
越南、缅甸茶树种质资源遗传多样性ISSR分析
2022-03-12 05:57苏鸿锋晏嫦妤
广东茶业 2022年1期
苏鸿锋 晏嫦妤
(华南农业大学园艺学院,广东 广州 510642)
越南拥有丰富的茶树资源,大致分为小叶茶(C.sinensis(L.)O.Kuntze var. sinensis)、阿萨姆茶(C.sinensis var. assamica(Masters))和掸茶(C.sinensis var. pubilimba Chang),其中小叶茶品种占比最大,掸茶品种占比最少[1]。缅甸茶树多为乔木和小乔木的大叶品种,仅在北部地区混有叶形较小的茶树,其主要的茶树品种类型包括掸部种(T.S. Burma and shan)、云南大叶种(T.S. yunan)、葡萄种(T.S. Putao)、阿萨姆种以及一些野生型茶树[2]。但是关于越南、缅甸茶树种质资源遗传背景的研究较少。本研究采用ISSR分子标记技术,对华南农业大学茶树资源圃中的17份越南、缅甸茶树种质资源进行遗传多样性分析,以期了解这些资源的遗传背景,为进一步开发利用提供依据。
1 材料与方法
1.1 材料
材料来源于华南农业大学校内茶园,摘取茶树单株嫩叶用于提取基因组DNA(表1)。
1.2 实验试剂与设备
实验试剂包括快捷型植物基因组DNA提取试剂盒(DP321) 、2×Taq PCR Mastermix(KT201)、GeneGreen核酸染料(RT210)、D2000 DNA Marker(MD114) 、70%乙醇溶液(v/v)和异丙醇溶液等。
主要仪器设备包括离心机、WiseBath恒温水浴锅、BG-Power600型电泳仪、SQ510C立式压力蒸汽灭菌锅、超低温冰箱、Link-Bio PCR热循环仪、Link-Bio超微量核酸蛋白分析仪、伯乐GelDoc XR+凝胶成像仪、IM-25制冰机、鼓风烘干箱等。
1.3 方法
1.3.1 茶树DNA的提取
利用由天根生化科技(北京)有限公司提供的快捷型植物基因组DNA提取试剂盒(DP321)对17份茶树材料DNA进行提取,并及时放入-4℃冰箱中保存备用。
1.3.2 引物的筛选与PCR扩增
经筛选,选用引物UBC826、UBC830、UBC849、UBC873、UBC880,引物序列见表2。PCR反应体系和PCR扩增条件分别见表3和表4。

表2 引物序列信息

