
Elevation gradient distribution of indices of tree population in a montane forest:The role of leaf traits and the environment
2022-03-08 02:19:00ZuhuaWangRongZhengLilinYangTinghongTanHaiboLiMinLiu
Forest Ecosystems 2022年1期
Zuhua Wang ,Rong Zheng ,Lilin Yang ,Tinghong Tan ,Haibo Li ,Min Liu,*
a College of A&F Engineering and Planning,Tongren University,Tongren 554300,Guizhou,China
b Guizhou Provincial Key Laboratory for Biodiversity Conservation and Utilization in the Fanjing Mountain Region,Tongren University,Tongren 554300,Guizhou,China
c College of Life and Technology,Inner Mongolia Normal University,Hohhot 010022,Inner Mongolia,China
d National Nature Reserve Administration of Fanjing Mountain,Tongren 554400,Guizhou,China
Keywords:Elevation gradient Species distribution Leaf traits Climate change Leaf economic spectrum Mt.Fanjingshan
ABSTRACT Background: To disentangle the controls on species distribution in the context of climate change is a central element in proposed strategies to maintain species diversity.However,previous studies have focused mainly on the roles of abiotic factors(e.g.,climate and soil properties),with much less attention given to the roles of biotic factors such as functional traits.Here,we measured eight leaf traits for 240 individual trees of 53 species and analyzed the variation in traits and population composition indices and their relationships with soil properties,climate factors,and leaf traits.Results:The tree density,frequency and species importance values of the overall species and saplings significantly increased with increasing elevation,while the same indices (except for species frequency) of adults did not significantly change.The largest percentage of variation of species importance value (greater than 50%) was explained by climate,but leaf traits played a critical role in driving elevation distribution patterns of both saplings and adults;the abundance of saplings significantly increased with elevation,with increased leaf carbon contents,while the abundance of adults did not change in accordance with a nutrient conservation strategy associated with the leaf economic spectrum.Conclusions: Our results suggest that the elevation gradient distribution of woody plant species is dependent on tree size and that local atmospheric humidity and leaf traits cause considerable variation in species distribution along subtropical mountain elevations.We provide evidence of which leaf traits play a key role in the elevation gradient distribution of different sizes of woody tree species.
1.Introduction
Global change has a crucial effect on species distribution (Batllori et al.,2020;Chen et al.,2011)and impacts the functions and services of forest ecosystems (Batllori et al.,2020;Wieczynski et al.,2019),which are closely related to human well-being(Wang et al.,2021).Nonetheless,the response of tree species distributions to climate warming vary(O'Sullivan et al.,2020;Wason and Dovciak,2017) because species distributions are often affected by abiotic and biotic factors.However,previous studies have focused mainly on the roles of abiotic factors(e.g.,climate and soil properties),with much less attention given to the roles of biotic factors such as functional traits (O'Sullivan et al.,2020).Plant functional traits are directly linked to the performance of organisms in the environment (Bruelheide et al.,2018),and the consistency and prevalence of many trait-climate correlations over space suggest that they might be good predictors of species distribution responses to climate change(Ahrens et al.,2020;Kemppinen et al.,2021).Consequently,it is important to disentangle the relative contributions of climate and functional traits to species distributions,which can help guide forest management practices and inform decision-making in the public sector.
With climate warming,the distribution of species has shifted upward toward montane areas where temperatures and humidity are suitable for growth (Steinbauer et al.,2018).Consequently,the number of trees is likely to increase in cold and moist regions.Recently,several studies suggested this idea and found that the highest tree densities were present in forested regions with limited temperature and moisture (Crowther et al.,2015),while in contrast,tree density generally increased with temperature and moisture availability (Crowther et al.,2015).These inconsistent results may be related to alternative resource availability.For instance,in colder (boreal or tundra) biomes,increased moisture levels can lead to hydric and permafrost conditions in low-lying areas,which then limit nutrient availability for tree development (Bonan and Shugart,1989).Additionally,the inconsistent results may be related to tree size.Compared with the distribution of adults,the distribution of young trees is likely to be more closely related to the current climate,which may lag behind recent climate change(Liu et al.,2020;Martin and Thomas,2013).Thus,young trees should advance to higher latitudes or elevations with climate warming,whereas adults may not exhibit this apparent tendency along the elevation gradient.However,it is still unclear how climate factors and soil properties affect the elevation distribution of species for each tree size.
