
化 石
2022-03-17 14:06:30
生物进化 2022年4期
云南澄江发现奇虾类演化树根部成员
奇虾类动物(anomalocarids)是最早称霸海洋的大型掠食动物和顶级捕食者。它们在寒武纪早期的出现,标志着寒武纪大爆发时期金字塔式食物链的存在以及复杂生态系统的建立。奇虾类由最早发现的奇虾属(Anomalocaris)而得名,在分类学上归属于射齿目(Radiodonta)。到目前为止,奇虾类最早的化石记录可追溯至约5.2 亿年前的寒武纪早期,最晚延续至约4.0 亿年前的早泥盆世。它们的演化历史至少有1.2 亿年之久,足迹遍布全球。奇虾类所在的射齿目已发现20 多个属,其中包括30 多个种。这些已发现的属种分属于四个科一级分类单元,即:奇虾科(Anomalocarididae)、抱 怪 虫 科(Amplectobeluidae)、 筛 虾 科(Tamisiocarididae)和赫德虾科(Hurdiidae)。最早发现于云南澄江帽天山的寒武纪早期澄江动物群,是独一无二的化石宝库和世界自然遗产。澄江动物群不仅具有奇虾类确切的最早化石记录,也是迄今保存奇虾类物种多样性最高的化石群。1994 年,由中国科学院南京地质古生物研究所陈均远研究员领衔的研究团队,在美国《科学》杂志上发表了包括两块完整标本在内的澄江动物群奇虾类化石,在学术界和社会大众中引起了轰动。其中一块在1990 年采集于帽天山的奇虾类标本,已成为澄江动物群的标志性符号之一。近期,科学家对包括这块完整的奇虾类标本在内的相关化石进行了精细的形态解剖研究,结果发现,这块经典的奇虾类化石标本在前附肢形态、桨状叶数量、尾扇组成等特征上均不同于其他已命名的奇虾类属种,应属于一个新属新种。研究者以这块化石标本为正模式标本,将其命名为帽天山开拓虾(Innovatiocaris maotianshanensis), 以纪念陈均远先生开拓创新的科学精神和对澄江动物群与寒武纪大爆发研究的贡献。系统发生分析表明,帽天山开拓虾不属于已知奇虾类四个科中的任何一个,其演化位置位于非赫德虾类的基部或赫德虾科的基部,靠近赫德虾科和非赫德虾类的分异点,即接近奇虾类演化树的根部。因此,帽天山开拓虾为奇虾类的起源和演化提供了一个重要参考点。(Journal of the Geological Society, 2022, jgs2021-164)

中国志留纪鱼类化石填补有颌类演化史空白

包括人类在内,地球上现存99.8%的脊椎动物都具有颌骨(上颌与下巴),统称为有颌脊椎动物或有颌类。有颌类的出现与崛起是“从鱼到人”的脊椎动物演化史上最关键的跃升之一,人类的很多重要器官与身体构型都可追溯到有颌类演化之初。较完善的有颌类化石记录一度只能追溯到志留纪晚期(4.25 亿年前),而分子钟推断的有颌类起源时间不晚于奥陶纪晚期(4.5 亿年前),这两者之间存在一段至少绵延三千万年的横跨整个志留纪的巨大空白。近十年来,我国的古生物学家踏遍国内志留纪地层可能含鱼的二百多个地点,终于在华南志留纪早期地层中发现“重庆特异埋藏化石库”和“贵州石阡化石库”,觅得大量特异埋藏保存的完整志留纪早期鱼类化石,找到了破解有颌类最初崛起与辐射分化之谜的钥匙。“贵州石阡化石库”时代为兰多维列世(志留纪早期)埃隆期最晚期,约4.39 亿年前,含有数量多、保存好的有颌类微体化石。其中,双列黔齿鱼(Qianodus duplicis)的齿旋代表最古老的有颌类牙齿,将牙齿最早化石证据前推了1400 万年。