表3 ISSR-PCR反应体系

表4 ISSR-PCR反应程序
1.3.3 数据处理与分析
采取人工读带的方法,在琼脂糖凝胶成像系统拍照后,读取琼脂糖凝胶上出现的条带,将在相同位置的地方出现的清晰可辨的条带记作“1”,相同位置的地方没有出现条带或无法辨析的条带记作“0”,并把五条引物扩增出来的所有条带的数据输入到Excel表格,计算出ISSR引物的多态性百分比。使用NTSYSpc2.10、Popgene32等软件进行遗传多样性分析。
2 结果与分析
2.1 引物扩增多样性分析
图1和图2为部分扩增结果,使用软件Popgene32对5条ISSR引物的扩增结果进行分析表明(表5),5条引物共扩增出45个位点,多态性位点38个,总多态性百分率为84.44%,平均多态性位点7.6个。其平均Nei’s基因多样性指数(H)为0.28,平均Shannon信息指数(I)为0.44。说明该17份茶树供试材料具有丰富的遗传多样性,可以作为育种研究的材料。
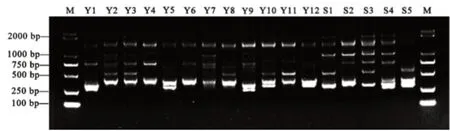
图1 引物UBC826的扩增结果
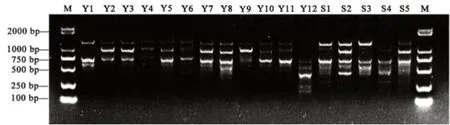
图2 引物UBC826的扩增结果
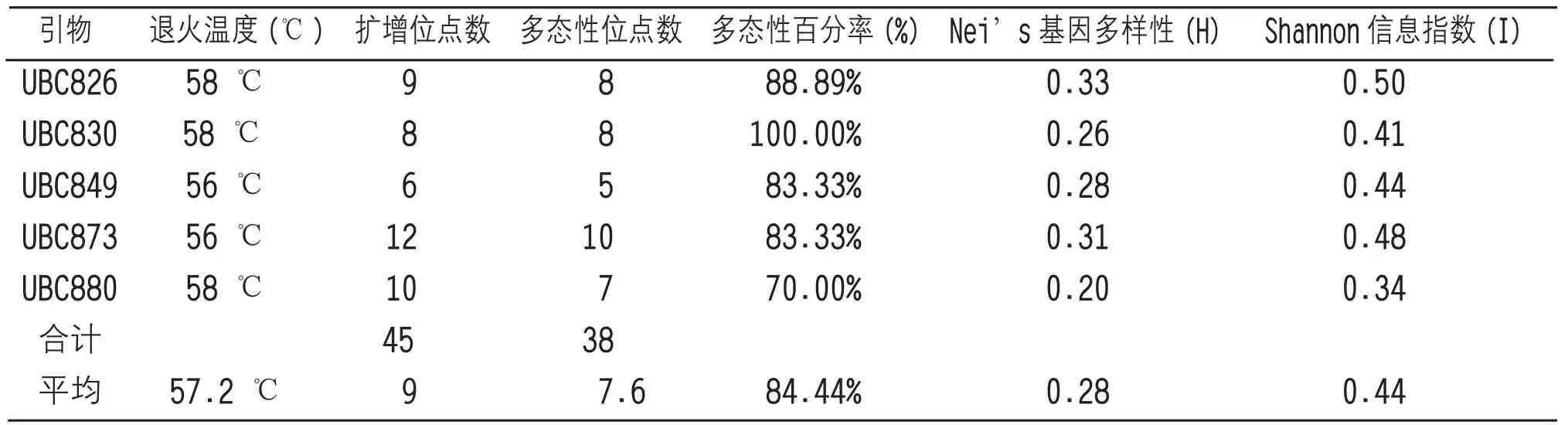
表5 引物扩增结果及多态性
2.2 遗传相似性分析
17份材料的遗传相似性系数与遗传距离结果如表6所示,遗传相似系数的变化范围在0.489~0.867之间,平均遗传相似系数为0.704,其遗传距离在0.143~0.716之间,平均遗传距离为0.358,说明其个体间存在较大的变异。其中编号为Y2和Y3、Y6和Y8的样本遗传相似性系数最高,为0.867;编号为S4和Y12的样本遗传相似性系数最低,为0.489。将17份茶树资源按品种划分为越南大叶种和缅甸掸部种后进行居群间分析得,总遗传多样性(HT)为0.2715,种群内基因多样性(HS)为0.2369,种群间基因多样性(DST)为0.0346,遗传分化水平(Gst)为0.1277,基因流为3.4163,两个居群间的遗传相似性为0.0949,说明该两个品种间存在中等程度的遗传分化,种群内基因交流较为频繁,而种群间的遗传交流较低。
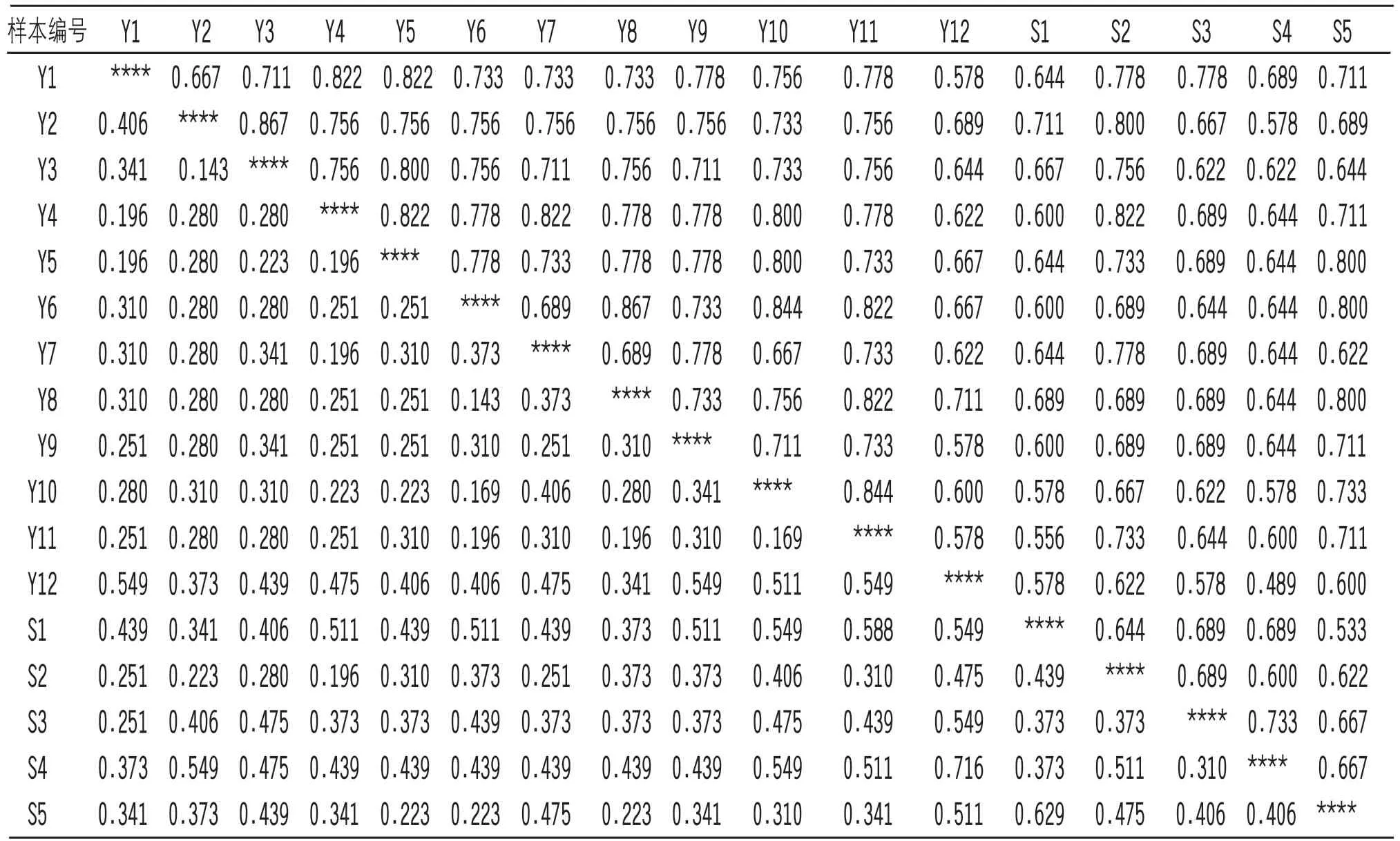
