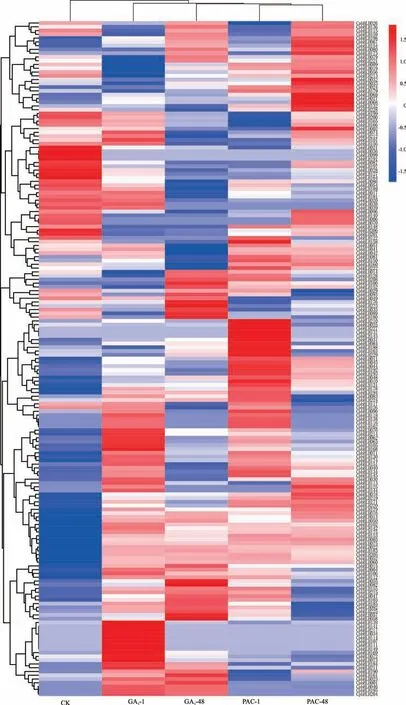
图4 象草CpbHLH 基因在GA3和PAC 处理1 和48 h 之后的表达分析Fig.4 Expression analysis of bHLH gene in C. purpureus after GA3 and PAC treatments for 1 and 48 hour
本研究结合转录组数据,选取了9 个在茎尖分生组织中相对表达量较高的CpbHLHs,进行赤霉素和多效唑处理之后的表达分析(图5)。研究结果显示,CpbHLH019、CpbHLH025、CpbHLH030、CpbHLH158在GA3处理之后的表达模式相似,在处理48 h 之后的表达量均高于1 h,且高于对照。CpbHLH013、CpbHLH055赤霉素处理1 h 之 后 的 表 达 量 高 于48 h,但CpbHLH002、CpbHLH017、CpbHLH156在GA3处 理 之 后 均 低 于 对 照。CpbHLH002、CpbHLH013、CpbHLH019在PAC 处 理48 h 后 表 达 量 高 于 处 理1 h 且 高 于 对 照。CpbHLH017、CpbHLH025、CpbHLH055、CpbHLH156在PAC 处理1 h 后的表达量均高于处理48 h 和对照,仅CpbHLH158经PAC 处理之后表达量低于对照。
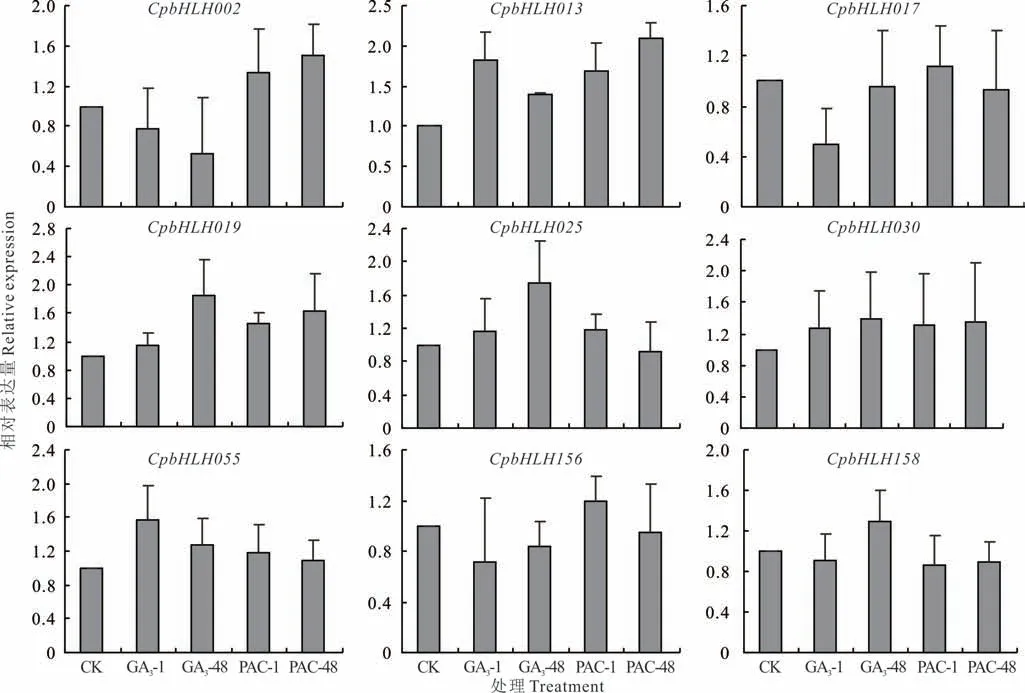
图5 CpbHLH 各成员在GA3和PAC 处理下的表达模式Fig.5 Expression pattern of CpbHLH genes under GA3 and PAC treatment
2.5 CpbHLH 基因家族的GO 分类结果分析
通过GO 功能对229 个CpbHLHs基因进行分类,发现可以将这些基因划分成47 个功能组(图6)。根据功能组的不同,又分为生物过程(biological process,20 个亚类),细胞组分(cellular component,15 个亚类)和分子功能(molecular function,13 个亚类)3 大类,分别占41.67%、31.25%、27.08%。在生物过程中,主要的亚类是“细胞过程”(cellular process),“代谢过程”(metabolic process);在细胞组分中,“细胞”(cell),“细胞组分”(cell part)和“细胞器”(organelle)是主要的亚类;在分子功能类别中,“结合”(binding)和“核酸结合转录因子活性”(nucleic acid binding transcription factor activity)占比最多。
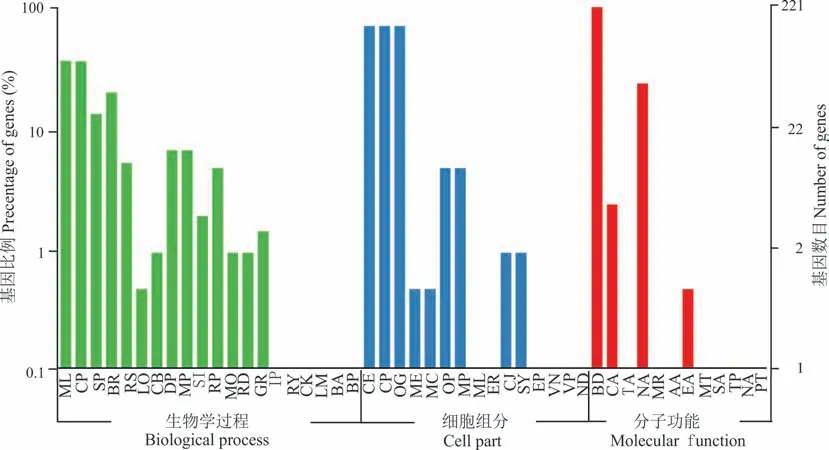
图6 象草bHLH 基因的GO 分类Fig.6 GO classification of CpbHLH genes
3 讨论
bHLH基因家族成员可以参与调控植物对逆境胁迫响应和信号转导等,在植物的生长发育过程中发挥着重要的作用[33-34]。到目前为止,关于模式植物:拟南芥以及水稻等bHLH 转录因子家族相关研究已经较为成熟,象草作为一种优质牧草,其推广种植具有重要意义,而其关于bHLH家族的研究却未有相关报道[35]。