而新塑梵净山鱼(Fanjingshania renovata)棘刺的发现显示,早在志留纪早期,原始软骨鱼类已经演化出典型的栅棘鱼形态,同时具有硬骨鱼类的组织学特征。上述进展也让奥陶纪、志留纪鱼类鳞片和棘刺化石分类位置的长期争论尘埃落定。“重庆特异埋藏化石库”时代为兰多维列世特列奇期,约4.36 亿年前,是目前世界上唯一保存志留纪早期完整有颌类化石的特异埋藏化石库,堪称“鱼类的黎明”,将完整有颌类的化石记录前推了1100 万年,将若干人类身体结构的起源追溯到4.36 亿年前的化石鱼类中。其中,无颌的盔甲鱼类灵动土家鱼(Tujiaaspis vividus)为脊椎动物成对附肢起源提供了关键化石证据;有颌的蠕纹沈氏棘鱼(Shenacanthus vermiformis)是迄今所知最早的保存完好的软骨鱼,确证了鲨鱼是从“披盔戴甲”的祖先演化而来。而另一种有颌鱼类奇迹秀山鱼(Xiushanosteus mirabilis)则糅合了多个盾皮鱼大类的特征,为探究有颌类生命之树根部主要类群的起源和脊椎动物头骨演化提供了珍贵资料。(Nature, 2022, 609,7929)
牙结石透露东亚最早现代人食谱
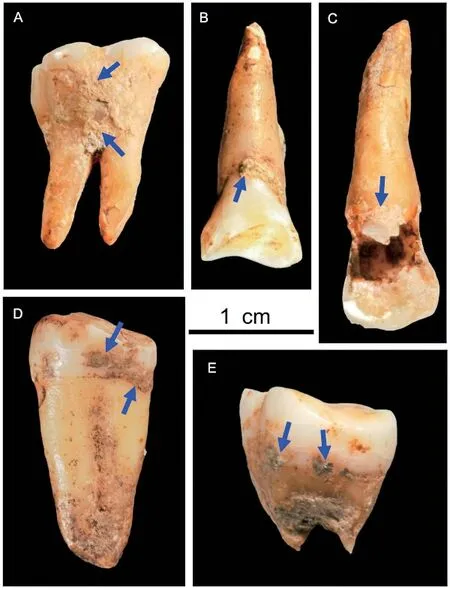
了解人类的食物来源是揭示现代人起源和演化的关键一环。人类演化的成功在很大程度上可以归功于我们获得和消费各种食物的能力。东亚作为人类起源和演化关键区域,至今仍缺少能反映早期现代人食物来源(尤其是植物类食物)的直接化石证据。近日,国内的古生物学家对湖南道县福岩洞出土的东亚最早现代人(12 万—8 万年前)进行了牙结石的提取与分析,发现牙结石中包含橡子、块根、块茎等多种植物产生的淀粉粒。该研究首次提供了东亚早期现代人对富含碳水化合物植物资源消费的直接证据,反映了早期现代人的生存策略及其对环境的适应。此外,研究者还在牙结石内发现了来自针叶植物的具缘纹孔管胞,并在牙齿上观察到许多细小且平行的划痕。这些发现与之前学者报道过的西班牙4.9 万年前尼安德特人牙结石中的植物残存及划痕一致。这表明,福岩洞道县人与尼安德特人一样习惯用松柏类摩擦牙齿,说明东亚早期现代人就已存在可能的剔牙行为。(Frontiers in Plant Science, 2022, 13: 989308)
恐龙蛋化石揭示恐龙灭绝新机制

恐龙自2.35亿年前的晚三叠世出现以来,随着地球环境的更替,属种不断演化,在地球上繁衍了1.7 亿年之久。它们于6600 万年前从地球上消失,其灭绝之谜是世人所关注的热点问题。恐龙属于卵生动物,恐龙蛋的孵化成功率直接决定了恐龙种群的繁盛与否,而恐龙蛋的孵化需要合适的温度、湿度甚至二氧化碳浓度。因此,恐龙蛋作为恐龙在地球上繁衍生息的主要载体,不仅可以反映恐龙的繁殖习性,同时其在地层中的富集埋藏规律也能反映恐龙生存时期的古环境特征。陕西山阳盆地是我国为数不多的可用于研究晚白垩世-古新世恐龙灭绝之谜的陆相盆地。科学家在山阳盆地晚白垩世山阳组中系统采集了1000 多件原位埋藏的恐龙蛋和蛋壳标本,恐龙蛋类型主要为瑶屯巨形蛋、长形长形蛋和坪岭叠层蛋,其母体恐龙分别为窃蛋龙类和鸭嘴龙类,这与山阳盆地恐龙骨骼化石所代表的恐龙类型十分吻合,另外该盆地内还产有少量的暴龙类骨骼。恐龙蛋和恐龙骨骼表明当时盆地内的恐龙多样性处于一个相对较低的水平。年代学结果显示山阳盆地恐龙化石分布的时限是6824 万—6638 万年前,表明山阳盆地内的恐龙多样性在其灭绝前的约200 万年间一直处于比较低的水平。