表6 供试材料的遗传相似系数与遗传距离
2.3 聚类分析
使用软件NTSYSpc2.10分析17份材料种质资源的原始二元矩阵数据,按UPGMA法(Unweighted Pair-Group Method with Arithmetic Mean)对供试材料种质资源进行聚类分析(图3)。结果显示,在遗传相似系数为0.61处,17份供试材料只有编号为Y12的茶树样本被单独划分出来。在遗传相似性GS为0.68处,17份茶树资源被划分为3大类,类群Ⅰ包括13份茶树资源,其中有11份越南茶树资源和2份缅甸茶树资源、类群Ⅱ包括3份缅甸茶树资源、类群Ⅲ包括1份越南茶树资源(Y12)。
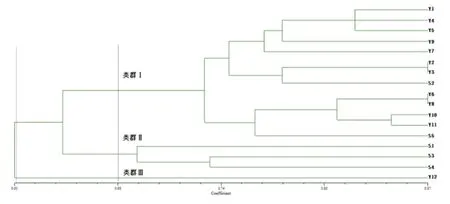
图3 17份茶树种质资源的聚类分析
3 讨论与结论
3.1 17份茶树种质资源的遗传多样性分析
本研究结果说明供试材料的茶树遗传多样性丰富,个体间变异程度较大,可以作为育种研究中的优良材料。茶树Y2与Y3、Y6与Y8具有最高的遗传相似度(0.867),证明它们有比较一致的遗传基础;茶树Y12与S4的遗传相似程度最低(0.489),变异丰富较大。越南大叶种的遗传相似度(0.756)大于掸部种的遗传相似度(0.667),因此掸部种茶树的变异程度大于越南大叶种茶树,掸部种茶树具有更丰富的遗传多样性。茶树Y12在遗传相似度为0.61处被单独划分出来,证明其变异最大,可以作为日后特异种质进行进一步研究。缅甸掸部种茶树S2和S5被聚类到类群Ⅰ(越南大叶种群体),其原因有待进一步分析。
3.2 与不同地区与国家的茶树资源遗传多样性比较
基于ISSR分子技术(表7),朱晨等[3]利用该法研究53份福建茶树资源,结果表明其多态性位点占比为94.84%,基因多样性指数为0.224,Shannon信息指数为0.368,遗传相似系数为0.895。孙雪梅等[4]利用该法研究云南67份茶树资源,结果表明其多态性位点占比为97.76%,基因多样性指数为0.27,Shannon信息指数为0.42,遗传相似系数为0.693。高树文[5]利用该法研究28份山东茶区资源,结果表明其多态性位点占比为86.0%,遗传相似系数为0.693。李浩宇等[6]对苏州洞庭地区的51份茶树资源进行分析,结果表明其多态性位点占比为92.11%,遗传相似系数介于0.37~0.89。