本研究在象草基因组中总共鉴定得到229 个bHLH基因。根据染色体定位分析,大多数CpbHLHs基因被定位到象草染色体的顶端和底端,该结果与二穗短柄草(Brachypodium distachyon)中的研究结果相似[36],说明CpbHLHs基因进行了一定程度的收缩与扩张,这些基因更容易在染色体的顶端或底端被图位克隆[23]。
基因进化过程中结构变异起着重要作用,其中内含子会随着生物体内基因组的进化而进化,而进化主要体现在其种类和含量上[37]。Xu 等[38]研究发现,内含子、外显子会通过获得与丢失,插入或删除等方式来进化。本研究发现229 个CpbHLHs基因的外显子个数为1~12,内含子个数为0~11,说明CpbHLHs基因在进化过程中出现了外显子、内含子的插入或丢失,导致CpbHLHs基因中外显子与内含子的个数差异甚大,该现象在一定程度上影响CpbHLHs基因发挥其生物学功能。使用在线软件MEME 对象草229 条bHLH基因的保守基序进行分析,发现CpbHLHs基因存在10 个具有高度保守性的氨基酸基序,且同一亚家族中的大多数保守基序相似。表明每个亚家族中编码蛋白的功能是稳定的,几乎所有的CpbHLHs 蛋白均包含Motif1 和Motif2,且总是相邻,共同构成bHLH 结构域,所有这些bHLH 保守基序在同一亚家族的独特性及保守性也是对CpbHLHs基因家族的进化分类的佐证。同时推测,除bHLH 结构域外的其他保守基序也是每个亚家族发挥其相应功能的关键。总之,对bHLH的基因结构以及保守基序的分析,为bHLH基因的进化与分类提供了参考依据。它们在bHLH 中数量的变化也从侧面反映出bHLH 蛋白功能的分化,进一步证明bHLH 蛋白在植物生长发育的各个阶段所表现出的功能多样性。
系统进化树分析结果表明,CpbHLHs可以划分为18 个亚家族。与已报道的水稻、拟南芥及苹果(Malus pumila)[39]中的bHLH基因家族的结果比较,象草中发现的bHLH基因数量多于其他已报道的植物,这种情况可能与象草的四倍体进化事件有关[2,30]。在植物中,Pires 等[40]对源自藻类和陆生植物的500 多个bHLH 蛋白序列进行分析,将其分为26 个亚家族;白菜(Brassica campestris)中的203 个bHLH 蛋白可被划分为24 个亚家族[41];番茄(Solanum lycopersicum)中的152 个bHLH 蛋白可以划分为24 个亚家族[42]。与水稻相比,CpbHLHs家族成员并没有分布在所有亚类中,有4 个亚类中没有CpbHLH家族的成员,推测这可能是象草在进化的过程中发生了基因的丢失。此外,CpbHLH家族在G 亚类中的成员数量最少,仅为2 个,说明该亚类内的成员进化速度相对缓慢,推测这些基因在功能上相对比较保守,可能对象草的生长发育有着重要的调控作用[43]。象草在C 亚类中的bHLH成员数量最多,这与水稻、拟南芥[27]的聚类结果相似,该亚类转录因子的功能主要被预测为逆境响应和参与依赖脱落酸(abscisic acid,ABA)的信号传导[44]。