
基于SSR 标记的河南省狗牙根遗传多样性及群体遗传结构分析
2022-03-26 02:27:32任雪锋邓亚博臧国长郑轶琦
草业学报 2022年3期
任雪锋,邓亚博,臧国长,郑轶琦
(河南科技大学园艺与植物保护学院,河南 洛阳 471000)
狗牙根(Cynodon dactylon)隶属禾本科狗牙根属,别名行仪芝,爬根草,百慕大草,是世界上暖季型草坪草的广布种之一,广泛应用于绿地建植,水土保持,运动场铺设,畜牧等[1-3]。狗牙根原产于非洲,在全世界广泛分布[4]。我国拥有丰富的狗牙根种质资源,主要分布在黄河流域及以南地区,另外在华北、新疆、河南、甘肃等地区都有分布[5]。前人应用分子标记技术对我国的狗牙根种质资源遗传多样性进行了大量研究,Li 等[6]对我国11 个省的95 份狗牙根利用ISSR 分子标记进行了遗传多样性研究,Zheng 等[7]对我国20 个省共157 份狗牙根利用SRAP 分子标记进行了遗传多样性及群体遗传结构的分析与评价,周少云[8]对我国华南地区的118 份狗牙根应用ISSR 分子标记进行了遗传多样性研究,凌瑶等[9]运用AFLP 与SRAP 标记对四川、重庆、贵州、西藏、云南以及非洲地区共59 份狗牙根的遗传多样性进行了分析,刘杰[10]运用ISSR 分子标记对采自安徽省的150 份狗牙根的遗传多样性进行了分析,李会彬等[11]运用SSR 分子标记对河北省的34 份狗牙根的遗传多样性进行了分析,李江华等[12]应用SRAP 分子标记对我国新疆地区的48 份狗牙根材料的遗传多样性进行了研究,以上研究均表明我国的狗牙根具有丰富的遗传多样性。
河南省地处中国中部,是南北气候带的过渡区域,植物资源丰富[13]。狗牙根在河南省分布广泛,主要分布于海拔1000 m 以下的浅山、丘陵、平原地区[14]。目前对河南省狗牙根种质资源的鉴定与评价工作较少。邓亚博等[15]基于河南省狗牙根的表型性状对遗传多样性进行了研究,结果表明河南省狗牙根具有丰富的表型遗传多样性,遗传变异主要来源于居群内,另外纬度和年降水量是影响狗牙根表型性状变异的主要因素。由于表型性状容易受外部环境的影响,因此有必要应用DNA 分子标记对河南省狗牙根的遗传多样性及群体遗传结构进行系统研究。本研究应用SSR 分子标记,对采集于河南省15 个狗牙根居群的遗传多样性和群体遗传结构进行分析和评价,以期为合理开发和利用河南省狗牙根种质资源,推动我国草坪草新品种的培育奠定理论基础。
