
兴奋性和抑制性自反馈压制靠近Hopf 分岔的神经电活动比较*
2022-03-18 10:13黎丽赵志国古华光
物理学报 2022年5期
黎丽 赵志国 古华光
1) (广东省科学院智能制造研究所,广东省现代控制技术重点实验室,广州 510070)
2) (河南工学院理学部,新乡 453003)
3) (同济大学航空航天与力学学院,上海 200092)
突触输入刺激神经元产生的电活动,在神经编码中发挥着重要作用.通常认为,兴奋性输入增强电活动,抑制性输入压制电活动.本文选取可调节电流衰减速度的突触模型,研究了兴奋性自突触在亚临界Hopf 分岔附近压制神经元电活动的反常作用,与抑制性自突触的压制作用进行了比较,并采用相位响应曲线和相平面分析解释了压制作用的机制.对于单稳的峰放电,快速和中速衰减的兴奋性自突触分别可以诱发频率降低的峰放电和混合振荡(峰放电与阈下振荡的交替),而中速和慢速衰减的抑制性自突触也可以分别诱发频率降低的峰放电和混合振荡.对于与静息共存的峰放电,除上述两种行为外,中速衰减的兴奋性和慢速衰减的抑制性自突触还可以诱发静息.兴奋性和抑制性自突触电流在不同的衰减速度下,分别作用在峰放电的不同相位,才能诱发同类压制行为.结果丰富了兴奋性突触压制电活动反常作用的实例,获得了兴奋性和抑制性自突触压制作用机制的不同,给出了调控神经放电的新手段.
1 引言
神经元的电活动,如静息、阈下振荡、峰放电、簇放电和混合振荡(混合振荡为阈下振荡和峰放电两种行为的交替活动)[1,2],是由神经元本身固有的特性和外界输入决定,在神经系统的信息处理中发挥着重要作用[3,4].运用非线性动力学的研究方法和理论,可以揭示神经元的固有特性以及对外界输入响应的不同.例如,根据不变圆上的鞍结分岔和Hopf 分岔附近的不同动力学特征,不仅可以解释1 型和2 型兴奋神经元放电频率不同的原因[1],还可以解释神经元对噪声[5,6]、脉冲输入[7]和突触输入[8,9]等刺激表现出的不同响应.突触输入作为神经元接受刺激的主要来源之一,对神经元的电活动有着重要影响.根据突触对神经元膜电位的影响,可以区分为兴奋性和抑制性突触.通常认为,兴奋性输入可以将膜电位提升到动作电位阈值处,具有促进放电的作用,而抑制性输入压制膜电位使其远离动作电位阈值,具有抑制放电的作用[10-12].然而,由于神经系统表现出的非线性特点,一些奇特的、不同的现象被发现.例如,对应于Hopf 分岔的2 型兴奋神经元,由于其动作电位的阈值曲线所表现出的非线性特点,神经元在收到抑制性输入后可以产生抑制后反跳放电[1,13].根据2 型兴奋神经元的此特点,在模型和实验中构建抑制性自反馈回路,可以使神经元从静息变为连续放电[14].对应于稳定平衡点的静息态,在兴奋性输入之前施加抑制性输入,抑制性输入可以通过抑制后反跳拉近膜电位到阈值曲线的距离,从而促进神经元的放电[15],这一现象也被电生理实验所验证[16].除了对单神经元电活动的影响外,兴奋性和抑制性突触也会使神经元网络产生新的现象[17].通常认为兴奋性耦合促进同步,抑制性耦合压制同步[17].然而,当调节突触衰减速率[17,18]或者引入时滞后[19,20],兴奋性耦合可以抑制同步,抑制性耦合可以促进同步.在神经元网络中,兴奋性突触引发的同步可以使整个神经元网络电活动中止[21].这些发现丰富了对兴奋性和抑制性突触在神经系统中作用的认识,进一步丰富了神经动力系统中的物理现象.
神经元除了接收其他神经元的突触输入外,还可能接收来自于自突触的输入,即神经元通过自突触连接自身,形成自反馈回路[22].自突触已经在海马、大脑皮层和新皮层等神经系统中的多类神经元中被发现[23-25].实验中发现了自突触的一些功能,例如,兴奋性自突触可以引起神经元的持续放电[26]和簇放电[25],而抑制性自突触可以压制神经元的放电,并且可以提高神经元放电时间精确性[23,27].理论仿真发现,自突触在单神经元和神经元网络的电活动调控中发挥着更多的功能和作用[28].对于神经元网络,主要发现自突触可以引起网络的共振、同步和螺旋波等集群行为[29-35].对于单神经元,发现自突触具有调节神经元电活动和放电模式的作用[36,37].例如,兴奋性自突触可以增强神经元的簇放电、引起不规则放电,而抑制性自突触可以抑制神经元的簇放电、促进规则放电[38,39];具有时滞的兴奋性自突触可以引起1 型、2 型和3 型兴奋神经元从静息变为放电[40-43].这些研究结果与通常所认为的兴奋性输入促进放电而抑制性输入压制放电的观点一致.然而,最近的一些研究也给出了不同的认识.具有时滞的兴奋性自突触可以引起簇放电中峰的个数减少[44],抑制性自突触可以引起簇放电中峰的个数增加[45,46].借助于2 型兴奋神经元在抑制性脉冲电流的作用下可以产生抑制后反跳放电这一现象,发现调节抑制性自突触中的时滞,可以引起2 型神经元从静息变为连续放电,并比较了与兴奋性自突触引起连续放电的不同[14,47].相位响应曲线(phase response curve,PRC)主要被用来研究周期振荡的相位在外界摄动下的变化[48].对于周期峰放电,PRC 可以预测神经元在摄动下峰放电时间的变化情况,也通常被用于解释神经元网络同步等现象[9,49,50].根据2 型兴奋神经元对兴奋性/抑制性脉冲电流的相位响应曲线不同,解释了通过改变自突触中的时滞,来调节脉冲自突触电流作用在周期峰放电的相位,使兴奋性自突触表现出压制神经元周期峰放电的作用,使抑制性自突触表现出促进神经元周期峰放电的作用[51,52].随后,进一步的研究发现,根据2 型兴奋神经元在亚临界Hopf 分岔点附近的动力学特点,通过调节兴奋自突触中的时滞,可以引起神经元从放电分别变为静息和混合振荡这两种行为来抑制放电和降低放电频率[52].
