
土壤-刺槐系统水流阻力对水分胁迫的响应
2022-03-15 05:20武小飞岩晓莹郭天崎朱谧远黄明斌
水土保持研究 2022年2期
武小飞, 杨 帆, 岩晓莹, 郭天崎, 朱谧远, 黄明斌
(1.西北农林科技大学 资源环境学院, 陕西 杨凌 712100;2.西北农林科技大学 水土保持研究所 黄土高原土壤侵蚀与旱地农业国家重点实验室, 陕西 杨凌 712100)
刺槐(RobiniapseudoacaciaL.)是豆科落叶乔木,原产于北美地区。由于其耐旱耐贫瘠、速生、适应性强且固氮效果好等特性被我国北方地区广泛引种[1],用于改善生态环境、防治水土流失,其面积达177.8万hm2[2]。刺槐作为黄土高原主要的造林树种之一,其生长常受到不同程度的水分胁迫。水分胁迫不仅影响植物的生长发育,还会对植物的水力特性产生深远影响,如叶片气孔关闭[3]、木质部水势随水分胁迫的加重呈指数下降[4]、根系水流阻力增大导致根区吸水受限等[5]。
按照Van den Honert模型(1948),在稳态流条件下,水分在土壤-植物-大气连续体系统(Soil-Plant-Atmosphere Continuum,简称SPAC)传输的驱动力是水势差,水分传输受水势差和水流阻力的共同影响。水从土壤到根、茎、叶的液态水流传输是维持植物正常生长的基础[6],从叶到大气以蒸腾为主的汽态传输是植物耗水的主要途径,无论是液态还是汽态水流传输,均受到水流阻力的影响。SPAC系统的水流阻力主要包括土-根、茎-叶、叶-气阻力。邵明安等[7]研究发现,水分从土壤输送到冬小麦根茎,水势降低1.0 MPa,从根茎到叶片降低0.3 MPa,从叶部汽化扩散到大气中降低90.0 MPa。Zhang等[8]研究番茄发现,水势从土壤到根木质部降低0.7~1.1 MPa,从根到叶片降低0.8~1.2 MPa,从叶片到大气降低81.2~166.8 MPa;水势的降低与各部分水流阻力相对应,水流阻力主要存在于从叶片气孔向大气扩散的环节。张喜英[9]对冬小麦(TriticumaestivumL.)和夏玉米(ZeamaysL.)等作物、巩玉霞[10]对小叶锦鸡儿(CaraganamicrophyliaLam.)和杨柴(HedysarummongolicumTurez.)等灌木的研究结论与上述学者的相近,但水流阻力的绝对大小不同。有关水分胁迫对水流阻力影响的研究表明,植物受到轻度水分胁迫时,枝条木质部阻力是植物体水分传输的主要阻力,较强的枝条水流阻力有效地减少枝叶的水分散失[11],使根部输送来的水分均匀分配,保证植物正常的生长发育;当植物遭遇严重的水分胁迫时,土壤水势下降,土壤水分与植物土-根界面的水势差减小[12],根系木质化和栓化,使根系木质部阻力增大[13],此时枝叶部位降低枝条阻力以竞争根系吸收的有限水分[14],缓减水分胁迫对植物的伤害。确定水流阻力存在的主要位置和大小是描述植物对水分胁迫响应的基础,有助于定量研究植物水分传输过程。然而,目前对黄土高原土壤-刺槐系统水流阻力的研究较少。本研究旨在通过探究土壤-刺槐系统水流阻力的分布及时间变化,揭示土壤-刺槐-大气系统水分运移规律,为研究黄土高原刺槐对干旱环境的适应性提供理论依据。