1 材料与方法
1.1 材料及来源
在河南省选取15 个采样点,每个采样点采集一个居群,每个居群采集16~20 份样本,居群内各样本间采样间隔不小于10 m,每份样本采集生长良好,无病虫害的叶片,装入有硅胶的自封袋中进行保存。采样地点见图1,采样点概况见表1。
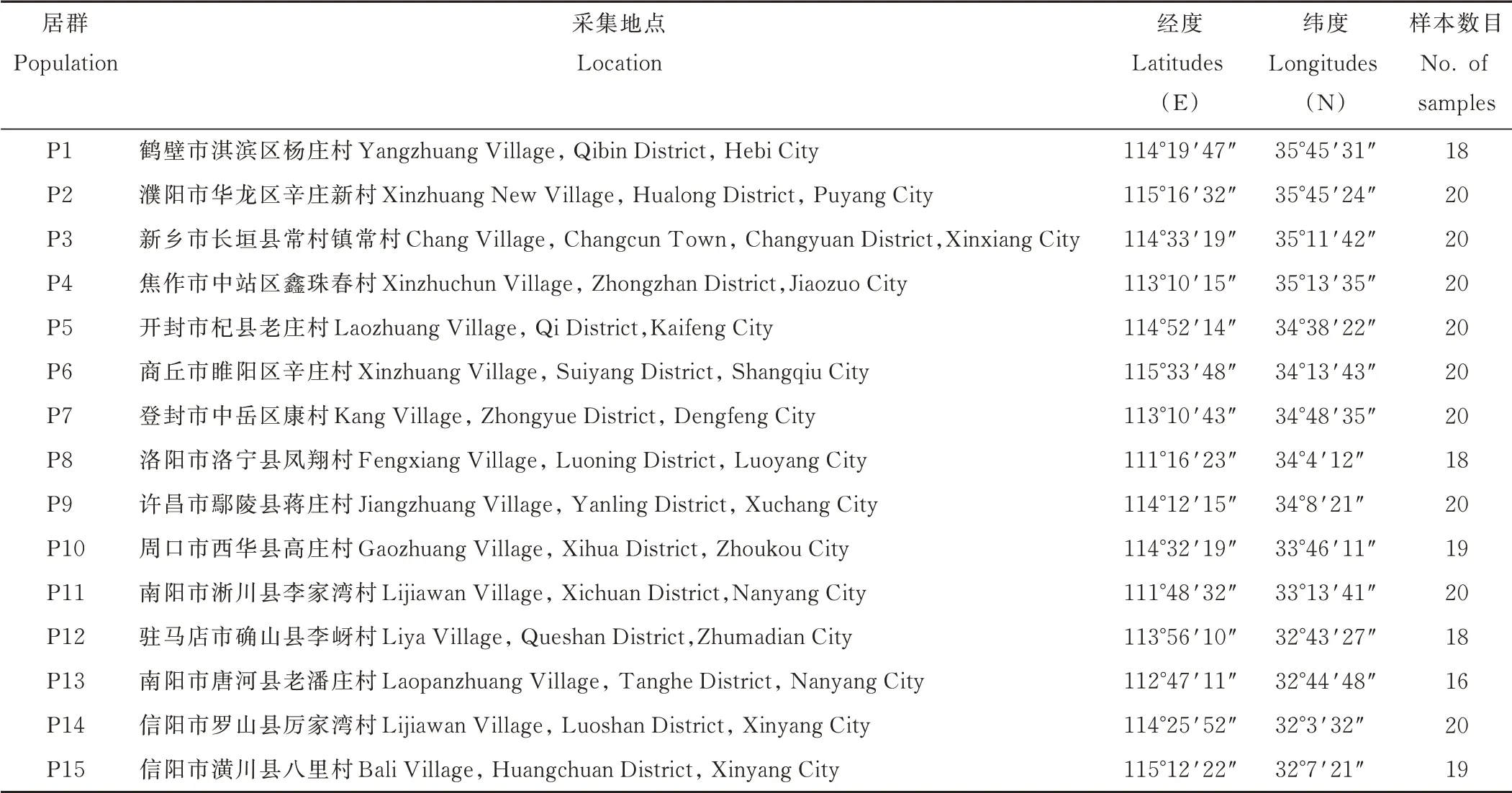
表1 狗牙根居群采集点概况Table 1 Survey of collecting sites of bermudagrass populations
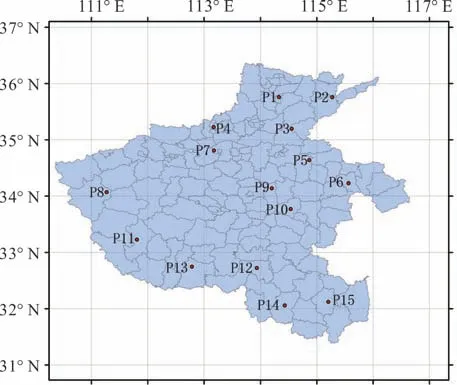
图1 狗牙根居群采集点分布Fig.1 The map of collection site for bermudagrass populations
1.2 试验方法
采用植物DNA 提取试剂盒(DP350-02,北京天根生化科技有限公司)提取DNA。对提取的DNA 质量用1%的琼脂糖凝胶电泳进行检测,对DNA 的浓度与纯度用紫外分光光度计(T6 新世纪,北京普析通用仪器有限责任公司)进行检测,将DNA 溶液稀释至50 ng·μL-1,放置-20 ℃保存备用。从本实验室前期筛选的狗牙根SSR 引物中,选择多态性高,重复性好的10 对引物(表2)进行分析,引物序列、扩增反应体系以及扩增程序参照Wang 等[16]的报道。

表2 本研究所采用的SSR 引物序列Table 2 Sequences of SSR primers used in the study
1.3 数据统计与分析
对扩增后的产物进行人工判读,对每一条带,按照有、无分别赋值1 和0,建立SSR 分子标记0/1 矩阵。使用POPGENE 1.32 软件[17]计算遗传多样性的相关参数,包括观测等位基因数、有效等位基因数、Nei’s 遗传多样性指数、Shannon’s 信息指数、多态性条带百分率、Nei’s 遗传距离、遗传一致度、居群间基因分化度(gene differentiation coefficient,Gst)、基因流(gene flow,Nm)等。将POPGENE 计算的居群遗传一致度导入NTSYSpc 2.1 软件[18],按照UPGMA 法进行聚类分析,并对狗牙根居群间地理距离与遗传距离进行Mantel 检验。用NTSYS-pc 2.1 软件计算供试材料的遗传距离矩阵,将计算结果导入MEGA 6.0[19]中采用UPGMA 法形成288 份狗牙根的树状聚类图。
