
不同季节桃金娘光合特性与光系统PSII活性研究
2022-03-15 09:07:14杨通文高秀梅韩维栋
西南农业学报 2022年12期
杨通文, 高秀梅, 韩维栋
(广东海洋大学滨海农业学院, 广东 湛江 524088)
【研究意义】光合作用是植物有机物积累和代谢的重要途径,是有机物生产的基础,决定植物的生产力[1],光合作用受温度、光照辐射、蒸腾速率、气孔导度、胞间CO2浓度等因子影响,这些因子随季节变化而改变,在不同季节里生态因子对植物光合作用影响程度也会发生改变[2]。植物光合作用内的净光合速率、蒸腾速率、气孔导度、胞间CO2浓度等光合参数能够反应植物对光能的利用能力[3]。植物叶片内叶绿素荧光参数包含丰富的光合作用信息[4-5],能够更加直观地展示植物光化学活性,反应叶片光合作用过程中光系统对光能的吸收、传递、耗散、分配等方面具有独特作用, 与“表观性”的气体交换指标相比, 更能反映“内在性”特点[6]。叶绿素荧光测定技术能够灵敏反应植物光系统PSII反应中心氧化-还原状态、类囊体膜的高能状态和色素间的能量传递效率[7];植物荧光变化容易受到外界环境的影响,可以直接或间接了解植物光合作用过程,反应植物光合能力的重要参数[8]。叶绿素荧光测定技术具有快速、灵敏、非破坏测量等优点,在植物生理研究、作物育种、栽培管理等领域中得到广泛应用[9-11]。桃金娘(Rhodomyrtustomentosa)是桃金娘科,属灌木,野生桃金娘资源在我国广泛分布于海南、广西、云南、江西、台湾、贵州和湖南南部地区[12-13]。果实含有多种人体需要的微量元素和多糖、多酚以及黄酮类有机物具有抗氧化、抗菌、抗病毒、降糖和抑制乙酰胆碱酶活性等药理作用[14-16];桃金娘的株型大小与生长环境相适应,叶小且革质化,根系发达,耐高温和干旱,对水肥要求不高,移栽容易成活;开花期长达到一个月以上,在气温、水肥管理以及修剪合适的情况下,叶片能够不断新发出来,在城市绿化建设、园林应用等方面也具有潜在价值[17]。【前人研究进展】国内外关于桃金娘的研究集中在根、茎、叶和果实的化学成分以及所含化合物提取和药用价值应用方面;其次是种子发芽处理和苗的栽培移植等方面[18];最后是关于桃金娘在我国地理区域分布和种子传播等方面[13, 19]。【本研究切入点】植物光合作用强弱与环境因子有极大关系,环境因子随着季节变化而改变,植物为了适应环境生理生化也会做出相应变化。【拟解决的关键问题】本试验以3年生桃金娘苗为试验材料,研究在不同季节下光合日变化与环境因子的关系以及不同季节叶绿素荧光参数变化情况,为以后桃金娘在生理生化研究方面奠定基础,对桃金娘推广种植和移植栽培提供理论指导。
1 材料与方法
1.1 试验地概况