Leaf traits,which are closely related to tree performance(Reich and Cornelissen,2014),species coexistence (Perez-Ramos et al.,2019) and species distribution(Vesk et al.,2020),are the most sensitive tree organs to climate change(Ahrens et al.,2020).Consequently,leaf traits may be good predictors of species distribution responses to climate change(Soudzilovskaia et al.,2013).For example,species with conservative traits(i.e.,a relatively small leaf size,specific leaf area(SLA),Nmass)are generally favoured in hot,dry climates as well as water-limited conditions(Wright et al.,2004;Wieczynski et al.,2019).Furthermore,drought tolerance has been shown to increase under hotter,drier conditions(Blackman et al.,2017;Asao et al.,2020).Compared with those without conservative traits,species with conservative traits have a slower growth rate and need less leaf N and leaf P to maintain the tissue growth(Adler et al.,2014).Therefore,from the perspective of leaf morphological traits and chemical traits,disentangling the effects of climate change on species distribution may be a suitable approach.However,accumulating evidence has proven that leaf traits significantly differ between saplings and adults (Hietz et al.,2017);thus,the effects of leaf traits on species distribution may be dependent on tree size.Despite this,there is a knowledge gap concerning the relationships between leaf traits and species distribution at different tree life stages(i.e.,saplings,adults).
Elevation gradients provide natural climate variation in which key environmental factors that affect plant growth and tree distributions,including atmospheric temperature (AT),atmospheric humidity (AH),and their associated soil properties (e.g.,soil pH,soil temperature,soil depth),change considerably within a confined area (Gong et al.,2020;Sundqvist et al.,2013).Along these gradients,plants also undergo changes (e.g.,changes in leaf traits) to adapt to the associated climate conditions (Ahrens et al.,2020;Parmesan and Hanley,2015;Soudzilovskaia et al.,2013).Thus,elevation gradients constitute an ideal platform for studying tree distribution responses to global climate change(Gornish and Tylianakis,2013;Sundqvist et al.,2013).The present study was conducted in a species-rich subtropical montane forest in southern China,where significant changes in elevation(from 600 to 2300 m a.s.l.)occur.To explore the elevation patterns concerning species distributions and their potential drivers,we used a climate dataset and collected information about various leaf traits (see Table 1) along an elevationgradient to answer the following main objectives:1) determine the elevation gradient distribution of overall species,saplings,and adults;and 2) determine the relatively important roles of climate factors,soil properties and leaf traits on the elevation gradient distribution of the overall species,saplings,and adults.

Table 1 Functional traits and their categories,units,and abbreviations.
2.Methods and materials
2.1.Study site and design
The present study was conducted on Mt.Fanjingshan,which is located in the northeastern part of Guizhou Province,Southwest China(27.78-28.02°N,108.60-108.81°E).This region features a humid subtropical monsoon climate with a mean annual temperature (MAT) of 5.0-17.0°C,and the mean annual precipitation(MAP)is approximately 1100-3000 mm.The elevational range of Mt.Fanjingshan is>2000 m,and thus,it has a relatively complete vertical vegetation gradient.On Mt.Fanjingshan,4,394 plant species have been recorded.The vegetation vertical zoning is divided into five zones:evergreen broad-leaved forest,evergreen deciduous broad-leaved mixed forest,Zhongshan deciduous broad-leaved forest,subalpine coniferous forest,and an alpine shrub meadow (Fig.S5).The soils are mainly mountainous yellow soil or yellow-brown soil (Dystric Cambisol soils according to the FAO classification system),with a silty loam texture(Peters,1992).