使用软件Structure 2.3.4[20]分析供试材料的群体遗传结构,将群体数目预测值K设置为2~10,将开始时的MCMC(markov chain monte carlo)不作数迭代设置为10000 次,然后将不作数迭代后的MCMC 设置为100000次,重复运行5 次,将运行结果导入在线程序Structure harvester(http://taylor0. biology. ucla. edu/structureHarvester/)进行最佳群体数预测。根据各材料在各亚群中Q值是否大于0.7[21]来划分不同亚群。
2 结果与分析
2.1 SSR 扩增结果
应用SSR 引物对288 份狗牙根材料进行检测,扩增结果见表3。10 对SSR 引物共扩增出173 条条带,其中多态性条带共163 条,平均每对引物扩增出16.3 条多态性条带,引物CDCA77-78、CDCA747-748、CDE127-128、CDE375-376 扩增的多态性条带数最多,各有18 条,引物CDCA155-156 扩增的多态性条带数最少,有13 条。引物CDCA379-380 和CDE127-128 多态性百分率最高,达到100%,引物CDCA747-748 和CDE375-376 多态性条带百分率最低,为90.00%,平均多态性条带百分率为94.29%。Nei’s 基因多样性指数最高的是引物CDCA55-56(0.3850),最低的是引物ES295577(0.1187),平均Nei’s 基因多样性指数为0.2307。Shannon’s 遗传多样性信息指数最高的是引物CDCA55-56,达到0.5678,最低的是引物ES295577,为0.2016,平均Shannon’s 遗传多样性信息指数为0.3736。综上所述,所选引物均具有较高的多态性,对供试材料具有较高的分辨率。
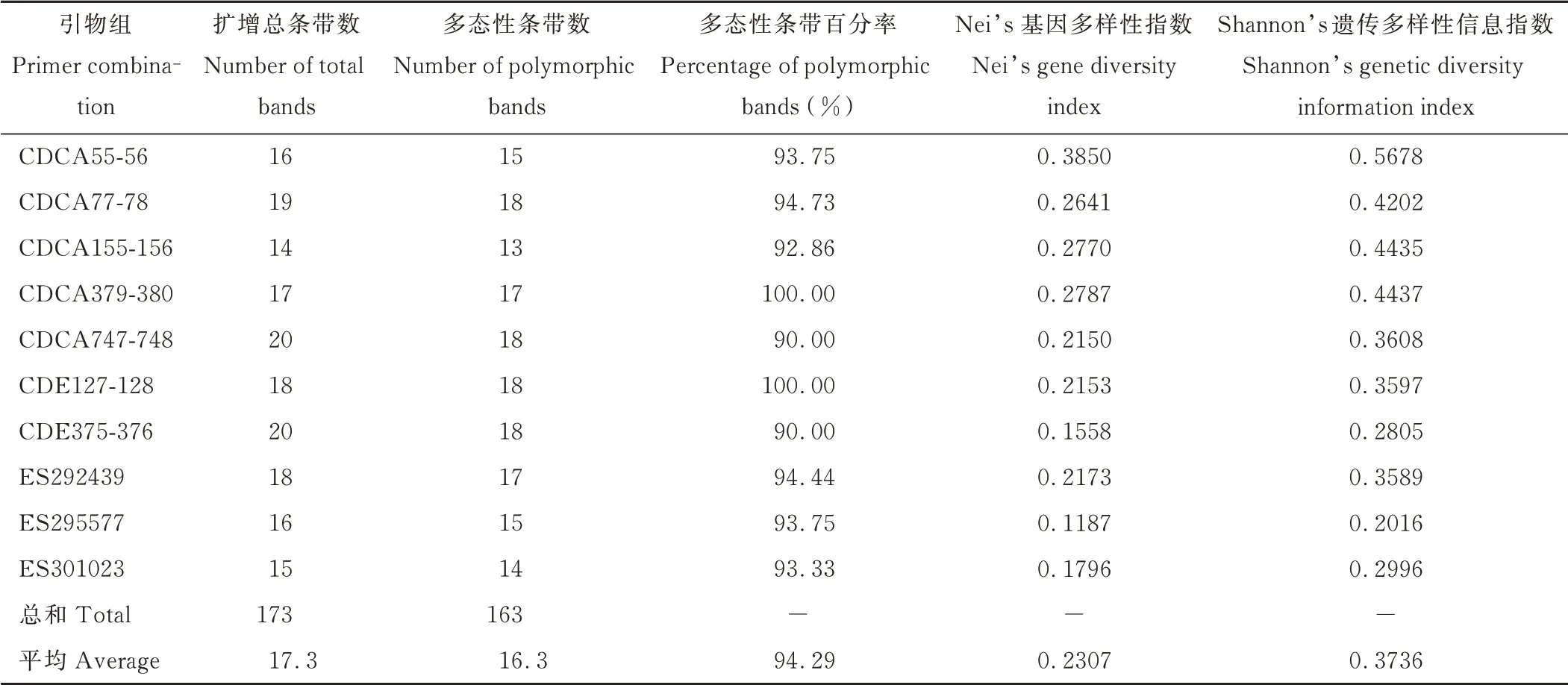
表3 引物扩增结果Table 3 Primers amplification results
2.2 遗传多样性分析