综合秦岭东部其他晚白垩世盆地以及山东莱阳、广东南雄等地发现的恐龙化石,研究人员发现在7200 万年前左右中国的恐龙多样性有一个明显的降低趋势,这与北美西部的恐龙化石分布特征有相似之处。因此,由该研究可推测:在晚白垩世时期,随着自然生态系统和恐龙自身的协同演化,全球的恐龙多样性发生了持续性衰退,降低了恐龙这个类群的环境适应能力,并导致其无法从由德干火山爆发或小行星撞击等重大灾害事件所引起的环境剧变中生存和复苏,从而最终走向灭绝。(PNAS, 2022, 119:e2211234119)
云南发现最早的长臂猿化石

长臂猿科现存20 种,主要生活在中南半岛的热带、亚热带地区,包括我国华南和东南亚。在我国境内现存仅4—6 种。然而,此类化石非常稀少,多数发现于华南和东南亚不老于200 万年前的洞穴地点中。云南元谋新发现的被命名为元谋小猿(Yuanmoupithecus xiaoyuan)的小型猿类,被中国科学家团队确定为迄今发现的最早的长臂猿,这一发现将长臂猿化石记录向前推至距今800 万—700 万年前的晚中新世时期。元谋小猿牙齿和新发现的不完整的左下侧面骨化石的形态特征,与现生长臂猿尤其是冠长臂猿类非常相似,但有些特征更原始,指示其可能为所有现生长臂猿的祖先。根据元谋小猿牙齿大小计算,元谋小猿与现生长臂猿的平均体重相近,大约为6 千克左右。尽管元谋小猿标本非常稀少,但经过细致的分析对比研究,最终可以证实元谋小猿是现生长臂猿最可能的直接祖先。而此前发现于印度中中新世(距今1380 万—1250 万年)的一枚下第三臼齿(被命名为Kapi ramnagarensis)则不是原研究者所声称的更早的长臂猿,而是与现生猿类没有密切关系的更原始的灵长类,可能与印度晚中新世古猿有亲缘关系。元谋小猿的发现填补了东亚小型猿类进化史的一段空白。遗传学研究表明,长臂猿大约是在距今2200 万—1700 万年之间从人和猿共同祖先的支系分化出来的,而冠长臂猿类祖先分化的时间大约为800 万年前。此次发现和之前云南昭通发现的金丝猴最早的祖先——中猴化石的证据表明,分子证据与化石证据推断的物种起源时间基本吻合,古生物形态学和分子生物学研究结果可以相互印证。然而,关于长臂猿科的演化,目前还有1000多万年的“缺环”有待发现,在华南和东南半岛持续的野外科考可能发现有关长臂猿进化史的更多关键证据。距今600 万—200 万年间的空白时段尤其值得关注,因为这个时段正是古老型灵长类动物群和现代型灵长类动物群交替的关键时期。(Journal of Human Evolution,2022, 171: 103251)
猜你喜欢
当代水产(2022年8期)2022-09-20 06:47:02
中老年保健(2022年7期)2022-09-20 01:19:00
云南农业科技(2021年6期)2021-12-30 06:29:00
学苑创造·A版(2021年10期)2021-10-30 10:34:43
文化创新比较研究(2020年10期)2021-01-14 01:06:20
农民致富之友(2020年27期)2020-09-12 14:07:33
创造(2020年12期)2020-03-17 08:59:12
民族音乐(2019年2期)2019-12-10 13:14:55
西藏农业科技(2019年1期)2019-07-25 00:37:02
民族音乐(2016年2期)2016-04-04 16:58:36