1 材料与方法
1.1 试验概况
试验在西北农林科技大学玻璃温室(34.27°N,108.07°E)进行,温室配备完整的遮阳、控温、补光等设备,满足刺槐植株生长的光温条件。试验用土类型为杨凌土,室内的温度(T)范围为18.0~30.0℃,相对湿度(RH)范围为60.0%~80.0%。土的物理及化学性质见表1。

表1 土的物理及化学性质
1.2 试验设计
试验设置了8个水分梯度,用以评估水分胁迫对土壤-刺槐系统水流阻力的影响。于2019年4月,将两年生的刺槐幼苗修剪后(保证相近的株高及根系分布)移栽至10.8 L容积的塑料盆中(塑料盆上直径27 cm,下直径22 cm,高度31 cm),每盆1株,并于盆栽土表覆盖1.5 kg,0.4~0.8 cm的石英砂以抑制土表蒸发。试验用土为过5 mm筛的杨凌农田土,土壤容重为1.40 g/cm3。于4—5月给所有盆栽充分浇水保持水分含量达到田间持水量,保证刺槐健康生长。随后将所有盆栽随机均分为8组,进行水分处理,即田间持水量的(30±2.5)%,(40±2.5)%,(50±2.5)%,(60±2.5)%,(70±2.5)%,(80±2.5)%,(90±2.5)%和(100±2.5)%,每个水分处理包含刺槐7株。水分处理1个月后测定刺槐生长季内的水力特性,包括7—10月的蒸腾速率、根水势、叶水势等,从而计算水流阻力。试验期间的水分处理通过每2 d称取塑料盆重量完成补(控)水,并把刺槐生物量作为皮重去除(生物量取决于相对株高和叶面积,可用二者模拟计算出植株的生物量)。
1.3 试验内容与方法
1.3.1 土壤物理及化学性质测定 采用Master sizer 2000激光粒度仪(Malvern Instruments, Malvern, England)测定土壤颗粒组成,并根据美国农业部粒级制进行土粒分级。土壤有机碳含量和全氮含量分别采用重铬酸钾外加热法和半微量凯氏定氮法测定。土壤全磷含量采用氢氧化钠熔融-钼锑抗比色法测定。田间持水量参考吴元芝[15]采用威尔科克斯法的测定结果。
1.3.2 蒸腾速率测定 刺槐叶片的蒸腾速率[Tr,mmol/(m2·s)]采用便携式光合仪(Li-6400,LI-COR,USA)测定,于晴天上午09:00—11:00测定冠层向阳、发育良好、小枝中上部的完全展开叶,在光合仪的菜单栏中输入叶面积随即测定Tr。叶面积(LA,cm2)采用经验公式[16]:LA=LL·LW·X/100,X为叶面积回归系数,值为0.7868。使用数显游标卡尺测定叶长(LL)、叶宽(LW,mm)。每个处理选择3株刺槐,每株刺槐选取3片叶子。
1.3.3 叶水势、根水势和土壤水势测定 叶水势(ψl,MPa)的测定采用压力室仪(Plant Moisture Stress, Model 1000, Albany, OR, USA),在晴天的中午,从每个处理的3棵植株上选择冠层部位相似、带有10片以上叶子的刺槐小枝,立即取样并装入盛有湿巾的装样室,多次测定以减少误差。根水势(ψr,MPa)的测定使用小液流法,将刺槐根系从土壤中分离,迅速装入冰盒,带到实验室测定距盆栽土壤表面5 cm以下的根水势。由于根水势测定需要破坏刺槐生长的土壤,故每月的根水势仅从各水分处理的2个盆栽中选取3~4条根系测定。土壤水势(ψs,MPa)通过土壤含水量与水分特征曲线计算得到[15,17]。