狗牙根居群遗传多样性分析结果见表4,观测等位基因数为1.4220~1.7572,最大的为P5 居群(开封市杞县老庄村),最小的为P1 居群(鹤壁市淇滨区杨庄村),平均为1.5437;有效等位基因数为1.1971~1.3555,最大的为P4 居群(焦作市中站区鑫珠春村),最小的为P1 居群,平均为1.2552;Nei’s 基因多样性指数为0.1186~0.2144,最大的为P4 居群,最小的为P1 居群,平均为0.1151;Shannon’s 遗传多样性信息指数为0.1830~0.3293,最大的为P4 居群,最小的为P1 居群,平均为0.2398;多态性条带百分率为42.20%~75.72%,最大的为P5 居群,最小的为P1 居群,平均为54.37%。在居群水平上,15 个狗牙根居群各参数差异较大,其中来自P4 居群的3 个参数值(有效等位基因数、Nei’s 基因多样性指数、Shannon’s 遗传多样性信息指数)均最大,P1 居群的5 个参数值最小,综合表明P4 居群的遗传多样性水平最高,P1 居群的遗传多样性水平最低。物种水平上的Nei’s 基因多样性指数、Shannon’s 遗传多样性信息指数、多态性条带百分率分别为0.2288、0.3717、100%,表明采集于河南省的狗牙根材料具有丰富的遗传多样性。
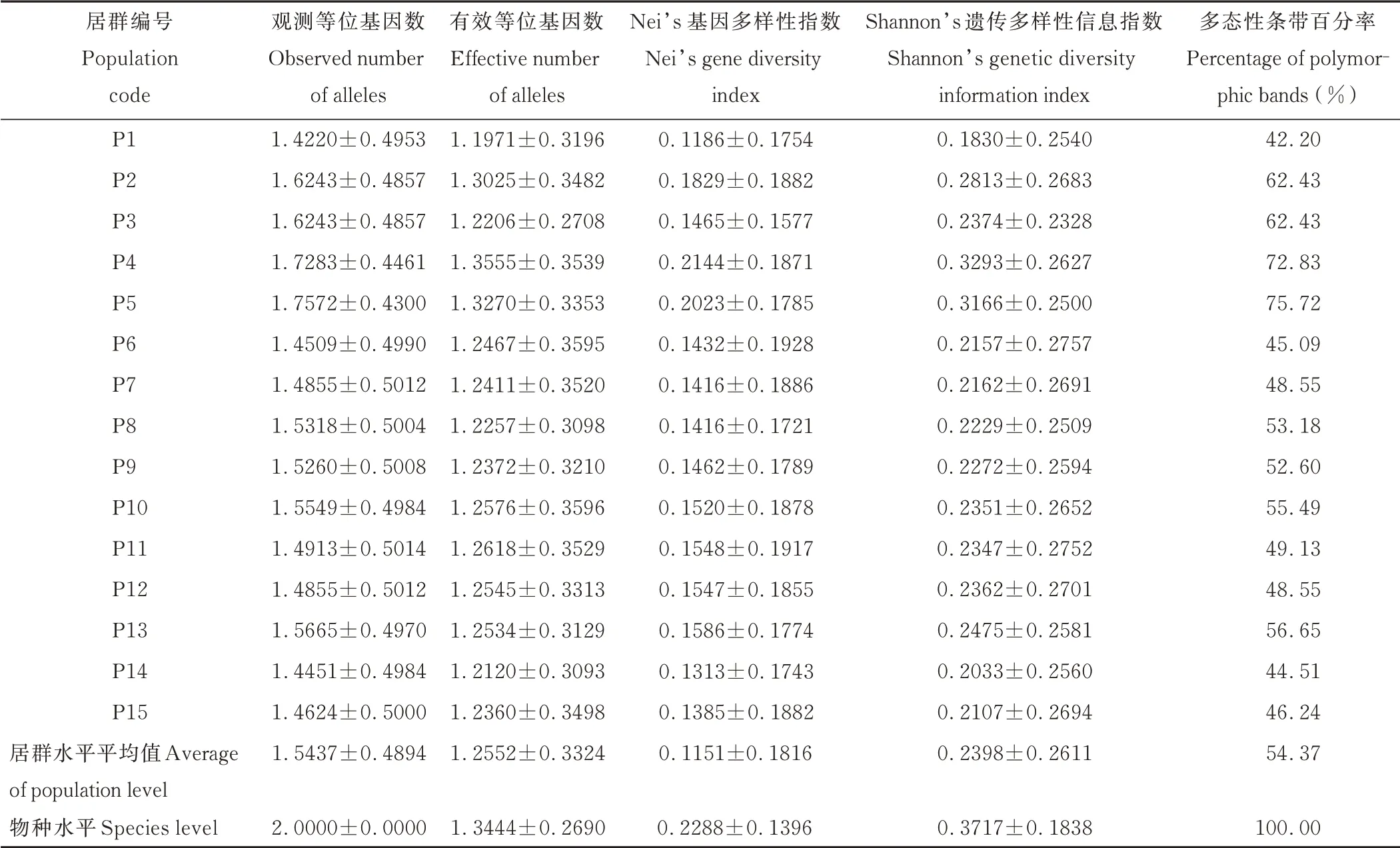
表4 河南省狗牙根居群的遗传多样性Table 4 Genetic diversity of bermudagrass populations in Henan Province(mean±SD)
2.3 居群遗传分化分析
基于POPGENE 1.32 计算居群间的遗传分化系数和基因流结果表明,15 个居群间的遗传分化系数(Gst)为0.3857,即发生在居群间的遗传变异达到38.57%,61.43%的遗传变异存在于居群内,表明河南省狗牙根居群的遗传变异主要发生于居群内。基因流(Nm)为0.7964,表明居群间存在一定程度的基因交流,但不频繁。
2.4 狗牙根居群的聚类分析
基于POPGENE 1.32 计算15 个居群间遗传一致度,其变化范围是0.746~0.964,平均为0.767,遗传一致度最小的是来自鹤壁市淇滨区杨庄村P1 居群和信阳市潢川县八里村P15 居群,为0.746,遗传一致度最大的是来自商丘市睢阳区辛庄村P6 居群和登封市中岳区康村P7 居群,为0.964。15 个居群UPGMA 聚类分析结果如图2 所示。