兴奋性和抑制性突触是哺乳动物等生物中神经元相互连接的主要方式,在调节神经元及网络放电节律中发挥着重要作用,其调节作用主要依赖于突触电流的特点和神经元固有的动力学特性.在实际神经系统中,兴奋性突触包括AMPA 和NMDA等突触递质,抑制性突触中包括GABAA和GABAB等突触递质[53],不同递质使得突触电流表现出不同的动态特点,例如,突触电流衰减的快慢.2 型兴奋神经元是神经系统中广泛存在的一种神经元类型,并且实验已经发现包括海马锥体神经元和中间神经元、新皮层中间神经元等都表现出2 型兴奋的特征[47,54].为了更多地认识含有不同递质的自突触对2 型兴奋神经元电活动的影响,不同于之前研究[51,52]中调节快速衰减突触模型中的时滞,本文将选取无时滞的、衰减速率可变的突触模型,改变自突触模型中的衰减常数,来调控自突触电流的作用相位,分别研究兴奋性和抑制性自突触对亚临界Hopf 分岔点附近神经元峰放电的影响.在Hopf 分岔附近的峰放电的单稳区以及峰放电和静息的共存区,除了发现之前利用相位响应曲线解释的兴奋性自突触可以引起频率增加和降低的周期峰放电的研究结果外,还发现了引起更低放电频率的混合振荡和静息这两种电活动来压制神经元放电,并利用相轨迹在不稳定平衡点附近和不稳定极限环附近的行为进行了解释,进一步丰富了兴奋性自突触反常规作用的认识.除此之外,也与抑制性自突触的压制作用进行了比较,并给出了兴奋性和抑制性自突触压制神经元放电的差异.该结果不仅丰富了兴奋性输入压制神经元电活动反常作用的结果,也进一步扩展和认识了不同递质的抑制性自突触和兴奋性自突触对神经元电活动的调控作用.
2 模型和方法
为了研究不同衰减特点的自突触对亚临界Hopf 分岔附近放电行为的影响,采用具有2 型兴奋特征的ML 模型[7],具体描述如下:

其中变量V为膜电位;w为钾离子流的激活变量;参数C为膜电容;gNa和ENa分别为钠离子流的最大电导和反转电位;gK,dr和EK分别为钾离子流的最大电导和反转电位;gL和EL分别为漏电流的电导和反转电位;Iapp为常值施加电流.本文所采用参数为:gNa=20 ms/cm2,gK=20 ms/cm2,gL=2 ms/cm2,ENa=50 mV,EK=-100 mV,EL=-70 mV,C=2 μF/cm2,βm=-1.2mV,γm=18 mV,γw=10mV,βw=-13 mV和φw=0.15.Iapp为调控参数.
将自突触电流[18]-gauts(V -Eaut) 引入到ML神经元中,得到自突触耦合的ML 模型,具体如下:

其中s为自突触激活变量;Γ(V)=[1+exp(-10(Vθthres))]-1,θthres=-15 mV为自突触激活阈值;gaut和Eaut分别为自突触的电导和反转电位,其中下标aut可以取为“exc”和“inh”,分别代表兴奋性和抑制性自突触.反转电位Eaut取为Eexc=30 mV 和Einh=-80 mV 来分别保证自突触为兴奋性和抑制性.α和β分别为突触电流的上升和衰减常数.上升常数α决定突触电流上升的快慢,并固定为12 ms-1;衰减常数β决定突触电流衰减的快慢,β值越大,突触电流衰减越快,β越小,突触电流衰减越慢,其可以调节突触电流的作用相位,是本文研究自突触对神经元放电活动影响的主要参数.gexc,ginh和β为调控参数.
模型仿真采用0.001 ms 步长的龙格库塔方法.分岔图(图1(b))和PRC(图2)是基于动力学分析软件XPPAUTO 计算获得[55].

图1 ML 神经元模型(方程(1)和方程(2))动力学行为随Iapp变化 (a) 分岔,红色(虚)实线为(不)稳定平衡点,黑色(空)实心圆为(不)稳定极限环的最小值和最大值;(b) 放电频率Fig.1.Changes of dynamical behavior of the ML neuron model (Equation (1) and (2)) with respect to Iapp :(a) Bifurcations,red (dashed) solid curve denotes (unstable)stable equilibria,and black solid (empty) circles denote minima and maxima of (unstable) stable limit cycles;(b) firing frequency.
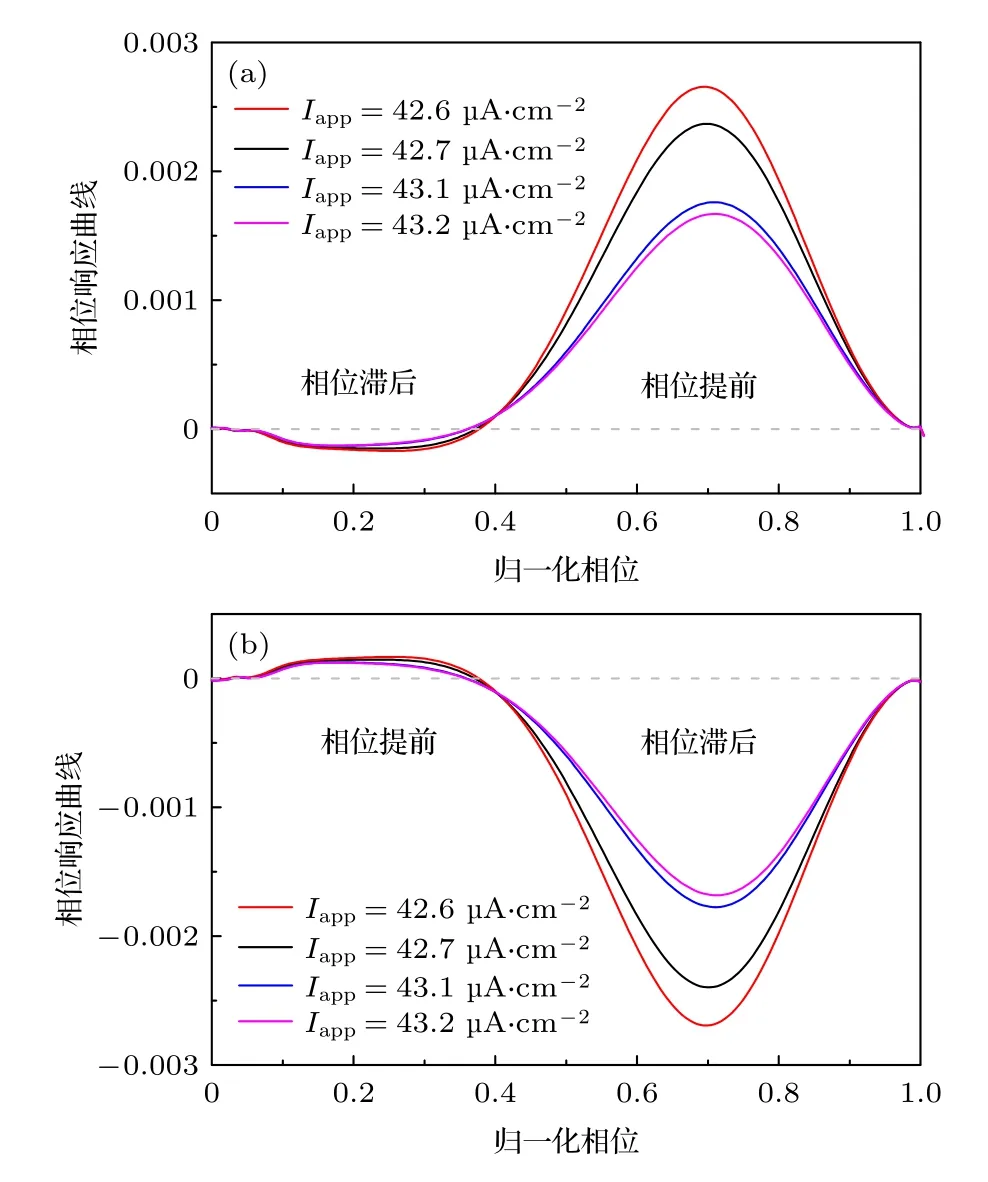
图2 ML 神经元模型(方程(1)和方程(2))对不同电流脉冲的相位响应曲线 (a) 兴奋性电流脉冲;(b) 抑制性电流脉冲Fig.2..Phase response curve (PRC) of the ML neuron model (Equation (1) and (2)) to pulse current stimulations with different effects:(a) Excitatory current pulse;(b) inhibitory current pulse.