1.3.4 根系扫描 使用扫描仪(Cano Scan LIDE 300, Canon, Vietnam)测定根系指标。将刺槐根系连同盆栽土壤装入60目的网袋,用水冲洗至根系表面无明显土壤附着。随即带回实验室,于4℃冰箱中冷藏待测。待所有植株根系洗净,使用根系扫描仪扫取根系,通过WinRHIZO Pro 2012b软件分析获得各水分含量下刺槐根系的生长情况(表2)。

表2 刺槐根系指标
1.4 数据分析
使用Excel 2010进行数据整理及计算均值、标准误差,使用Origin 2018作图,使用SPSS 22进行显著性分析,并通过以下公式计算水流阻力:
1.4.1 土壤阻力 利用Gardner[18]通过对单根稳态流的研究给出的公式计算土壤阻力(Rs):
(1)
式中:K(θ)为土壤导水率(cm/s);Zr为根系深度(cm);L为根长密度(cm/cm3),即单位体积土壤中的根系长度;r为根半径(cm);Rs为土壤阻力(s)。由s变换到(MPa·m2·s)/mmol需乘1.764×10-10。
1.4.2 根系总阻力 根系总阻力(Rr)由土-根界面阻力、根系吸收阻力(径向阻力)和根系内木质部传导阻力(轴向阻力)组成。Rr由水势差与Tr求得:
(2)
式中:ψs为土壤水势(MPa);ψr为植物根水势(MPa);Tr为植物叶片的蒸腾速率[mmol/(m2·s)];Rs为根系总阻力[(MPa·m2·s)/mmol]。
1.4.3 植物传导阻力 植物传导阻力(Rp)包括植物根到茎、叶的传输阻力,计算公式为:
(3)
式中:ψl是植物叶水势(MPa);Rp是植物传导阻力[(MPa·m2·s)/mmol]。
1.4.4 叶-气系统的水流阻力 叶-气系统的水流阻力(Rla)由下列公式计算:
(4)
式中:ψa为大气水势(MPa);Rla为叶-气系统水流阻力[简称叶-气阻力(MPa·m2·s)/mmol]。ψa使用以下公式算出[19]:
ψa=4.6248×105T×ln(RH)
(5)
式中:T为林冠上空气的绝对温度(K);RH为林冠上空气相对湿度(%)。
1.4.5 土壤-植物系统总阻力 根据电模拟原理,SPAC系统中各部分阻力是串联的,故总阻力(Rt)为各分量的代数和,即:
Rt=Rla+Rp+Rr+Rs
(6)
式中:Rt为土壤-植物系统的总阻力[(MPa·m2·s)/mmol]。
2 结果与分析
2.1 水分胁迫对生长季内刺槐蒸腾速率、根水势及叶水势的影响
图1A为不同水分处理下刺槐7—10月的Tr变化情况。各月的叶片Tr随水分含量的增加表现出先增大后维持稳定的趋势。Tr在30%~70%(或80%)的田间持水量范围随水分含量的增加而增大,当水分含量高于80%的田间持水量后Tr基本稳定。水分处理间的Tr存在显著差异,以8月为例,30%,40%~50%与60%~100%田间持水量间存在显著差异。生长季的刺槐叶片Tr于7—9月增大,10月减小。其中,9月和10月的叶片Tr变化趋势相近。