在遗传一致度0.89 处,将15 个居群分为5 组,第1 组只包含来自鹤壁市淇滨区杨庄村的P1 居群,第2 组包含P2、P3、P4、P5、P13、P14、P15 共7 个居群,分别来自濮阳市华龙区辛庄新村、新乡市长垣县常村镇常村、焦作市中站区鑫珠春村、开封市杞县老庄村、南阳市唐河县老潘庄村、信阳市罗山县厉家湾村、信阳市潢川县八里村,第3组包含P8、P9、P10 共3 个居群,分别来自洛阳市洛宁县凤翔村、许昌市鄢陵县蒋庄村、周口市西华县高庄村,第4组包含P6、P7 两个居群,分别来自商丘市睢阳区辛庄村、登封市中岳区康村,第5 组包含P11、P12 两个居群,分别来自南阳市淅川县李家湾村、驻马店市确山县李岈村。聚类结果表明,居群没有完全按照地理来源进行聚类,如组成第2 组的居群包括豫东地区的P5、豫北地区的P2、P3、P4 居群和豫南地区的P13、P14、P15 居群。为进一步阐明地理距离和遗传距离之间的相关性,经Mantel 检验,相关系数r=-0.0950(P=0.2708>0.05),表明供试狗牙根居群的地理距离与遗传距离之间无相关性。
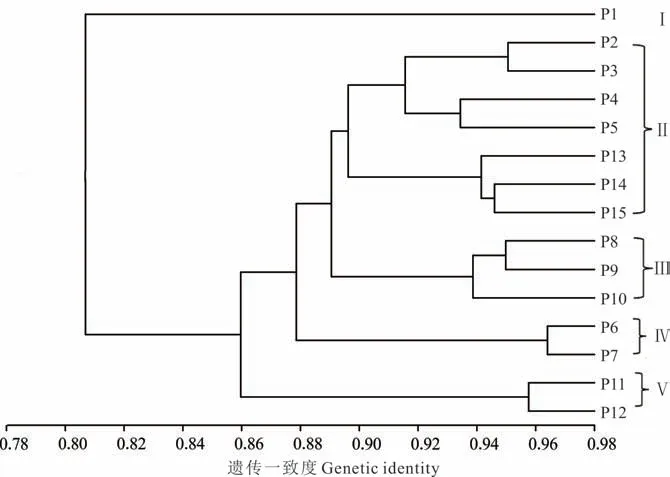
图2 15 个居群间UPGMA 聚类分析Fig.2 UPGMA cluster analysis among 15 populations
2.5 288 份狗牙根聚类分析
供试材料的遗传距离为0.0173~0.5205,平均遗传距离为0.3113,其中遗传距离最小的材料是来自P10 居群的184 号和186 号,遗传距离最大的是来自P1 居群的14 号和P15 居群的272 号。根据聚类结果将所有材料分成为3 组(图3)。Ⅰ组包含P1 居群、P2 居群、P6 居群、P7 居群、P8 居群、P9 居群的所有材料,P3 居群的10 份材料,P5 居群的2 份材料,P10 居群的16 份材料;Ⅱ组包含P11 居群、P12 居群、P13 居群、P14 居群、P15 居群的所有材料,P10 居群的3 份材料;Ⅲ组包含P4 居群的所有材料,P3 居群的10 份材料,P5 居群的18 份材料。结果表明,绝大多数来自同一居群的材料被分到同一组,并且在同一组中的材料并没有严格按照材料的地理来源进行聚类。
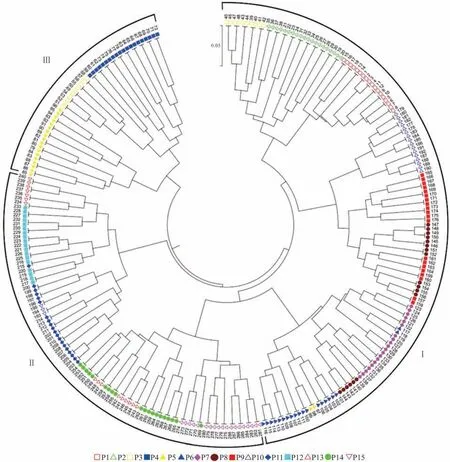
图3 288 份狗牙根材料UPGMA 聚类分析Fig.3 UPGMA cluster analysis of 288 bermudagrass materials