3 结果
3.1 无自突触的ML 模型的动力学行为
3.1.1 分岔和放电频率
无自突触的ML 神经元模型(方程(1)和方程(2))存在平衡点,并且在合适的参数可以产生Hopf分岔(见附录A).通过分岔软件XPPAUTO 数值获得的分岔图,如图1(a) 所示.随着Iapp增大,稳定平衡点(红色实线)经过亚临界Hopf 分岔(Iapp≈42.7974 μA/cm2),变为不稳定平衡点(红色虚线),并分岔出不稳定的极限环(黑色空心圆).不稳极限环与稳定极限环(黑色实心圆)碰撞,产生极限环的鞍结分岔Iapp≈42.1785 μA/cm2.当Iapp位于Hopf分岔和极限环的鞍结分岔之间时,神经元表现出静息(稳定平衡点)和周期峰放电(稳定极限环)两种行为的共存,其中不稳极限环为这两种行为吸引域的分界线.当Iapp位于Hopf 分岔的右侧时,神经元仅表现出峰放电.相应地,ML 神经元模型的放电频率随Iapp的变化表现出2 型兴奋的特征,如图1(b)所示.
本文将研究自突触对ML 神经元在Hopf 分岔点附近的峰放电的影响.选取Iapp=42.6 μA/cm2和Iapp=42.9 μA/cm2(Iapp=43 μA/cm2)分别作为共存区和单稳区的代表进行结果展示.为了保证神经元处于峰放电状态,初值取为(V(0)=-20.21999,w(0)=0.01824).
3.1.2 兴奋性和抑制性脉冲激励下峰放电的相位响应
周期峰放电的PRC 可以反映放电频率的增加或降低.对于周期峰放电,其周期或峰峰间隙记为T0.当施加脉冲摄动到峰峰间隙的第一个峰后Δt时,对应相位为θ=Δt/T0,第二个峰出现的时间发生改变.此时,新的周期时间或峰峰间隙变为Tp.第二次放电的相位变化是θ的函数,具体如下:

其中 Δ(θ)代表相位偏差.以θ为横坐标,Δ(θ) 为纵坐标的曲线为PRC.
PRC 采用直接摄动法,通过施加兴奋性或者抑制性脉冲电流到处于放电状态的ML 神经元,获取相应的相位变化情况,其中电流脉冲为持续时间0.05 ms的去极化电流Ipulse=1.0 μA/cm2(图2(a))或者超极化电流Ipulse=-1.0 μA/cm2(图2(b)).
对于2 型兴奋神经元,在兴奋性电流脉冲的摄动下,PRC 在小的θ值时表现出相位滞后(PRC的值小于零),在大的θ值时表现出相位提前(PRC的值大于零),如图2(a)所示.相位滞后(提前)意味着峰峰间隙的第二个峰,在摄动下的放电时间延后(提前).由于抑制性和兴奋性作用相反,对于抑制性电流脉冲摄动,其PRC 与兴奋性摄动的PRC关于横坐标轴对称,即兴奋性摄动下的PRC 相位滞后(提前)区变为抑制性摄动下的PRC 的相位提前(滞后)区,如图2(b)所示.需要说明的是,对于兴奋性刺激的PRC,其相位提前区比相位滞后区对刺激敏感(相位偏差大),而对于抑制性刺激的PRC,相位滞后区更加敏感.
图2 所示,当兴奋性和抑制性自突触提供的电流刺激作用在相位滞后区,可以使峰峰间隙的第二个峰延后,引起放电频率的降低.对于两种突触不同的是:为了诱发放电频率降低的峰放电,兴奋性自突触电流须作用在较小相位,而抑制性自突触电流须要作用在较大相位.本文所用的自突触模型中,衰减常数β可以调节突触电流的作用相位.β较大时,突触电流作用在较小相位,而β较小时,突触电流可以持续作用到较大相位.通过调节兴奋性和抑制性自突触中的参数β可能诱发频率降低的电活动.下文将从这一推测开始研究自突触对峰放电的压制作用.
3.2 自突触诱发频率降低的周期峰放电的比较
引入自突触到ML 神经元模型中(方程(3)—(5)),自突触项未改变ML 神经元模型(方程(1)和方程(2))的平衡点的稳定性和分岔(详见附录B),故可以结合ML 神经元模型(方程(1)和方程2))的特点和自突触耦合神经元模型(方程(3)—(5))中的突触电流来解释自突触项引入引起动力学行为改变.下面将分别考虑兴奋性和抑制性自突触对亚临界Hopf 分岔附近放电的影响.
3.2.1 共存的峰放电Iapp=42.6 μA/cm2
依据上述推测,对于兴奋性自突触,以选取较大的β为例.当gexc=2.0 μS/cm2和β=1.0 ms-1时,兴奋性电流可持续作用到相位滞后区(灰色虚线左侧为第一个峰峰间隙的相位滞后区),诱发出放电频率小于无自突触的神经元峰放电(黑线,61.69 Hz)的周期峰放电(红线,53.22 Hz),如图3(a)和图3(b)所示.