由图1B可知,ψr随着水分含量的增加而增大。各月的ψr在50%的田间持水量处理以下随水分含量的增加而增大;当水分含量高于50%的田间持水量处理后,ψr的增长较少。显著性分析表明水分处理间的ψr存在显著差异,其中8月的ψr在30%,40%,50%和60%~100%田间持水量处理下存在显著差异。从时间尺度上看,在30%~50%的田间持水量范围,ψr于7—9月增大,10月减小;在50~100%的田间持水量范围,ψr于7—10月呈降低趋势。
ψl(图1C)随时间及土壤水分含量的变化趋势与ψr(图1B)相似。水分含量低于田间持水量的50%时,ψl随着水分含量的增加而迅速增大;高于50%后其增长变缓。部分月份的ψl在50%~60%及80%~90%的田间持水量范围有变小的趋势。对ψl进行显著性分析发现,9月各水分处理间的ψl无显著差异,其他月的水分处理间存在显著差异。从时间尺度上看,水分含量低于田间持水量的50%时ψl的大小:9月>10月>8月>7月;高于50%时ψl的大小7月>8月≈9月>10月。

图1 生长季内刺槐蒸腾速率、根水势及叶水势随土壤水分含量的变化情况
2.2 土壤-刺槐系统各部分水流阻力对水分胁迫的响应
2.2.1 土壤阻力 图2 A显示了Rs随水分含量的增加呈现减小的趋势。当水分含量低于50%的田间持水量时,Rs随水分含量的增加而迅速减小;高于50%时Rs随水分含量的变化较小。对Rs进行显著性分析发现,30%与40%~100%的田间持水量范围的Rs存在显著差异,40%~100%水分范围的处理间无差异。
2.2.2 根系总阻力 图2 B为生长季内Rr随土壤水分含量的变化情况。各月的Rr在50%的田间持水量以下随水分含量的增加而迅速减小,其中7月、8月的Rr减小最快;当水分含量高于50%的田间持水量时,Rr下降缓慢,且各水分处理的Rr在月份间的差异很小。显著性分析结果表明,水分处理间的Rr存在显著差异。总体来讲,7月、8月的Rr值较9月、10月的大。
2.2.3 植物传导阻力Rp(图2 C)随土壤水分含量的增加也呈现减小的趋势,其在30%~50%的田间持水量范围内减小的速率快,在50%的田间持水量以上减小速率变缓。Rp的显著性分析结果表明,7月、8月和10月的各水分处理间均存在显著差异,9月的水分处理间无差异。当水分含量低于50%的田间持水量时Rp的大小:7月>8月≈10月>9月;高于50%时Rp的大小7月≈8月>9月≈10月,该范围的Rp差异较小。
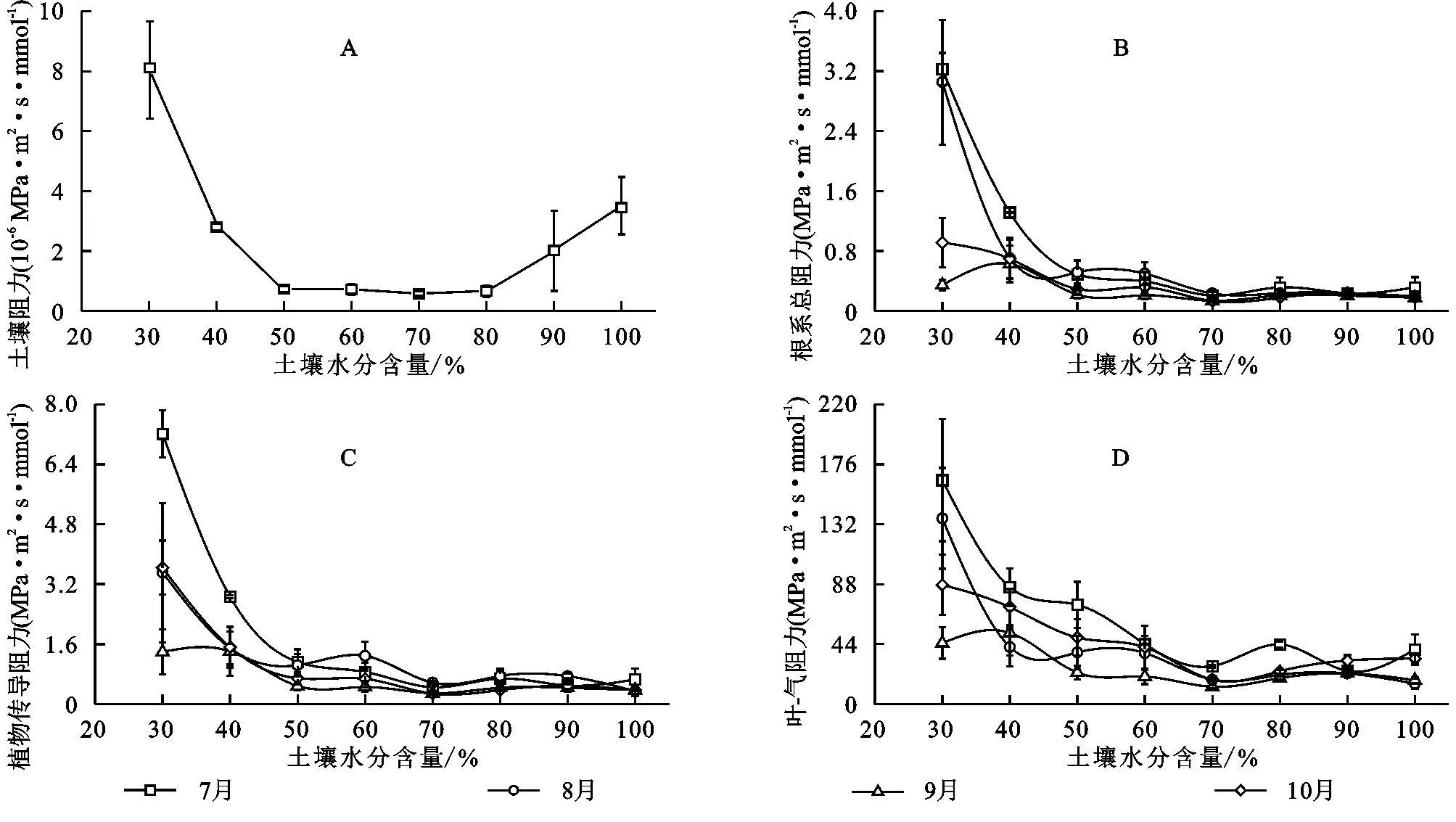
图2 土壤-刺槐系统的土壤阻力、根系总阻力、植物传导阻力及叶-气阻力在不同水分含量及月份间的变化情况
2.2.4 叶-气阻力 图2 D为Rla随土壤水分含量的变化情况。Rla在30%~50%的田间持水量以下随水分含量的增加而迅速减小;在50%~70%的范围减小速率变缓;在70%的田间持水量以上有增大的现象。各月的Rla在水分处理间也存在显著差异。Rla在生长季的变化趋势为:水分含量低于70%的田间持水量时Rla的大小:7月>10月>8月>9月;高于70%时Rla:7月>10月>8月≈9月。