广东省湛江市麻章区广东海洋大学(湖光校区)位于110°17′ E,21°9′ N,处于北回归线以南,属于热带和亚热带地区,海拔25 m,离海近,全年受到海洋气候的调节,夏天炎热多雨,冬无严寒,降雨量少,年平均降雨量1400~1700 mm,年平均气温23 ℃,土壤偏酸性,多为红壤、砖红壤土等,适合热带、亚热带植物生长,多数为试验用地,植物大多数为农作物和试验树种,周围植物生长丰富。
1.2 试验材料
2020年10月从广州苗圃公司购买3年生桃金娘苗100株,种植在广东省湛江市广东海洋大学(湖光校区)种苗实验实习基地,每株间隔50 cm,2021年1月存活95株,不定期除草施肥浇水。
1.3 试验方法
1.3.1 光合指标测量 于春季(4月)、夏季(7月)、秋季(10月)、冬季(1月)之中选取连续晴天的3 d测量,测量时间为7:00—18:00,每隔1 h测量1次,使用便携式光合仪(LCi-SD)测量,选取桃金娘顶端向下展开第2~3片叶片,共选取10株,每株1片叶子,每片叶子测5次,取总平均值。测定光合指标:有效光合辐射(PAR)、叶片温度(Ti)、空气CO2浓度(Ca)、空气相对湿度(RH)、净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、气孔导度(Gs)、叶片水分利用率[(WUE)=Pn/Tr]、叶片光能利用率[(LUE)=Pn/PAR]。
1.3.2 荧光参数测量 于春季、夏季、秋季、冬季各选3 d,每天9:00 —11:30,使用叶绿素荧光氟笔(FP110 PAR-Fluor Pen)测量桃金娘荧光参数,同样是由顶端向下选取第2~3片叶子,共10株,使用仪器配带的夹子,夹住叶片进行20 min暗处理,然后进行各项荧光参数指标的测定。
非化学淬灭(NPQ)协议曝光和暗恢复阶段持续时间、脉冲数量、间隔时间见表1。

表1 曝光和暗恢复阶段的持续时间以及脉冲之间的数量和间隔
OJIP协议:最小荧光(Fo)、最大荧光(Fm)、可变荧光(Fv)、PSII潜在活性(Fm/Fo)、PSII光能转化率(Fv/Fm)、单位反应中心吸收的光能(ABS/RC)、单位反应中心捕获的用于还原Qa的能量(TR0/RC)、单位反应中心捕获的用于电子传递的能量(Eto/RC)、单位反应中心耗散掉的能量(Dio/RC)。
非化学淬灭(NPQ)协议测定荧光参数:光稳态下化学淬灭(QP-Lss)、光稳定下非化学淬灭(NPQ-Lss)、光称定下量子产率(QY-Lss)最大量子产率(QY-max)和Kautsky诱导峰值。Kautsky诱导峰值:样本暗处理使其光系统完全开放后,将样品暴露在光照下持续几十到几百秒后引出一种叫做Kautsky效应的舜态。