2.6 群体遗传结构分析
本研究采用Structure 软件对狗牙根材料进行群体遗传结构分析,K与ΔK的变化关系如图4,当K=2 时ΔK出现最大值,由此判断可将288 份狗牙根材料分为2 个亚群,K=2 时的群体遗传结构如图5 所示。分析各材料在不同亚群的Q值发现第1 个亚群(红色)主要来自P1、P2、P3、P6、P7、P8、P9、P10 居群,共8 个居群,包含144 份材料(50.0%),第2 个亚群(绿色)主要来自P11、P12、P13、P14、P15 居群,共5 个居群,包含96 份材料(33.3%),两大亚群的Q值≥0.7,属于单一型群体,其余48 份材料(16.7%)主要来自P3、P4、P5 居群,该亚群Q值<0.7,没有明确的亚群归属,形成了混合型群体,群体遗传结构分析与288 份狗牙根材料的聚类分析结果基本一致。
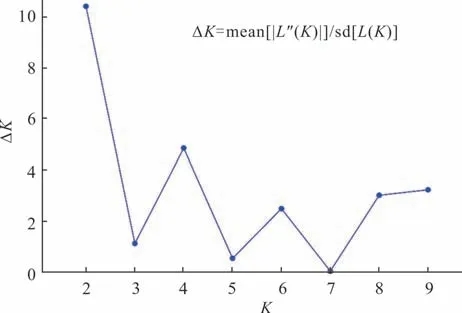
图4 基于Structure 软件分析的K 与ΔK 的变化趋势Fig.4 Trend of K and ΔK based on Structure analysis
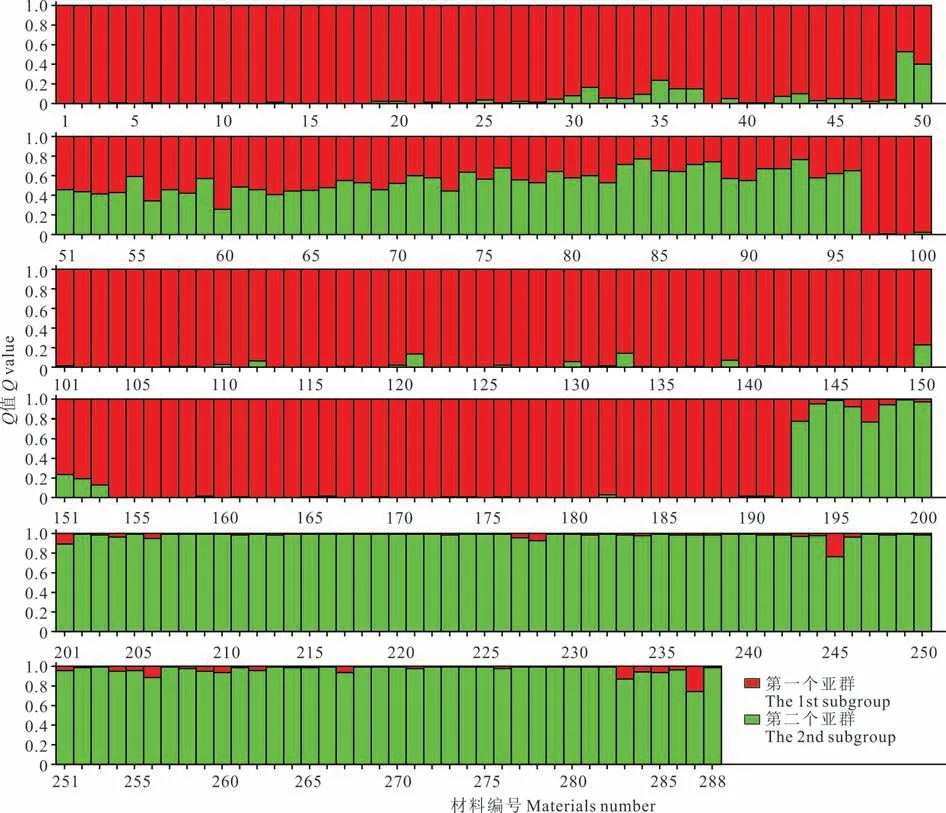
图5 基于SSR 标记的K=2 时288 份狗牙根群体遗传结构Fig.5 Genetic structure of 288 bermudagrass accessions at K=2 based on SSR markers
3 讨论
3.1 多态性分析
SSR 分子标记技术具有高多态性,高稳定性,操作简单等优点,可快速分析植物的遗传多样性。本试验对采自河南省15 个居群共288 份狗牙根材料,用10 对SSR 引物共扩增出163 条多态性条带,多态性条带百分率达到94.29%。张延辉等[22]对新疆地区的106 份野生狗牙根,运用20 对SSR 引物进行研究,多态性位点比率为87.86%;李会彬等[11]对河北省的34 份狗牙根材料,用21 对SSR 引物共扩增出185 条多态性条带,多态性条带比率占81.90%;凌瑶等[23]对我国西南地区5 个省份的55 份野生狗牙根材料进行SSR 与AFLP 联合分析,其中用18对SSR 引物扩增的产物,多态性条带比率为75.10%。与上述研究结果相比,本研究所选择的10 对SSR 引物能有效用于检测狗牙根的遗传多样性,同时表明河南省野生狗牙根具有丰富的遗传多样性。