2.3 土壤-刺槐系统总阻力对水分胁迫的响应
图3A表明,Rt在30%~50%的田间持水量范围内随水分含量的增加而迅速减小,在50%~70%的田间持水量范围减小速率变缓,在70%的田间持水量以上呈现的增加现象与Rla相似。值得注意的是,土壤-刺槐系统的Rt随土壤水分含量及时间的变化趋势与Rla基本相同。各月Rt的水分处理间存在显著差异。从时间尺度来看,土壤-刺槐系统的Rt大小:7月>10月>8月>9月。由3B可知,各部分水流阻力占总阻力的百分比于7—10月在波动,但总体遵循着一定的大小比例,即:在土壤-刺槐系统内Rla占比最大,占Rt的96.0%以上;Rp次之,占比1.9%~2.2%;Rr占比0.8%~1.4%;Rs最小,与系统内其他阻力相比小106~108倍,其大小可以忽略,故在图3B中未参与比较。
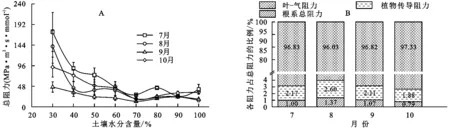
注:B中数字代表相应阻力占总阻力的比例。
3 讨 论
生长在30%~70%田间持水量范围内的刺槐,通过调节气孔导度使叶片Tr随着水分含量的增加而增大,以缓减水分胁迫对植物的不良影响[20];当水分含量高于田间持水量的80%时,刺槐叶片Tr基本稳定,这与以往关于侧柏的研究结果相近[21]。刺槐叶片Tr于7—9月增大、10月减小的变化趋势,是刺槐适应干旱环境的结果[22]。7—9月刺槐处于旺盛的生长时期,叶面积、Tr等随生长时间的增加而迅速增大,刺槐植株快速生长;10月由于气温降低、光照时长等的减少及刺槐固有的落叶习性,使刺槐的生长受限、Tr减小。植物水势可以反映出植物的水分利用状况。植物在遭遇水分胁迫时,根区吸水受限导致植物根直径减小、根轴向萎缩[23],使ψr下降,降低了液流速率,从而引起ψl的下降[24]。刺槐的ψr和ψl随着水分条件的改善均呈现增大的趋势,二者在30%~50%的田间持水量范围增加最快,且7—10月刺槐的水势逐渐降低,降低的植物水势增大了水势差,有助于刺槐根系吸水及植物传输水[6]。
水流阻力受水分胁迫的影响,水从土壤进入植物根系,经茎、叶扩散到大气的全过程贯穿着土壤-植物系统各部分的阻力。水分胁迫导致植物根系收缩,土-根界面空隙变大,Rs增大[25]。邵明安等[7]认为水分含量低于田间持水量的40%时Rs较大,否则作用甚微,这与本研究仅30%田间持水量下的Rs与40%~100%田间持水量之间存在显著差异相符。由于土壤水分含量增加使刺槐的Tr和ψr均增大,故Rr随水分含量的增加而减小。其中7月、8月的Rr值更大的原因可能是该阶段叶片处于生长发育前期,叶面积较小、蒸腾作用相对较弱。Rp与Rr的变化趋势相近,是因为水分在植物内部传输过程中阻力变化较小且变化趋势相近[26]。Rla与水分含量间也呈负相关关系,这是ψl和Tr共同作用的结果[1]。Rla在70%的田间持水量以上有较小的增大趋势是因为植物生长差异引起了该水分范围的叶水势(图1C)降低。