光曲线(Lc)协议:光照强度设置10、20、50、100、300、500、1000 μmol/(m2·s)等7个梯度,测量荧光参数:最大荧光(Fm)、可变荧光(Fv)、量子产率(QY)。
1.4 试验数据处理
试验测量数据使用Excel 进行整理和作图,使用SPSS 22.0软件对数进行相关分析和多元线性回归分析。
2 结果与分析
2.1 桃金娘不同季节生态因子日变化
由图1可知,日均有效光合辐射(PAR)春季[1133 μmol/(m2·s)]>夏季[1121 μmol/(m2·s)]>秋季[997 μmol/(m2·s)]>冬季[982 μmol/(m2·s)];叶片温度(Ti)春季和夏季最高温度达40 ℃以上,日均温春季(38.3 ℃)>夏季(38.1 ℃)>秋季(34.7 ℃)>冬季(31.9 ℃);冬季空气CO2浓度(Ca)高于其它3个季节,Ca日均值冬季(432 mmol/mol)>秋季(394 mmol/mol)>春季(384 mmol/mol)>夏季(380 mmol/mol);空气相对湿度(HR)日变化总体均呈现先下降后上升趋势。
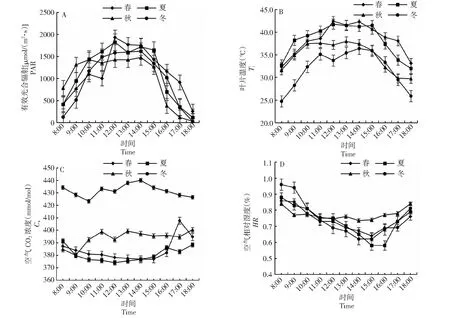
图1 桃金娘不同月份下生态因子日变化Fig.1 Diurnal variation of ecological factors in different months of R. tomentosa
2.2 桃金娘不同季节净光合速率(Pn)与生理因子日变化
由图2-A可知,夏季桃金娘蒸腾速率(Tr)日变化呈“双峰型”,春季、秋季、冬季桃金娘Tr日变化呈“单峰型”;夏季Tr峰值[3.77 mmol/(m2·s)]、日均值[2.79 mmol/(m2·s)],高于其它3个季节。由图2-B可知,秋季Pn日变化呈“单峰型”,春季、夏季和冬季Pn日变化均呈“双峰型”;春季在14:00时出现“光合午休”,夏季在13:00时出现“光合午休”,冬季在15:00时出现“光合午休”;日均Pn:秋季[7.74 mmol/(m2·s)]>夏季[5.91 mmol/(m2·s)]>冬季[4.97 mmol/(m2·s)]>春季[4.09 mmol/(m2·s)],Pn峰值:秋季[11.8 mmol/(m2·s)]>冬季[9.18 mmol/(m2·s)]>夏季[7.11 mmol/(m2·s)]>春[6.21 mmol/(m2·s)]。由图2-C可知,夏季、秋季和冬季桃金娘胞间CO2浓度(Ci)日变化呈“U型”,早上和傍晚高,中午低,春季桃金娘Ci日变化呈“W型”;Ci日均值冬季[287 mmol/(m2·s)]>秋季[268 mmol/(m2·s)]>夏季[267 mmol/(m2·s)]>春季[197 mmol/(m2·s)]。由图2-D可知,桃金娘夏季和秋季气孔导度(Gs)日变化均呈现先下降后上升再下降趋势,冬季Gs日变化呈“M型”;气孔导度日均值:夏季[0.17 mmol/(m2·s)]>秋季[0.14 mmol/(m2·s)]>冬季[0.09 mmol/(m2·s)]>春季[0.05 mmol/(m2·s)]。