3.2 遗传分化分析
河南省15 个居群的遗传分化系数(Gst)为0.3857,说明居群间的遗传分化占总体遗传变异的38.57%,居群内占61.43%,居群遗传分化主要发生于居群内。居群间的基因流(Nm)为0.7964,根据Govindaraju[24]对基因流(Nm)大小的划分,共分为3个等级,Nm≥1.00 为高水平,0.25≤Nm≤0.99 为中等水平,Nm≤0.25 为低水平,说明狗牙根居群间存在着中等水平的基因交流,即居群间存在着一定程度的基因交流,但不频繁,有限的基因流导致狗牙根居群间存在着一定程度的遗传分化。马涛[25]对湖南省野生狗牙根的遗传多样性研究中发现,居群间的遗传分化系数为0.4693,基因流为0.5075,指出居群间存在较高的遗传分化。凌瑶等[23]对西南地区的野生狗牙根遗传多样性研究中同样也发现居群内遗传分化系数达到0.6741,居群间存在较高的遗传分化。狗牙根属于异花授粉植物,在同一居群内杂交现象频发,后代存在较大的遗传分化,导致遗传变异主要来源于居群内;同时不同居群之间由于地理隔离,种子和花粉的广泛传播受到阻碍,贫乏的基因交流使居群内遗传变异大于居群间,使得居群间存在着遗传分化。
3.3 聚类分析
15 个狗牙根居群的聚类分析结果表明各居群没有完全按照地理来源进行聚类,Mantel 检验的结果也显示遗传距离与地理距离之间无相关性。288 份狗牙根材料的聚类分析结果表明,同一居群的材料或相邻居群的材料有优先聚类的趋势,但没有严格按照材料的地理来源进行聚类,在刘杰[10]、刘伟[26]、周少云[8]等的研究中也检测到这种情况。传统的种群遗传学主要是基于距离隔离理论(isolation by distance,IBD)来探讨种群的遗传结构,该理论对解释同一分布区域的不同种群存在高度的遗传分化及种群的渐变样式等存在不足[27-28]。研究发现不同种群因自然选择压力而导致的适应性变异,对种群的遗传结构形成也具有重要影响[29-30],由此建立了适应性隔离理论(isolation by adaptation,IBA)。目前,IBD 和IBA 都被认为是影响种群遗传结构的关键因素,因此在今后的研究中,可以通过分析不同狗牙根居群的环境因子对居群遗传分化的影响,揭示狗牙根种群遗传分化的成因。
3.4 群体遗传结构分析