李建华等[7]对四川地区37份茶树资源进行分析,结果表明其多态性位点占比为95.2%,遗传相似系数介于0.689。除此之外,周治淼等[8]利用SRAP分子标记对广东地区25份茶树资源进行分析,结果表明其多态性基因位点占比88.67%,遗传相似系数为0.3195。Shen CW等[9]利用RAPD分子标记分析湖南240份茶树资源,结果表明其多态性位点占比为88.9%,遗传相似系数0.57[9]。乔小燕等[10]利用EST-SSR技术分析广东、广西共105份茶树资源,其中广西茶树资源的多态性信息含量(PIC)为0.41,地区间遗传相似系数在0.83~0.94之间;李烨昕[11]利用SSR技术对安徽65份茶树资源,结果表明其Nei’s基因多样性指数为0.4751,Shannon信息指数为0.7869,遗传相似系数在0.946~0.987之间。曹烁[12]基于SNP(单核苷酸多态性)分子标记分析贵州地区51份茶树资源,结果显示其基因多样性为0.472,Shannon信息指数为0.66,遗传相似系数为0.570。Xu YX等[13]基于SCoT(目标起始密码子多态性)分子标记对浙江18份茶树品种资源进行分析,结果表明其多态性位点百分比为83.7%,遗传相似系数在0.587~0.814之间。
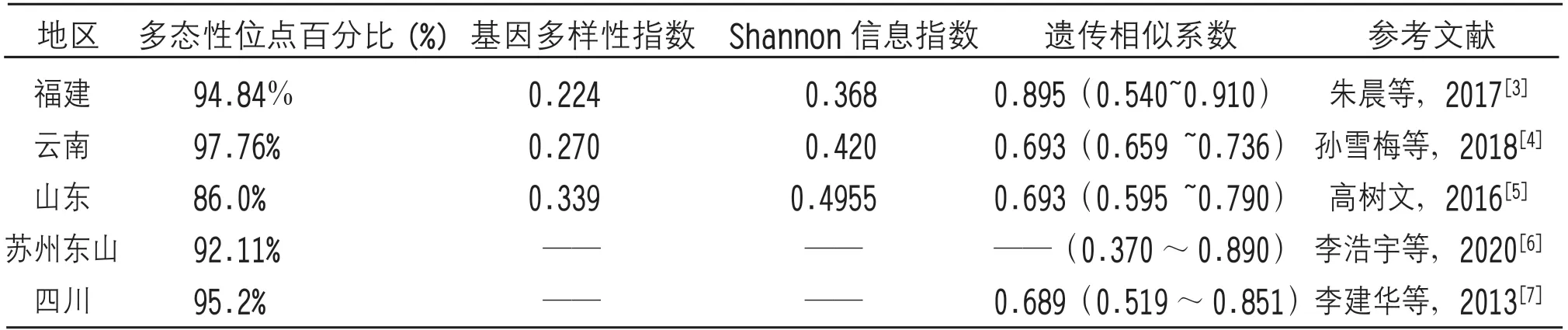
表7 基于ISSR标记的不同地区茶树种质资源的遗传多样性分析数据
由以上数据分析表明,17份茶树材料的多态性百分比小于福建、云南、四川等茶树资源的多态性百分比,接近于广东、湖南、浙江等地区茶树资源;其基因多样性指数接近云南和福建,Shannon信息指数接近云南和湖南;其遗传相似性小于福建、安徽和广西等茶树资源,高于广东、湖南、贵州等地,接近于云南、山东、四川、贵州等地茶树资源的遗传相似系数。总之,17份茶树材料遗传多样性情况与云南茶树资源基本一致。
不同的国家中,中国、日本和肯尼亚三个国家之间茶树资源基因多样性,中国最为丰富,其多态性位点百分比(PPL)为96.1%、Nei’s基因多样性(H)为0.22、Shannon信息指数(I)为0.35[14],与本研究的茶树资源遗传多样性接近。Francis Wachira等[15]研究结果得:越南茶树资源的基因多样性为0.260,Shannon信息指数为0.389,低于本实验数据,并且其遗传相似性更接近印度、斯里兰卡等国。