对于抑制性自突触,选取较小β为例.当ginh=1.0 μS/cm2和β=0.3 ms-1时,抑制性自突触电流衰减较慢,会持续作用到的相位滞后区(灰色虚线的右侧为第一个峰峰间隙相位滞后区),由于相位滞后区对抑制性刺激十分敏感(图2(b)),抑制性自突触可以引起频率降低的周期峰放电(红线,46.62 Hz 小于无自突触神经元的61.69 Hz),如图3(c)和图3(d)所示.
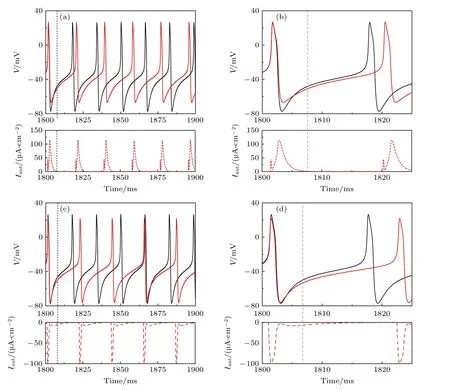
图3 当 Iapp=42.6 μA/cm2时,不同自突触诱导的频率降低的周期峰放电 (a) gexc=2.0 μS/cm2 和 β=1.0 ms-1 的兴奋性自突触;(b) 图(a)的局部放大图;(c) ginh=1.0 μS/cm2 和 β=0.3 ms-1 的抑制性自突触;(d) 图(c)的局部放大图.黑和红色实线分别为ML 神经元模型(方程(1)和方程(2))和自突触耦合ML 神经元模型(方程(3)—(5))的膜电位,红色虚线为自突触电流,灰色虚线为黑色实线第一个峰峰间隙PRC 的相位滞后和提前的临界点,图(a)和图(b)中灰色虚线的左右两侧分为兴奋性刺激PRC 的相位滞后和提前区域,图(c)和图(d)中灰色虚线的左右两侧分为抑制性刺激PRC 的相位提前和滞后区域Fig.3.Periodic spiking with reduced firing frequency induced by different autapses whenIapp=42.6 μA/cm2 :(a) Excitatory autapse withgexc=2.0 μS/cm2andβ=1.0 ms-1;(b) enlargement of Figure (a);(c) inhibitory autapse withginh=1.0 μS/cm2 and β=0.3 ms-1 ;(d) enlargement of Figure (c).Black and red solid curves are the membrane potential for the ML neuron model(Equation (1) and (2)) and the ML neuron model with autapse (Equation (3)—(5)),respectively.Red dashed curve is the autaptic current,gray dashed vertical lines is critical border between phase delay and phase advance regions of the PRC for the first interspike interval of the black solid curve,the left and right sides of the gray dashed line in Figure (a) and Figure (b) respectively are the phase delay and advance regions of the PRC,the left and right sides of the gray dashed line in Figure (c) and Figure (d) respectively are phase advance and delay regions of the PRC.
3.2.2 单稳峰放电Iapp=42.9 μA/cm2
对于单稳峰放电,兴奋性和抑制性自突触也都能诱发频率降低的周期峰放电,结果与共存峰放电类似,为避免重复,本文不再提供该结果.
3.3 自突触诱发共存峰放电变为静息的比较
由于神经元表现出的非线性特点,自突触电流除了能够诱发出频率降低的周期峰放电外,还能诱发静息和混合振荡等放电活动降低的行为.
对于兴奋性自突触,当耦合强度gexc足够大、突触衰减常数β适中时,可以使共存峰放电变为静息.对于Iapp=42.6 μA/cm2的共存的周期峰放电(图4(a1)中的黑色实线).当引入gexc=2.0 μS/cm2和β=0.4 ms-1的兴奋性自突触后,神经元产生一个动作电位(蓝色实线),并在该动作电位激活的突触电流(蓝色虚线)作用下,变为静息(红色实线),如图4(a1)和图4(a2)所示,其中青色圆点为突触电流改变神经元行为的关键相位点.
为了揭示兴奋性自突触诱导共存峰放电变为静息的机制,图4(a1)中膜电位的相轨迹(蓝线)与无自突触时的共存的峰放电(黑线)和静息(红色实心圆)在图4(a3)和图4(a4)中给出.相轨迹(蓝色实线)逆时针旋转,在突触电流的作用下从青色圆点处穿越稳定极限环,然后从不稳极限环(黑色虚线)的左上方穿越进入不稳定极限环内部.在突触电流衰减为零后,相轨迹(红色实线)收敛到稳定平衡点(红色实心圆),形成静息.这就是兴奋性自突触可以引起周期峰放电变为静息的动力学过程.
抑制性自突触诱发的静息如图4 右列所示.当Iapp=42.6 μA/cm2时,引入ginh=1.0 μS/cm2和衰减常数为β=0.1 ms-1的抑制性自突触,神经元膜电位(蓝色实线)在抑制性突触电流(蓝色虚线)的作用下,经一个动作电位,变为静息(红色实线),如图4(b1)和图4(b2)所示.
对应于图4(b2)中膜电位的相轨迹(蓝色和红色实线),在图4(b3)和图4(b4)中给出.在抑制性突触电流持续作用下,相轨迹(蓝色实线)从左下方穿入不稳极限环(黑色虚线)内形成静息.不同于兴奋性自突触改变相平面左上方的相轨迹来诱导静息,抑制性自突触需要改变左下方的相轨迹来诱导静息.由于相轨线逆时针旋转,故对于抑制性自突触,就需要相对于兴奋性自突触更小的突触衰减常数β来保证突触电流衰减更慢,使突触电流作用到相平面左下方的相轨迹来诱导静息.
3.4 自突触诱发峰放电变为混合振荡的比较
3.4.1 共存峰放电Iapp=42.6 μA/cm2
Iapp=42.6 μA/cm2时,无自突触的神经元的行为为峰放电(图4(a1)中的黑色实线,放电频率61.69 Hz).当引入gexc=2.0 μS/cm2和β=0.56 ms-1的兴奋性自突触后,神经元从峰放电变为混合振荡(放电频率为16.44 Hz),如图5(a1) 和图5(a2)所示.
通过相平面上的相轨迹的变化来揭示混合振荡的产生机制,如图5(a3) 和图5(a4) 所示.在单个动作电位激活的兴奋性突触电流的作用下,左上方的相轨迹(蓝色实线) 会运行到不稳定极限环(黑色虚线)附近.在突触电流衰减为零后,相轨迹从不稳极限环附近向外螺旋运动形成阈下振荡(绿色实线),并收敛向稳定极限环(黑色实线)来产生动作电位(蓝色实线),从而形成混合振荡.