聚类分析和群体遗传结构分析都能够反映出供试材料的遗传多样性,相比聚类分析,群体遗传结构分析能够更加清晰地反映出材料间的遗传背景和基因交流情况,为后期进行关联分析奠定基础[31-32]。本研究基于贝叶斯模型的群体遗传结构分析方法,将288 份狗牙根材料分为2 个亚群,与288 份狗牙根UPGMA 聚类分析结果基本一致。第1 亚群(144 份)和第2 亚群(96 份)Q值≥0.7,表明供试的288 份狗牙根中绝大多数材料遗传背景单一,基因渗透较少。其余材料(48 份)的Q值<0.7,属于混合型群体,遗传背景较为复杂,存在一定的种质基因渗透。混合群体的形成在其他自交不亲和、异花授粉的多年生植物中也曾有报道[33-34],形成的原因可能是不同区域之间由于人及动物活动,或风力传播将狗牙根花粉、种子或匍匐茎等营养器官带离原居群,造成种质资源的交换和基因交流。群体遗传结构分析是进行关联分析的基础,可以有效消除基因型与性状之间的假关联等不准确因素[35-36]。本研究中供试的狗牙根群体遗传结构较简单,遗传变异较丰富,可以用于开展狗牙根重要性状的关联分析研究。
4 结论
本研究对河南省自然分布的狗牙根进行遗传多样性及群体遗传结构分析,结果表明,河南省狗牙根具有较高的遗传多样性,供试材料没有严格按照地理来源进行聚类,地理距离与遗传距离不具有相关性,居群间存在较大的遗传分化,Structure 群体遗传结构分析结果表明,河南省狗牙根存在一定的种质基因渗透,遗传背景较为复杂。本研究结果可以为河南省狗牙根的合理开发利用和新品种的培育奠定理论基础。
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26 08:55:24
浙江中医药大学学报(2021年6期)2021-07-12 03:05:14
草地学报(2018年5期)2018-11-07 02:25:00
系统工程与电子技术(2016年2期)2016-04-16 05:16:53
中国光学(2015年1期)2015-06-06 18:30:20
中国当代医药(2015年20期)2015-03-01 02:04:33
西南军医(2015年1期)2015-01-22 09:08:26
海岸工程(2014年4期)2014-02-27 12:51:28
中外医疗(2013年23期)2013-08-21 01:45:16
云南中医学院学报(2011年1期)2011-07-31 18:24:35