从Yoshinobu Katoh等学者[16]的研究可知,缅甸的茶树资源与滇缅茶(C. irrawadiensis)具有遗传相似性,而越南茶树资源与泰国茶树在形态分类上有密切关联。17份茶树资源遗传多样性接近中国、印度、泰国等南亚和东南亚国家的茶树资源,遗传多样性高于日本、肯尼亚等国。李远志等[17]研究认为越南大叶种、缅甸的掸部种与中国的云南大叶种在遗传上具有相似性,其原因可能是云南大叶种通过与越南和缅甸部分相连的河流,如怒江、龙川江等逐渐流到越南和缅甸后形成的两个茶树品种。
虽然ISSR分子标记应用广泛,但在研究茶树育种的时候,选取何种分子标记进行研究有待进一步探索。它取决于一系列因素,包括引物的分子性质、研究对象生物学特性和可以利用的资源等[18]。在Kumar Mondal[19]研究的认为,ISSR引物比SSR引物更容易获得,可以作为RAPD分子标记技术的一种补充手段使分析结果更加可靠,但ISSR标记不是共显性标记,可能会导致有效的双等位基因条带消失等缺点。目前有较多研究数据与本次研究数据进行比较,不过由于采取的分子标记手段有所不同,对于其分析结果可能有所差异。Zhang Y等[20]通过SRAP、SCoT、EST-SSR三种分子标记研究秦巴地区茶树资源,发现它们产生的遗传距离相关性很小,因此不建议用单一分子标记的手段评价遗传多样性和群体结构,而应该利用不同类型分子标记的结合进行综合评价。所以,对于不同地区的茶树资源、不同的茶树品种和不同的茶树群体,采取何种手段进行研究,还需要进行进一步的探究。本研究基于ISSR分子标记手段研究越南和缅甸茶树品种的遗传多样性,日后要在此基础上进行更多的探究,才能科学有效分析遗传多样性,合理利用茶树种质资源。
猜你喜欢
分子催化(2022年1期)2022-11-02
世界科学技术-中医药现代化(2022年3期)2022-08-22
茶道(2022年3期)2022-04-27
昆明医科大学学报(2022年2期)2022-03-29
昆明医科大学学报(2021年6期)2021-07-31
昆明医科大学学报(2021年3期)2021-07-22
烟草科技(2021年6期)2021-06-24
福建茶叶(2020年5期)2020-12-23
福建茶叶(2020年10期)2020-12-22
农技服务(2020年1期)2020-12-17