当抑制性自突触β值小于兴奋性自突触的β=0.56 ms-1时,也能诱发混合振荡.例如,对于周期峰放电的神经元(图4(a1)中的黑色实线),在引入ginh=1.0 μS/cm2和β=0.263 ms-1的抑制性自突触后,动力学行为变为混合振荡(放电频率10.07 Hz),如图5(b1) 和图5(b2) 所示.在相平面上(图5(b3)和图5(b4)),左下方的相轨迹(蓝色实线)在动作电位激发的抑制性突触电流的作用下,被压制到不稳定极限环(黑色虚线)附近,在突触电流衰减为零后,向外螺旋运动并收敛向稳定极限环来产生混合振荡.

图4 当Iapp=42.6 μA/cm2时,不同自突触诱发共存峰放电(黑色实线) 变为静息(红色实线) (a1)gexc=2.0 μS/cm2 和β=0.4 ms-1的兴奋性自突触;(a2) 图(a1)的局部放大;(a3) 图(a1)的相平面图;(a4) 图(a3)的局部放大;(b1) ginh=1.0 μS/cm2 和β=0.1 ms-1的抑制性自突触;(b2) 图(b1)的局部放大;(b3) 对应于(b1)的相平面图;(b4) 图(b3)的局部放大.在图(a1)和图(b1)中,黑色实线为ML 神经元模型(方程(1) 和方程(2)) 膜电位,蓝色和红色实线为自突触耦合ML 神经元模型(方程(3)—(5))膜电位,蓝色虚线为突触电流,蓝色实线是对应于突触电流的膜电位.在图(a3)和图(b3)中,蓝色和红色实线分别对应于图(a1)和图(b1)中蓝色和红色实线的相轨迹.灰色虚线和点线分别为方程(1)和方程(2)的 w 和 V 零等倾斜线,黑色实线和虚线分别为方程(1)和方程(2)的稳定和不稳定极限环,红色实心点为方程(1)和方程(2)的稳定平衡点,蓝色箭头表示相轨迹的方向,青色实心点为兴奋性和抑制性突触电流改变相轨迹方向的关键相位Fig.4.Resting state (solid red curve) evoked from coexisting spiking (solid black curve) by different autapses whenIapp=42.6 μA/cm2 :(a1) Excitatory autapse with gexc=2.0 μS/cm2 and β=0.4 ms-1 ;(a2) enlargement of Figure (a1);(a3) phase portrait corresponds to Figure (a1);(a4) enlargement of Figure (a3);(b1) inhibitory autapse with ginh=1.0 μS/cm2 andβ=0.1 ms-1;(b2) enlargement of Figure (b1);(b3) phase portrait corresponds to Figure (b1);(b4) enlargement of Figure (b3).In Figure (a1) and Figure (b1),black solid line is the membrane potential of the ML neuron (Equation (1) and (2)),blue and red solid lines are membrane potential of the ML neuron with autapse (Equation (3)—(5)),blue dashed curve is the autaptic current and the blue solid curve is membrane potential effected by the autaptic current.In Figure (a3) and Figure (b3),blue and red solid curves are the trajectories corresponding to the blue and red solid parts in Figure (a1) and Figure (b1),respectively,gray dashed (dotted)curve is the nullcline =0 (=0) of Equation (1) and (2),black solid (dashed) curve is the stable (unstable) limit cycle of Equation (1) and (2),red solid circle is the stable focus of Equation (1) and (2),blue arrow denotes the direction of the trajectory,cyan solid circle is a critical point that determines the direction of trajectory affected by the excitatory or inhibitory autaptic current.

图5 当Iapp=42.6 μA/cm2时,不同自突触诱发共存峰放电变为混合振荡(蓝、绿实线) (a1)gexc=2.0 μS/cm2 和β=0.56 ms-1的兴奋性自突触;(a2) 图(a1)的局部放大;(a3) 对应于图(a1)的相平面;(a4) 图(a3)的局部放大;(b1) ginh=1.0 μS/cm2 和β=0.263 ms-1的抑制性自突触;(b2) 图(b1)的局部放大;(b3) 对应于(b1)的相平面;(b4) 图(b3)的局部放大.在图(a1)和(b1)中,黑色实线为ML 神经元模型(方程(1)和方程(2))膜电位,蓝色和绿色实线为自突触耦合ML 神经元模型(方程(3)—(5))膜电位,蓝色虚线为突触电流,蓝色实线是对应于突触电流的部分.在图(a3) 和图(b3) 中,蓝色和绿色实线分别对应于图(a1) 和图(b1)中蓝色和绿色实线的相轨迹.灰色虚线和点线分别为方程(1)和方程(2)的 w和 V 零等倾斜线,黑色实线和虚线分别为方程(1)和方程(2)的稳定和不稳定极限环,红色实心点为方程(1)和方程(2)的稳定平衡点,蓝色箭头表示相轨迹的方向Fig.5.Mixed-mode oscillations (blue and green solid curves) evoked from coexisting spiking by different autapses when Iapp=42.6 μA/cm2:(a1) Excitatory autapse withgexc=2.0 μS/cm2andβ=0.56 ms-1 ;(a2) enlargement of Figure (a1);(a3)phase portrait corresponds to Figure (a1);(a4) enlargement of (a3);(b1) inhibitory autapse withginh=1.0 μS/cm2 and β=0.263 ms-1;(b2) enlargement of Figure (b1);(b3) phase portrait corresponds to Figure (b1);(b4) enlargement of Figure (b3).In Figure (a1) and Figure (b1),black solid line is the membrane potential of the ML neuron (Equation (1) and (2)),blue and green solid lines are membrane potential of the ML neuron with autapse (Equation (3)—(5)),blue dashed curve is the autaptic current and the blue solid curve is membrane potential effected by autaptic current.In Figure(a3) and Figure (b3),blue and green solid curves respectively are the trajectories corresponding to the blue and green solid parts in Figure (a1) and Figure (b1),gray dashed(dotted) curve is the nullcline =0 (=0) of Equation (1) and (2),black solid (dashed) curve is the stable (unstable) limit cycle of Equation (1) and (2),red solid circle is the stable focus of Equation (1) and (2),blue arrow denotes the direction of the trajectory.