本研究中,赤霉素处理后最大表达量基因CpbHLH055与多效唑处理1 h 后最大表达量基因CpbHLH076都为C 亚类,qRT-PCR 分析9 个随机挑选的CpbHLH基因在赤霉素与多效唑不同处理时间后的茎尖分生组织中的差异表达,这为bHLH 蛋白参与到赤霉素和多效唑诱导的生物学途径提供了证据。且其中CpbHLH017、CpbHLH025、CpbHLH030、CpbHLH055、CpbHLH156均为C 亚类。徐秀荣等[43]对 毛 竹(Phyllostachys edulis)bHLH 家 族 成 员 的C 亚 类 的4 个 基 因pebHLH072、pebHLH102、pebHLH112和pebHLH123在干旱和盐胁迫过程中的表达变化的研究结果支持了这一推测。说明该亚类内的成员可能对象草的生长发育有着非常重要的调控作用。
研究表明,水稻bHLH家族基因OsbHLH013、OsbHLH016、OsbHLH165参与调控花青素的合成;OsbHLH35可被干旱和盐胁迫诱导而上调表达,它可在水稻种子萌发阶段改变ABA 代谢基因的表达,降低ABA水平,提高种子的发芽率;基因OsbHLH001受冷胁迫诱导而上调表达,但不被ABA 或NaCl 所诱导;OsbHLH096参 与 缺 磷 胁 迫 反 应[45-47]。系 统 发 育 分 析 显 示(图3),CpbHLH家 族 基 因CpbHLH016、CpbHLH227与OsbHLH013、OsbHLH016、OsbHLH165聚集在一起,归属于U 亚族;CpbHLH139与OsbHLH35聚集在一起,归属于V 亚组,CpbHLH100与OsbHLH001聚集在一起,归属于R 亚族;CpbHLH193、CpbHLH045、CpbHLH044与OsbHLH096聚在一起,归属于C 亚族;通过同源基因功能相似性方法,预测到这些CpbHLH家族基因与水稻同源性基因具有相似功能。通过GO 分类发现,与分子功能和细胞组分相比,生物学过程的GO 分类数较多,共有20 个分类。说明CpbHLH 转录因子在参与多个生物学过程调控中发挥较为重要的作用,表明CpbHLH家族具有功能多样性,这些结果可为将来CpbHLH基因功能的进一步分析提供依据。
4 结论
本研究对CpbHLH基因家族进行分析,共鉴定筛选出229 个CpbHLH基因家族成员,并对其进行了生物信息学分析,发现各基因对赤霉素、多效唑处理之后的响应也不同。因此,bHLH基因在象草体内的调控机理还需要验证,进一步为象草的生长发育提供重要的信息。
猜你喜欢
安徽农业科学(2022年13期)2022-07-18
落叶果树(2022年3期)2022-06-07
科学之谜(2019年3期)2019-03-28
科学之谜(2018年8期)2018-09-29
现代农业科技(2016年20期)2016-12-20
中学生理科应试(2016年4期)2016-11-19
恋爱婚姻家庭·养生版(2016年9期)2016-09-07
湖北农业科学(2014年12期)2015-01-06
山西果树(2014年3期)2014-07-15
果农之友(2009年8期)2009-09-11