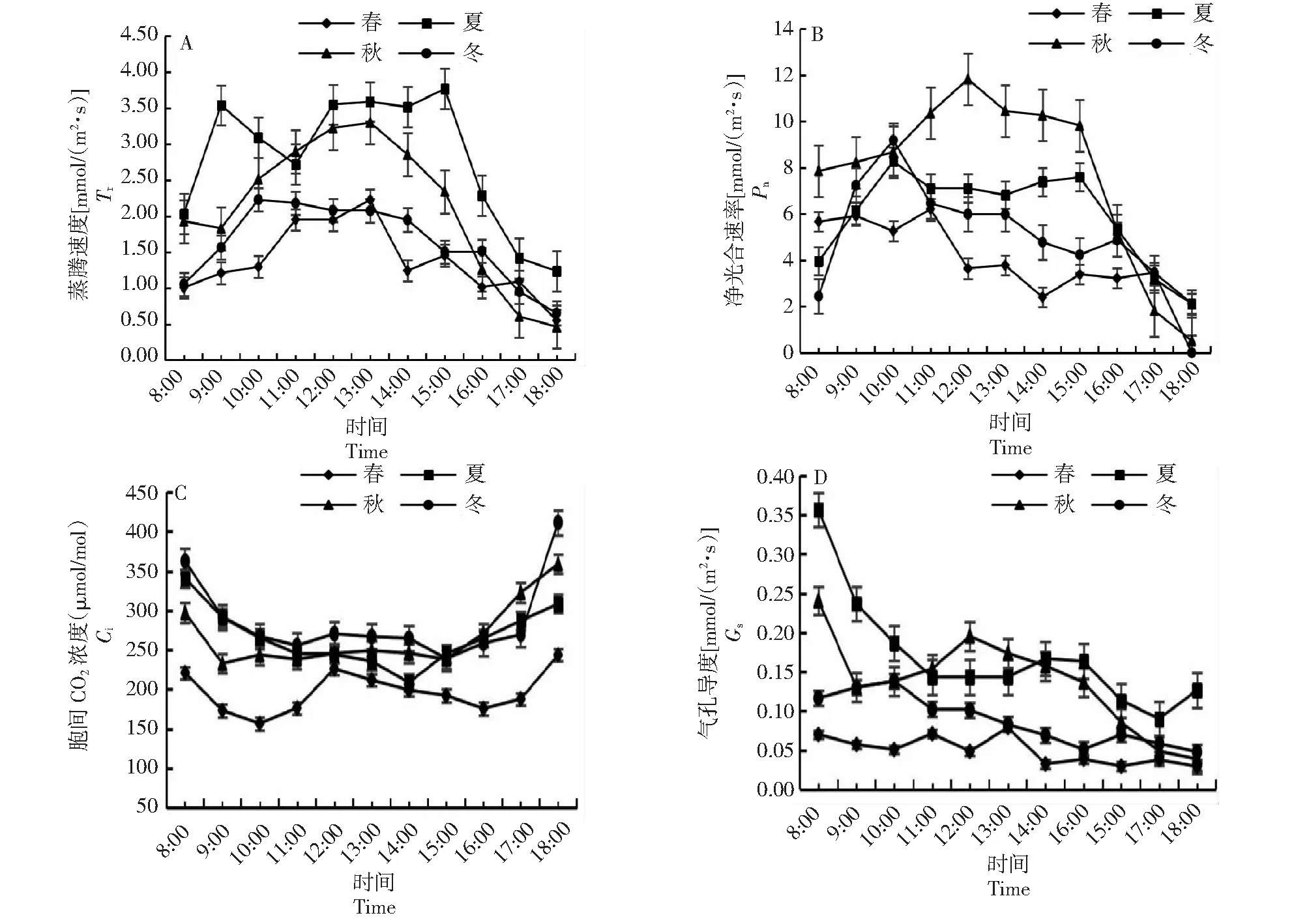
图2 桃金娘不同月份净光合速率,蒸腾速率、胞间CO2浓度、气孔导度日变化Fig.2 Diurnal changes of net photosynthetic rate, transpiration rate, intercellular CO2 concentration and stomatal conductance of R.tomentosa in different months
2.3 桃金娘不同季节水分利用率(WUE)和光能利用率日变化(LUE)
由图3-A可知,春季、夏季和冬季桃金娘对光能利用率(LUE)日变化总体均呈“U型”,秋季桃金娘LUE日变化呈“W型”;秋季日均LUE(0.0097 mmol/mol)最高,春季日均LUE(0.005 mmol/mol)最低。从图3-B可知,春季和冬季水分利用率(WUE)日变化呈“U型”,夏季和秋季WUE日变化呈“M型”;日均WUE:秋季(3.51 mmol/mol)>春季(3.26 mmol/mol)>冬季(2.90 mmol/mol)>夏季2.12 mmol/mol。
2.4 桃金娘不同季节净光合速率(Pn)与生理生态因子相关分析
从表2可知,桃金娘春季净光合速率(Pn)与HR、Gs呈正相关;夏季净光合速率(Pn)与PAR、Ti、Tr呈极显著正相关,与Ca呈极显著负相关,与Ci呈负相关;秋季净光合速率(Pn)与PAR、Ti、Tr、Gs呈极显著正相关,与Ci呈极显著负相关,与HR呈负相关;冬季净光合速率(Pn)与Tr呈极显著正相关,与PAR、Gs呈正相关,与Ci呈负相关。

表2 桃金娘春季、夏季、秋季、冬季净光合速率(Pn)与生理生态因子相关矩阵
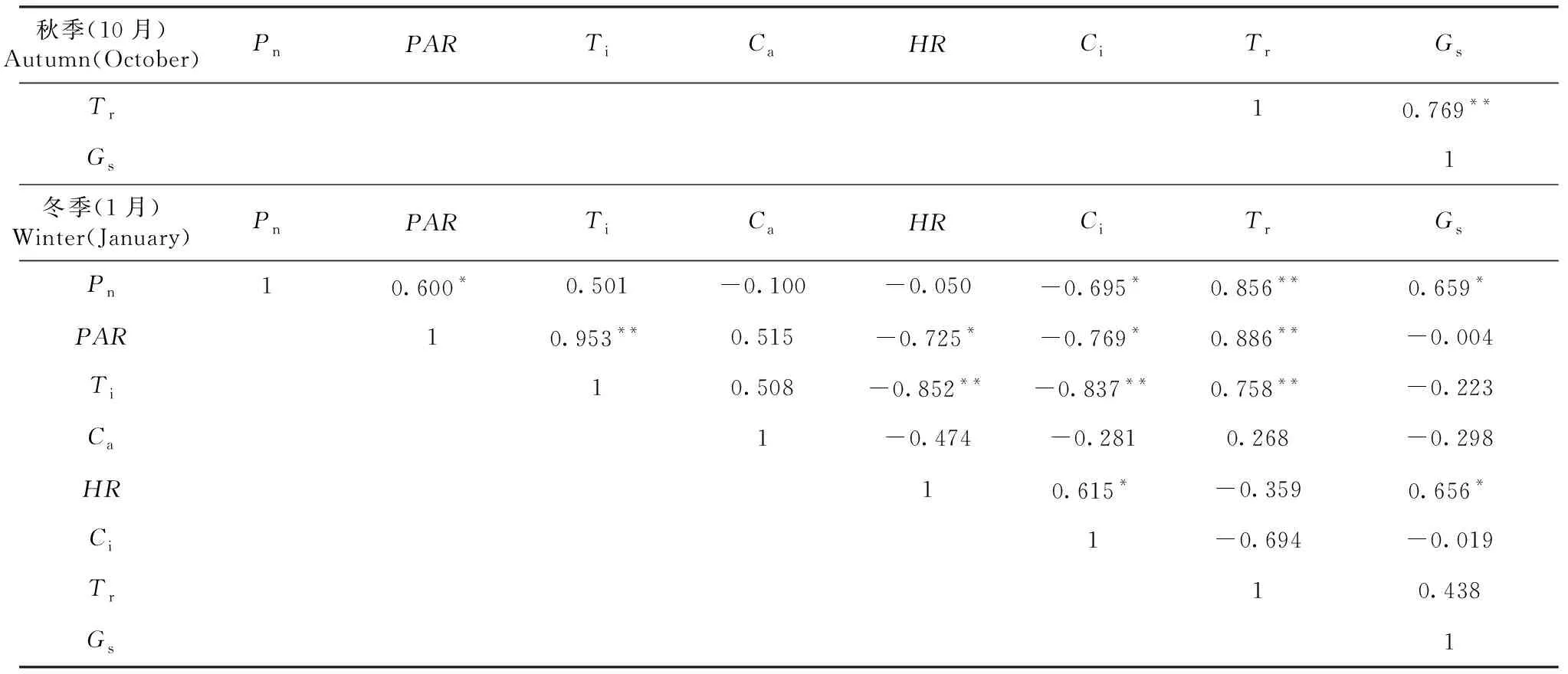
续表2 Continued table 2
对不同季节桃金娘Pn进行正态分布检测采用小样本Shapiro-wilk检测(春季sig.=0.229,夏季sig.=0.153,秋季sig.=0.051,冬季sig.=0.974),检测结果值均大于0.05符合正态分布,可以进行线性回归方程分析。对不同季节桃金娘Pn(Y)日变化与生理生态影响因子(XCi、XGs、XCa、XTi、XPAR、XTr)日变化进行逐步多元线性回归方程分析。得到不同季节下桃金娘最优多元线性回归方程(表3):Gs、HR、Ci是影响春季桃金娘Pn的主要因子,PAR是夏季净光合速率主要影响因子,Tr、Gs、Ci是秋季净光合速率主要影响因子,Tr、Ca是冬季净光合速率主要影响因子。