由于Rla占Rt的96.0%以上,故Rt与Rla呈相似的变化趋势,以往关于葡萄、玉米、小叶锦鸡儿等的研究均表明Rla是土壤-植物系统“耗能”最多的阻力部分,该过程主导了植物被动吸水的过程,引发了水由液态—气态的相变[10]。整体来讲,7—9月水流阻力逐渐减小,10月增大,其大小为:7月>10月>8月>9月。生长季内的水流阻力随水分含量的增加而减小,是刺槐调节自身与土壤水分间的关系、促进生长发育的结果。
根据刺槐生长季内Tr的变化趋势,我们认为刺槐蒸腾耗水的水分利用阈值为75%的田间持水量[15],大于该水分含量刺槐Tr处于最大值,小于该水分含量Tr随含水量的减少而降低。生长季内刺槐对水分的利用状况与植株从土壤中可获得的水量相关[27]。黄土高原降水少、蒸发量大、年际间降雨不均等使土壤蓄水不足,导致刺槐生长受水分胁迫的影响[28],但该地区的降雨多集中在刺槐生长季,一定程度上缓减了水分胁迫对植株带来的伤害[29],使刺槐得以生存。以往的研究结果表明,黄土高原土的年平均水分含量为田间持水量的50%~59%[28,30],结合本研究Tr,ψr,ψl和水流阻力等的变化、分布趋势,我们推测黄土高原刺槐林地合理的水分范围为50%的田间持水量以上。该水分范围使土壤-刺槐系统拥有相对高的Tr,低的水流阻力,以适应黄土高原干旱缺水的土壤环境,保证刺槐林可持续生长。
4 结 论
(1) 刺槐的Tr在30%~70%(或80%)的田间持水量范围,随着土壤水分含量的增加而增大,在80%的田间持水量以上基本保持稳定,生长季内的变化趋势为7—9月增大,10月减小;ψr和ψl在30%~50%的田间持水量范围均随水分含量的增加而迅速增大,此后变化较小,7—10月的ψr和ψl呈减小趋势;
(2) 水流阻力随着水分含量的增加而减小。Rs,Rr和Rp在30%~50%的田间持水量范围随着水分含量的增加迅速减小,在50%的田间持水量以上的变化较小;Rla在30%~50%的田间持水量范围减小的速率最快,在50%~70%的范围减小速率变缓,在70%的田间持水量以上有增大的现象。Rt的变化趋势与Rla相近,是因为Rla占Rt的96.0%以上,主导了总阻力的变化。水流阻力于7—9月减小,10月增大;
(3) 根据的刺槐Tr,ψr,ψl和水流阻力等的分布及变化趋势,我们推测生长在大于50%田间持水量范围的刺槐可以保证相对高的Tr、低的水流阻力,以适应黄土高原干旱缺水的土壤环境,促进刺槐林可持续生长。
猜你喜欢
中学生数理化·八年级物理人教版(2022年4期)2022-04-26
干旱地区农业研究(2021年6期)2021-12-08
中学生数理化·八年级物理人教版(2021年11期)2021-12-06
现代园艺(2020年2期)2020-12-22
农村实用技术(2020年9期)2020-11-19
文苑(2020年6期)2020-06-22
农民致富之友(2019年26期)2019-10-11
湖北林业科技(2019年4期)2019-09-16
小溪流(故事作文)(2018年10期)2018-12-28
农业与技术(2017年22期)2017-12-29