3.4.2 单稳峰放电Iapp=42.9 μA/cm2
类似于兴奋性和抑制性自突触在共存区内诱发的混合振荡,在单稳区内,这两种自突触分别在适中的β和较小的β值下也可以诱发混合振荡.例如,当Iapp=42.9 μA/cm2时,无自突触的神经元行为为周期峰放电(放电频率65.79 Hz),如图6(a1)中的黑色实线所示.引入ginh=2.0 μS/cm2和β=0.35 ms-1的兴奋性自突触,神经元的放电行为变为5.62 Hz 放电频率的混合振荡(图6(a1)和图6(a2)).引入ginh=1.0 μS/cm2和β=0.1 ms-1的抑制性自突触,神经元的放电行为变为2.34 Hz 放电频率的混合振荡(图6(b1)和图6(b2)).这两种突触在单稳区诱导的混合振荡机制也类似于共存区(图5),兴奋性和抑制性自突触电流分别改变相平面左上方和左下方的相轨迹来诱发混合振荡.不同于共存区,相轨迹被作用到不稳定平衡点附近来产生混合振荡(图6).

图6 当Iapp=42.9 μA/cm2时,不同自突触引起的单稳峰放电到混合振荡(蓝和绿实线) 的转变 (a1)gexc=2.0 μS/cm2 和β=0.35 ms-1的兴奋性自突触;(a2) 图(a1) 的局部放大;(a3) 对应于(a1) 的相平面图;(a4) 图(a3) 的局部放大;(b1)ginh=1.0 μS/cm2和 β=0.1 ms-1 的抑制性自突触;(b2) 图(b1)的局部放大;(b3) 对应于(b1)的相平面图;(b4) 图(b3)的局部放大.在图(a1)和图(b1)中,蓝色和绿色实线为自突触耦合ML 神经元模型(方程(3)—(5))膜电位,蓝色虚线为突触电流,蓝色实线是对应于突触电流的部分.在图(a3)和图(b3)中,蓝色和绿色实线分别对应于图(a1)和图(b1)中蓝色和绿色实线的相轨迹.灰色虚线和点线分别为方程(1)和方程(2)的 w和 V 零等倾斜线,黑色实线和虚线分别为方程(1)和方程(2)的稳定和不稳定极限环,红色空心点为方程(1)和方程(2)的稳定平衡点,蓝色箭头表示相轨迹的方向Fig.6.The mixed-mode oscillations (blue and red curves) evoked from the monostable spiking by different autapses when Iapp=42.9 μA/cm2:(a1) Excitatory autapse withgexc=2.0 μS/cm2andβ=0.35 ms-1 ;(a2) enlargement of Figure (a1);(a3)phase portrait corresponding to Figure (a1);(a4) enlargement of Figure (a3);(b1) inhibitory autapse withginh=1.0 μS/cm2 and β=0.1 ms-1;(b2) enlargement of Figure (b1);(b3) phase portrait corresponds to Figure (b1);(b4) enlargement of Figure (b3).In Figure (a1) and Figure (b1),blue and green solid lines are membrane potential of the ML neuron with autapse (Equation (3)—(5)),blue dashed curve is the autaptic current and the blue solid curve is membrane potential effected by the autaptic current.In Figure(a3) and Figure (b3),blue and green solid curves respectively are the trajectories corresponding to the blue and green solid parts in Figure (a1) and Figure (b1),gray dashed (dotted) curve is the nullcline=0 (=0) of Equation (1) and (2),black solid curve is the stable limit cycle of Equation (1) and (2),red hollow circle is the stable focus of Equation (1) and (2),blue arrow denotes the direction of the trajectory.
3.5 自突触参数 β和gexc(ginh)对不同放电行为的影响
为了展示兴奋性自突触和抑制性自突触压制放电的不同,不同动力学行为及放电频率在双参数平面β和gexc(ginh)上的分布在图7 中给出.

图7 不同 Iapp 下,不同自突触诱发的不同动力学行为及其频率在参数平面 β 和 gexc (ginh)上的分布.Iapp=42.6 μA/cm2 (共存峰放电) (a) 兴奋性自突触;(b) 抑制性自突触.Iapp=43 μA/cm2 (单稳峰放电) (c) 兴奋性自突触;(d) 抑制性自突触.黑线为不同电活动的分界线,区域①,②和③分别为频率降低的周期峰放电、混合振荡和静息,区域④为频率增加的周期峰放电.色标代表放电频率Fig.7.Distribution of different dynamical behaviors and firing frequency induced by different autapses on the parameter plane(gexc,β)or (ginh,β)when Iappis fixed at different values,Iapp=42.6 μA/cm2 (coexisting spiking):(a) Excitatory autapse;(b) inhibitory autapse.Iapp=43 μA/cm2 (monostable spiking):(c) Excitatory autapse;(d) inhibitory autapse.Black curve is the border of different firing activities.Regions ①,② and ③are periodic spiking with reduced firing frequency,mixed-mode oscillations,and resting state,respectively,region ④ is periodic spiking with enhanced firing frequency.The color scale represents firing frequency.
3.5.1 峰放电与静息的共存区
对于共存的峰放电(Iapp=42.6 μA/cm2),兴奋性和抑制性自突触诱发不同动力学行为的分布分别在图7(a) 和图7(b) 中给出,区域①,②和③分别为频率降低的周期峰放电、混合振荡和静息.除此之外,还有放电频率增加的峰放电(区域④).对于兴奋性自突触(图7(a)),频率降低的峰放电主要出现在β较大的范围(0.6 ms-1<β <1.0 ms-1),静息和混合振荡出现在中等β值范围(0.2 ms-1<β <0.6 ms-1)且突触电导较强(gexc>0.5 μS/cm2),而放电频率增加出现在β值较小区域.
对于抑制性自突触,与兴奋性的结果明显不同,频率降低的峰放电主要出现在中速衰减速率的突触常数β值范围(0.3 ms-1<β <0.6 ms-1),混合振荡和静息出现在慢速衰减速率的突触常数β值范围(β <0.3 ms-1).
3.5.2 单稳峰放电的不同动力学行为
兴奋性和抑制性自突触对单稳区内峰放电(Iapp=43 μA/cm2)的影响,如图7(c) 和图7(d)所示.与自突触作用下的共存峰放电相比:没有静息态出现;混合振荡区域明显扩大,大概对应于图7(a)和图7(b)的静息区;频率降低的周期峰放电和频率增强的峰放电的范围没有大的变化.静息区的消失是由于没有共存行为引起的.
3.5.3 放电频率的变化
从图7 中的色标变化可以观察到:对于兴奋性自突触,随着β的减小,放电频率先降低后增加,而对于抑制性自突触,放电频率递减.