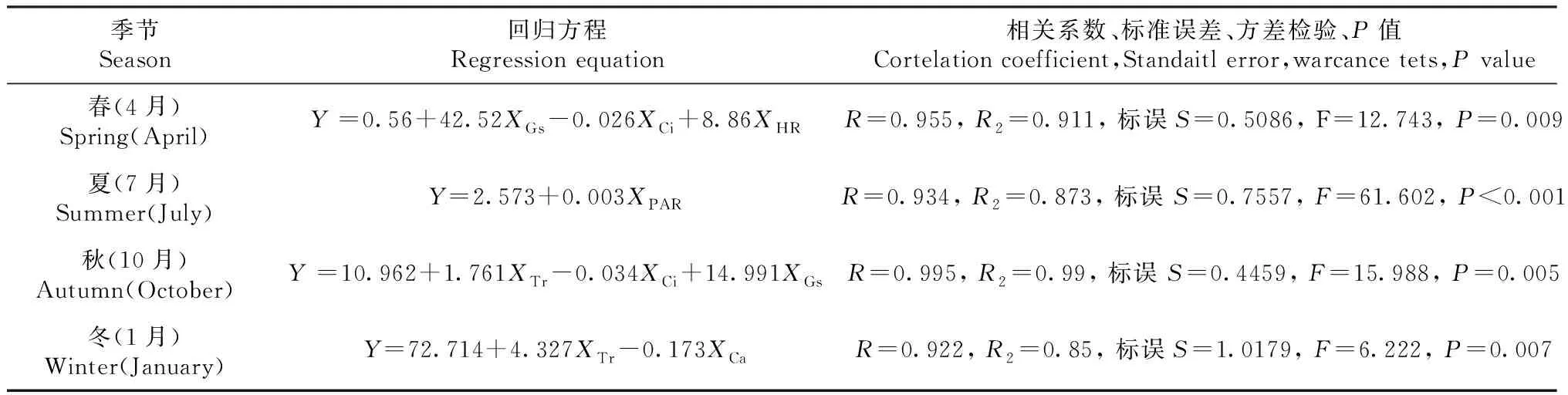
表3 不同季节桃金娘多元线性回归方程
2.5 不同季节桃金娘光系统PSII活性参数
2.5.1 桃金娘不同季节Fo、Fm、Fv值 从图4~5可知,桃金娘夏季的Fo、Fm、Fv值最大,冬季的值最小;夏季的光系统PSII的潜在活性(Fm/Fo)和PSII光能实际转化率(Fv/Fm)最高,其数值分别为2.76、0.64,春季和秋季居中,冬季桃金娘的光系统PSII的荧光数值最低。
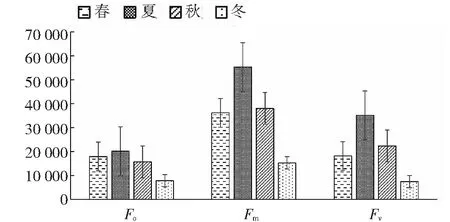
图4 不同季节桃金娘的Fo、Fm、Fv值Fig.4 The Fo, Fm and Fv values of R. tomentosa in different seasons
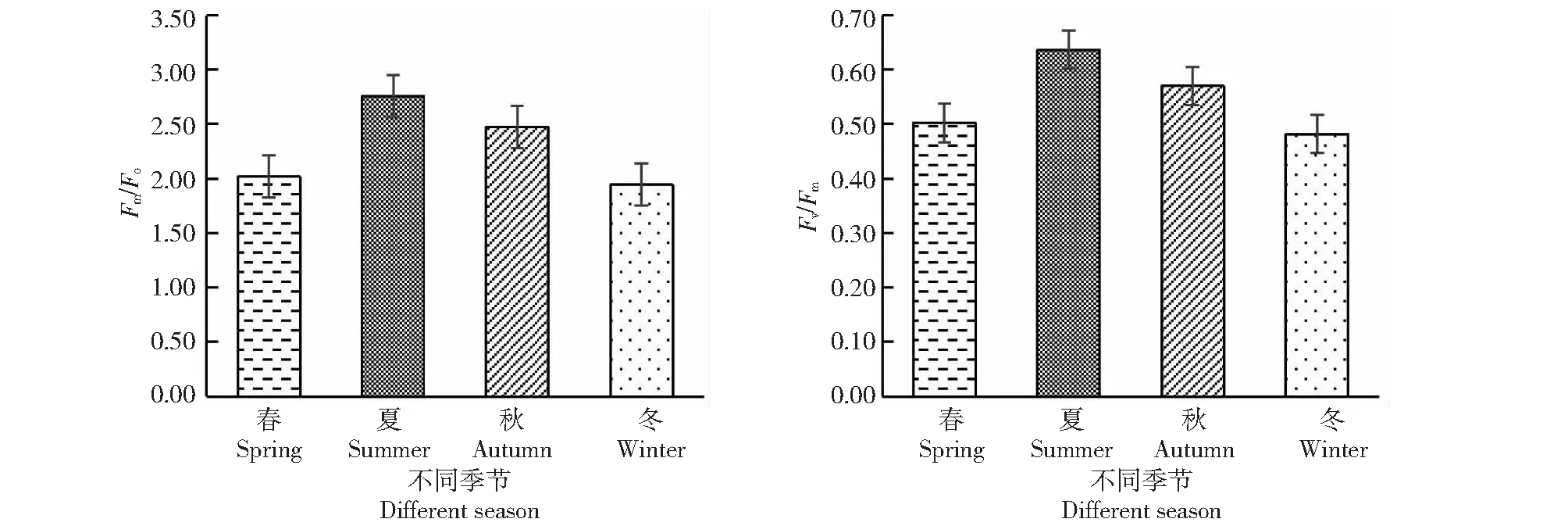
图5 不同季节桃金娘光系统PSII的Fm/Fo、Fv/Fm值Fig.5 Fm/Fo and Fv/Fm values of PSII of R.tomentosa optical system in different seasons
2.5.2 不同季节桃金娘光系统中单位反应中心能量捕获、利用以及消散情况 从图6可知,桃金娘光系统中反应单位捕获能量多少、利用以及消耗等情况,春季桃金娘的单位反应中心吸收(ABS/RC)的能力最大,冬季最小;夏季用于还原质体醌a的能量(TR0/RC)最大,冬季最小;秋季用于电子传递的能量(Eto/RC)最多,春季最小;春季单位反映中心耗散掉的能量(Dio/RC)最多,夏季最小。

图6 不同季节桃金娘单位反应中心能量吸收、分配以及消耗情况Fig.6 Energy absorption, distribution and consumption of R. tomentosa unit reaction center in different seasons
2.5.3 桃金娘不同季节下的化学淬灭(Qp-Lss)与非化学淬灭(NPQ-Lss) 从图7-A~7-B可知,夏季桃金娘的Kuatsky值最大,其次是秋季和春季、冬季最小。在光稳态条件下,夏季的Fm最大,冬季最小;春季、冬季的化学淬灭(QP_Lss)高于夏季、秋季,夏季、秋季的非化学淬灭(NPQ_Lss)高于春季和冬季,夏季、冬季的量子产率(QY_Lss)高于春季和秋季。暗处里后的量子产率(QY_max)结果是夏季=冬季>秋季>春季。