3.6 自突触诱发频率增加的周期峰放电
图7 中还有频率增加的周期峰放电(区域④).例如,当Iapp=42.6 μA/cm2,gexc=1.0 μS/cm2和β=0.26 ms-1时,兴奋性自突触诱导出频率增加的周期峰放电(87.95 Hz 大于单神经元的61.69 Hz),如图8(a) 和图8(b) 中红色实线所示;当Iapp=42.6 μA/cm2,ginh=1.0 μS/cm2和β=1.0 ms-1时,抑制性自突触诱导出频率增加的周期峰放电,其放电频率为62.5 Hz,大于单神经元的61.69 Hz,如图8(c)和图8(d)中红色实线所示.兴奋性和抑制性自突触压制神经元放电(图2(a)和图2(b))相比较,对于兴奋性自突触,只有β较小时,慢衰减的突触电流才可以作用在的相位提前区域(图2(a)),引起放电频率的增加;而对于抑制性突触,只有在β较大时,快衰减电流仅作用相位提前区域(图2(b)),诱发放电频率的增加.单稳放电的结果与图8 类似,本文不再给出结果.
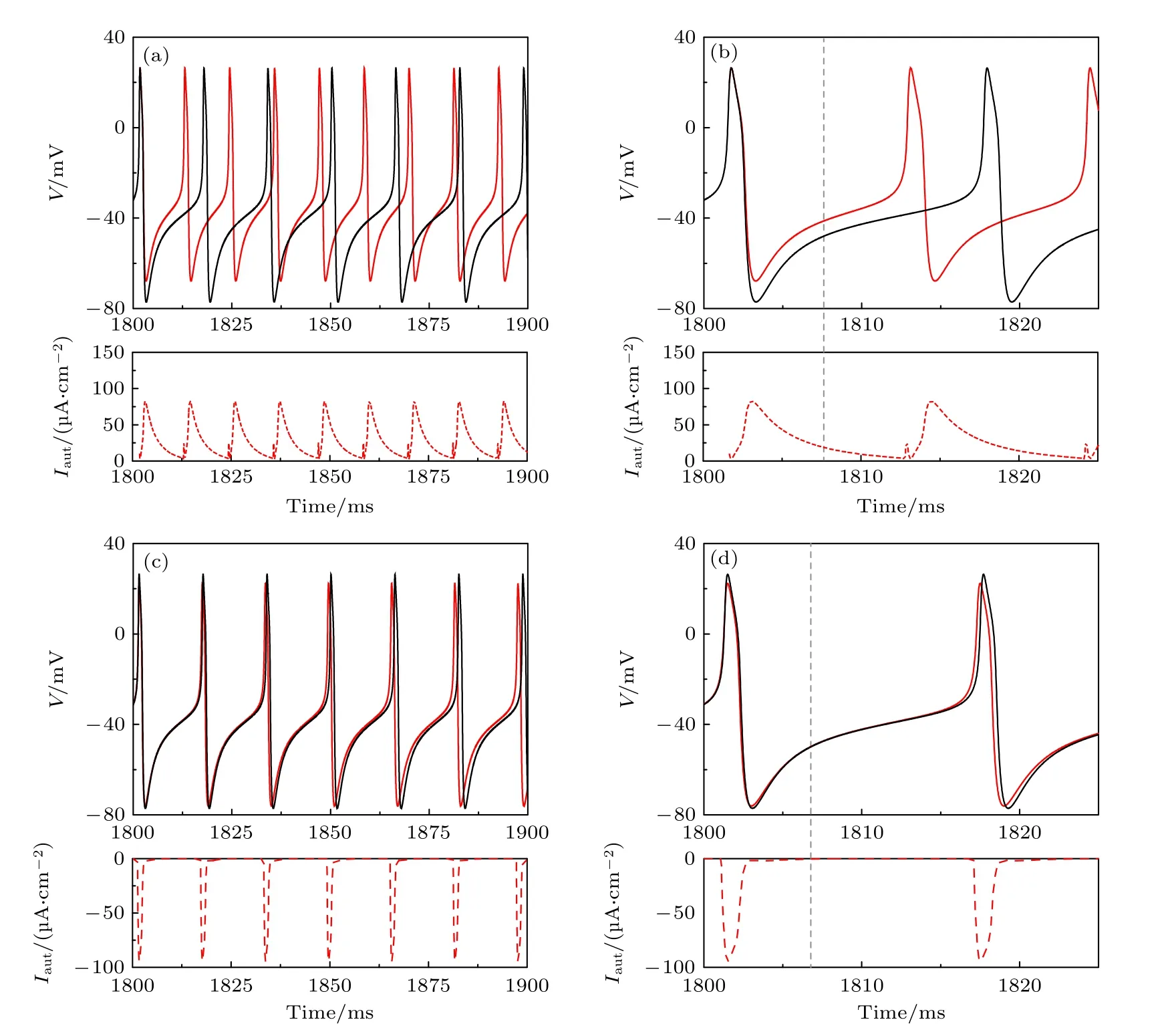
图8 当 Iapp=42.6 μA/cm2时,不同自突触诱导出的频率增加的周期峰放电 (a) gexc=1.0 μS/cm2和 β=0.26 ms-1 的兴奋性自突触;(b) 图(a)的局部放大;(c) ginh=1.0 μS/cm2 和 β=1.0 ms-1 的抑制性自突触;(d) 图(c)的局部放大.黑色实线为无自突触耦合神经元模型(方程(1)和方程(2))的膜电位,红色实线为自突触耦合神经元模型(方程(3)—(5))的膜电位,红色虚线为相对应的突触电流,灰色虚线为黑色实线第一个峰峰间隙PRC 的相位滞后和提前的临界点,图(a)和图(b)中灰色虚线的左右两侧分为兴奋性刺激PRC 的相位滞后和提前区域,图(c)和图(d)中灰色虚线的左右两侧分为抑制性刺激PRC 的相位提前和滞后区域Fig.8.Periodic spiking with increased firing frequency induced by different autapses whenIapp=42.6 μA/cm2 :(a) Excitatory autapse withgexc=1.0 μS/cm2andβ=0.26 ms-1;(b) enlargement of (a);(c) inhibitory autapse withginh=1.0 μS/cm2 and β=1.0 ms-1;(d) enlargement of Figure (c).Black and red solid curves are the membrane potential for the ML neuron (Equation(1) and (2)) and the ML neuron with autapse (Equation (3)—(5)),respectively,red dashed curve is the autaptic current,gray dashed vertical lines is critical border between phase delay and phase advance regions of the PRC for the first interspike interval(the black solid line),the left and right sides of the gray dashed line in Figure (a) and Figure (b) respectively are phase delay and advance regions of the PRC,the left and right sides of the gray dashed line in Figure (c) and Figure (d) respectively are phase advance and delay regions of the PRC.