图7 不同季节下桃金娘Kuatsky值、暗处里下QY和光稳态下QP、NPQ、QY值Fig.7 R.tomentosa Kuatsky value, QY in the dark and QP, NPQ, QY in light steady state in different seasons
2.5.4 不同光照强度下桃金娘的量子产率(QY)、光系统PSII活性(Fv/Fm) 从图8-A~8-B可知,春、夏、秋3季量子产率(QY)随着光照强度增加呈下降趋势,冬季的量子产率(QY)随着光照强度增加呈先上升后下降趋势,光照强度为50 μmol/(m2·s)时冬季桃金娘的QY最大;随着光照强度增加,桃金娘春、夏、秋、冬四季的PSII光能转化率(Fv/Fm)总体呈上升趋势,光照强度大于100 μmol/(m2·s)时PSII光能转化率(Fv/Fm)上升相对较快。

图8 不同光照强度下桃金娘的QY、Fv/Fm值Fig.8 QY,Fv/Fm values of R.tomentosa under different light intensities
3 讨 论
光合作用是植物生长发育的重要代谢途径,净光合速率是衡量植物光合作用的一个重要指标[20],净光合速率受外界生态因子和植物自身生理因子的影响,生理生态因子的变化与季节变化具有很大关系[21]。本试验研究显示:桃金娘冬季净光合速率与蒸腾速率呈极显著正相关,春季净光合速率与生理生态因子没有存在极显著的相关性,这一研究结果与刘旻霞等[22]对黄土高原中部3种典型绿化植物光合特性的季节变化研究结果一致。多元线性回归分析能够在多个因子中找出对因变量影响最大的因子[23],桃金娘净光合速率与生理生态因子做线性回归分析,找出影响净光合速率的主要因子,本研究结果显示,春季影响净光合速率(Pn)的主要因子有Gs、HR、Ci, 夏季则是PAR 为主要因子,Tr、Gs、Ci是影响秋季Pn主要因子,春季影响Pn的主要因子是Tr、Ca, 随着季节变化对桃金娘Pn影响的主要因子也随着改变。
植物的光能利用率(LUE)和水分利用率(WUE)在一定程度上可以反应植物在不同环境下资源配置,也是一种抗逆性的重要参考指标[24]。研究结果显示,桃金娘在不同季节对水分和光能利用不同,秋季WUE 最高,夏季WUE 最小,可能与湛江气候变化有关;秋季LUE 最高,同时光能利用率时间相对与其它3季较长。
植物光合日变化出现“光合午休”是植物适应环境的一个重要策略[25-26],许大全等[27]对植物“光合午休”生理特征研究表明,Pn下降、Gs下降、Ci下降为气孔限制所引起的“光合午休”;Pn下降、Gs下降、Ci上升为非气孔限制引起的“光合午休”。本研究结果显示:桃金娘春季、夏季、冬季Pn日变化都存在“光合午休”,春季Pn下降、Gs下降、Ci下降,表明春季“光合午休”是由气孔限制所引起的;夏季Pn下降、Gs下降、Ci上升,表明夏季“光合午休”由非气孔限制引起;冬季Pn下降、Gs下降、Ci下降,表明冬季“光合午休”也是由气孔限制引起,不同季节引起桃金娘“光合午休”类型各不相同。
植物荧光与光合作用密切相关,能够反应叶片叶绿体内部一些机构情况[4],暗适应后Fo指PSII反应中心完全开放的是荧光,Fo越大表明PSII反应中心遭到的破坏越严重[28-29];植物PSII反应中心完全关闭时荧光水平最大,受到光抑制时Fm值下降。Fm/Fo表示光系统PSII潜在活性[30]。本研究显示,夏季的Fo、Fv、Fm最大,冬季Fo、Fv、Fm最小,可能是夏季生态因子中有效光合辐射相对较高造成Fo升高,有效光合辐射升高使叶片温度升高,进而提升光系统PSII中相关酶活性升高,夏季光系统PSII潜在活性(Fm/Fo)高于其它3季,光系统PSII实际转效率(Fv/Fm)是夏季>秋季>春季>冬季,桃金娘夏季最大荧光、可变荧光增大可能有利于增加光能的存储量以及提高利用率。
植物光合作用吸收的光能要转变成稳定的化学能主要经过光系统PSII等一系列电子传递,把光能转化为不稳定的化学能ATP,然后在经过碳同化形成稳定有机物[31]。在整个光系统PSII过程中质体醌周转次数、质体醌库大小、质体醌还原速率等都会影响光合作用[32]。本研究结果显示,春季ABS/RC最多,同时Dio/RC也最大;夏季对TRO/RC利用使最高,消散掉的能量最少;秋季Eto/RC最大,不同季节下桃金娘光系统PSII对捕获能量分配的策略也各有不同。
当大量过剩光能不能及时用于光化学反应时, 就能形成光抑制现象, 植物为了避免PSII受到破坏, 往往通过叶黄素循环、荧光发射、光呼吸等热耗散途径来消耗过剩光能[33]。Kautsky峰值指将植物叶片暗适应后突然暴露在光照下,叶绿体分子吸收能量后由低能量的稳态转变为高能量不稳定的激发态,激发态的电子不稳定寿命非常短很快又回到稳态中去,在此过程中小部分激发态能量以红色的荧光形式消散掉的值[7]。本研究结果显示,突然暴露在光照条件下,桃金娘Kautsky峰值:夏季>秋季>春季>冬季,叶绿色素捕获的光能以红色的荧光形式消散掉的能量损耗:夏季>秋季>春季>冬季。
从植物捕获光能分配上来看,用于植物光合作用称为化学淬灭(Qp),没有用于光合作用而是以热量等形式消散掉称为非化学淬灭(NPQ)[34]。化学淬灭(Qp)越高植物对捕获光能利用率也就越高;非化学淬灭(NPQ)是植物为保护光系统,会把捕获过多的能量以热能的形式消耗掉。本试验研究结果显示,在光稳态下桃金娘Qp为春季>冬季>夏季>秋季,NPQ则为秋季>夏季>冬季>春季,QY为夏季>冬季>春季>秋季;桃金娘经过暗处理后的QY为夏季=冬季>秋季>春季。