4 结论
通常观点认为兴奋性输入具有促进放电的作用,而抑制性具有压制放电的作用[10,15].本文根据2 型兴奋神经元以及兴奋性和抑制性自突触的慢衰减的非线性特点,发现兴奋性自突触可以通过诱发放电频率降低的周期峰放电、静息和混合振荡来压制神经元的放电,提供了兴奋性输入压制放电反常作用的多种情况,并获得了与抑制性自突触压制作用的不同特征.根据2 型神经元的PRC,揭示了兴奋性和抑制性自突触降低峰放电频率的原因,根据亚临界Hopf 分岔点附近的动力学行为的特点和突触电流对相轨迹影响,解释了兴奋性自突触诱导静息和混合振荡的动力学机制,获得了兴奋性和抑制性自突触压制神经元电活动差异的原因.
兴奋性自突触和抑制性自突触可以诱导出相同的电活动降低的动力学行为,但诱导出的同一行为出现在不同的突触衰减速率下,说明不同突触参数结合不同的兴奋性或抑制性作用会引起不同的动力学行为,不同衰减特点的自突触具有不同的功能.快速衰减的兴奋性自突触和中速衰减的抑制性自突触可以诱导出频率降低的峰放电.在单稳和共存区,中速衰减的兴奋性自突触和慢速衰减的抑制性自突触可以诱导出具有更低频率的混合振荡.在共存区,除了可以诱导出混合振荡外,中速衰减的兴奋性自突触和和慢速衰减的抑制性自突触还可以诱导出静息.对于这两种自突触,诱导出同一行为出现在不同的衰减常数下,其主要原因是兴奋性和抑制性作用不同,诱发同一行为,需要作用在周期峰放电的不同相位.不同于之前文献中,调节兴奋性自突触中的时滞来诱发频率降低的峰放电、混合振荡和静息[52],本文发现在无时滞的情况下,通过调节兴奋性自突触中的衰减常数,也可以诱发这三种行为,拓宽了兴奋性自突触压制电活动的条件.
本文研究结果从理论上给出了不同衰减速率自突触调节电活动的潜在功能和作用,可以推广到其他模型中,具有重要生物意义.具有不同突触递质的突触,对应的突触电流衰减速率也不同,通过调节突触衰减常数可以获得含有不同递质特点的突触在调控神经元电活动中的作用.本文的结果给出了不同衰减速率的兴奋性和抑制性自突触对神经元放电响应的影响,尤其是兴奋性自突触可以抑制神经元放电响应这一反常作用,丰富了对神经动力系统的认识,揭示了自突触调控神经元电活动的非线性机制,有助于认识不同突触递质调控的自突触在在神经系统中的潜在功能.本文结果可以推广到FitzHugh-Nagumo,Connor-Stevens 和Hodgkin-Huxley 等其他具有亚临界Hopf 分岔的2 型兴奋神经元模型中[47,56,57].对于2 型兴奋神经元,神经元放电频率位于较窄的频率带内,兴奋性和抑制性自突触可以增加和降低2 型兴奋神经元的放电频率,是增大了神经元放电频率的范围,而放电频率范围的增大使神经元具有更强的编码能力,将有助于实现多种生物功能[58-60].例如,灵活易变的放电频率可以缩减皮层工作记忆异常的时间[60].
附录A
对于无自突触的ML 神经元模型(方程(1) 和方程(2)),令方程(2)的右端为零,解得w=w∞(V),并代入方程(1)中,将右端记为

由于gNa,gK,dr和gL为正数,以及m∞(V)>0和w∞(V)>0且为连续函数,故函数f(V)在(-∞,+∞)上至少存在一个实根V*.函数w∞(V)关于V单调递增,将V=V*代入w=w∞(V)仅能解得一个w=w*,故方程(1)和方程(2)至少存在一个平衡点(V*,w*).
ML 神经元模型(方程(1)和方程(2))在平衡点(V*,w*)T的雅可比矩阵为

相应的特征方程为
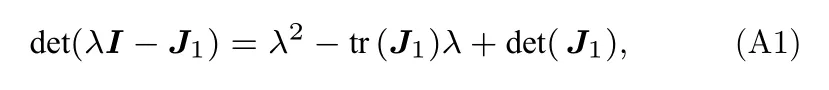
其中I为单位矩阵,tr(J1)=a+d,det(J1)=ad-bc.根据特征根的特点,可以分为如下情况:
情形i:如果 tr(J1)>0或者 det(J1)<0,特征方程(A1)有正根或正实部根,平衡点(V*,w*)不稳.
情形ii:如果 tr(J1)<0,当 det(J1)>0,特征方程(A1)的所有特征根具有负实部,平衡点(V*,w*)稳定;当det(J1)=0时,特征方程(A1) 有零特征根,系统(方程(1)和方程(2))存在鞍结分岔.
情形iii:如果 tr(J1)=0,当 detJ1>0 时,特征方程存在一对纯虚根,系统(方程(1)和方程(2))存在Hopf 分岔;当 detJ1=0 时,特征方程存在二重零根,系统(方程(1)和方程(2))存在Bogdanov-Takens 分岔.
附录B
对于含有自突触的ML 神经元模型(方程(3)—(5)),自突触只有在动作电位刺激后才可以激活,在正常的电生理范围,神经元的阈下活动如静息很难激活自突触电流,例如,对于图1 的静息膜电位V*,Γ(V*)≈0,Γ′(V*)≈0,进而可以从方程(5) 解得平衡点s* ≈0.当s* ≈0 时,模型(方程(3))中的自突触电流项接近于零,即自突触的引入没有改变ML 神经元的平衡点(V*,w*),因此,对于自突触耦合的ML 神经元模型(方程(3)—(5)),平衡点为(V*,w*,s*),其中s* ≈0.
含有自突触的ML 神经元模型(方程(3)—(5))在平衡点(V*,w*,s*)处的雅可比矩阵为

由于s* ≈0,Γ(V*)≈0和Γ′(V*)≈0,上述矩阵又可以写为

相应的特征方程为

方程(B1)的右端第二个括号内的二次多项式与无自突触的ML 神经元模型的特征方程(A1)相同.λ=-β为自突触引入到ML 神经元模型后新增加的特征根.然而,由于β为正,新增加的特征根λ=-β始终为负,不改变原系统(无自突触ML 神经元模型)平衡点的稳定性.综上所述,引入自突触到ML 神经元未改变原系统(无自突触的ML 神经元模型)的平衡点、稳定性和分岔.
猜你喜欢
中小企业管理与科技(2022年19期)2022-12-22
人民交通(2021年17期)2021-09-09
娃娃画报(2019年10期)2019-12-17
中国化工贸易·下旬刊(2019年6期)2019-10-21
诗潮(2019年8期)2019-08-23
阅读(科学探秘)(2019年5期)2019-07-19
教育教学论坛(2019年18期)2019-06-17
中西医结合心血管病电子杂志(2018年28期)2018-11-19
读者·原创版(2017年9期)2017-09-15
科学与财富(2016年38期)2017-05-23