植物的光响应曲线对了解植物光化学过程具有重要意义[35]。当植物光系统吸收的光能超过植物被利用能量极限时,植物光合作用会受到抑制,Fv/Fm两者变化是判断植物是否发生光抑制的重要指标[36-37]。本试验研究显示,桃金娘春季和冬季在光照强度为100 μmol/(m2·s)时,QY达到最大值,超过100 μmol/(m2·s)后QY都会随着光照强度增加而减小,夏季和秋季桃金娘QY随着光照强度增加而减小;春季、夏季、秋季的光系统PSII活性(Fv/Fm)的变化并未出现随着光照强度增强而下降的现象,冬季桃金娘在光照强度小于50 μmol/(m2·s)时,光系统PSII活性(Fv/Fm)随着光照强度增加呈递减趋势,当光照强度超过50 μmol/(m2·s)后,随着光照强度增加而增加。这一研究与王晓燕[38]的研究结果不一致。根据众多研究发现高等植物在非胁迫下Fv/Fm稳定在0.83左右[39],桃金娘在光照强度小于500 μmol/(m2·s)时,Fv/Fm<0.83,说明其是否受到光抑制还有待进一步研究。
4 结 论
桃金娘在春季和冬季的“光合午休”由气孔限制引起,夏季桃金娘的“光合午休”属于非气孔限制;不同季节下生理生态因子对光合作用的影响大小不同,桃金娘春季Pn受到Gs、HR以及Ci影响,夏季Pn受到PAR影响,秋季Pn受到Tr、Gs、Ci影响,冬季Pn受到Tr、Ca影响;桃金娘不同季节下光系统PSII活性(Fv/Fm)为夏季>秋季>春季>冬季,ABS/RC为春季>秋季>夏季>冬季,光稳态下QY为夏季>冬季>春季>秋季,最大QY为夏季>冬季>秋季>春季;春季和冬季桃金娘QY在光照强度100 μmol/(m2·s)时最大,夏季和秋季桃金娘QY随着光照强度增加而减小。
猜你喜欢
农业与技术(2021年2期)2021-02-05 10:56:02
当代工人(2020年11期)2020-07-06 16:41:17
中国防伪报道(2018年2期)2018-05-14 14:21:04
现代园艺(2017年19期)2018-01-19 02:49:57
新课程·下旬(2017年7期)2017-08-14 11:12:50
作文大王·中高年级(2016年11期)2016-12-09 20:50:52
学生天地(2016年24期)2016-05-17 05:47:26
学生天地·小学中高年级(2015年9期)2015-11-05 02:04:37
西北林学院学报(2015年5期)2015-01-03 05:12:20
食品研究与开发(2012年3期)2012-12-03 05:44:20