On the mountain,we selected a relatively uniform,smooth slope(one side only that lacks large ridges and valleys) and established sampling sites every 300 m in elevation starting from 600 m to 2,300 m,resulting in six sites.Each site consisted of 10 sampling points located every 15 m along a transect parallel to the elevation contour (Fig.S6).At each sampling point,we used the point-centred-quarter method (Cottam and Curtis,1956)to quantify the tree density,tree basal area,species relative frequency,and species importance value of each size class (overall,saplings and adults)by species for all trees>4 cm in diameter at breast height (DBH) following the methods of similar studies (Holway et al.,1969).The criterion for overall (DBH>4 cm),saplings (4 cm ≤DBH≤10.2 cm) and adults (DBH>10.2 cm) were based on the previous study(Wason and Dovciak,2017).
2.2.Leaf sampling and trait measurement
Sampling was conducted between July and August 2019.We selected one terminal,fully expanded,sun-exposed branch from the current growing season from each individual tree.The branches were cut using a 5.6 m telescoping pole (ARS Corp.,Senboku,Japan),labelled,and then placed in a cooler for transport to the laboratory.In the laboratory,the branches were placed in water to minimize leaf dehydration,and 20 leaves were selected from each branch for to measure the SLA.These leaves were separately laid flat and imaged together with a reference square(4 cm2)using a Nikon Coolpix 5000 digital camera(Nikon Corp.,Tokyo,Japan),and the total projected leaf area was calculated using the image-processing software ImageJ(Systat Software Inc.,Richmond,CA).The leaves were then dried in a forced-air oven at 70°C for 72 h and subsequently weighed with a scale with a precision of 0.0001 g determine the SLA.The SLA was calculated as the total leaf surface per dry mass (cm2•g-1).The dried leaves were ultimately ground to a fine powder using a coffee mill and stored at 4°C until chemical analyses.
The molybdate/ascorbic acid method was applied to measure the total P concentrations in the leaves after H2SO4-H2O2digestion (Jones,2001).The leaf N and C concentrations were determined by a CHN combustion analyser (TruSpec Micro Analyzer;LECO Corp.,St.Joseph,MO).The P concentration was expressed on a leaf dry mass basis(mg•g-1),and the C and N concentrations were expressed as percentages(%).Finally,we calculated the C:N and N:P ratios to assess plant N and P limitations.
2.3.Soil properties and environmental factors
Taking the target tree (determined by the point-centred-quarter sampling method) as the centre point,a 1 × 1 m2subplot was established to measure the rock coverage (%),litter coverage (%) and litter depth (cm).The soil depth (cm),humidity,temperature and pH were measured at three random sampling points in each subplot.Fresh soil samples were taken from each sampling point,placed in a sealed bag,and refrigerated.The soil was brought back to the laboratory within 24 h and divided into two parts,one part was considered a fresh soil sample and the other part was used for measuring the soil total carbon,total N,and total P after air drying.
We have arranged a meteorological observation station on each of the 6 elevational gradients.Consequently,the AT,AH and pressure data at each elevation gradient were retrieved from the data of the Mt.Fanjingshan Meteorological Station.The average daily AT,AH and atmospheric pressure(AP)were calculated according to the data.
2.4.Data analysis
The elevation patterns of tree density,tree basal area,species relative frequency,and species importance value for each size class (overall,saplings and adults)were assessed by linear mixed effects models,simple linear models,and polynomial regression models.The best-fit regression was selected according to the corrected Akaike information criterion(AICc).Linear mixed effects models (LMMs) were constructed with elevation as a fixed factor and species as a random factor.The normality,homogeneity and independence of the model residuals were checked(Zuur et al.,2009),and the datasets were log transformed if necessary.
To determine the influence of the broad predictor categories(climate,soil,and leaf traits)on the elevation distributions of the size class of each species (the second objective of our study),we constructed multiple LMMs for the species importance value to test the effects of climate,soil,and leaf traits on each species size class.First,to address collinearity of the predictors,we used principal component analysis(PCA)to determine the covariation between leaf traits (SLA,LDMC,LC,LN,LP,LCP,LNP,LCN),and the first two principal component scores represented the leaf traits in the models.Moreover,relationships between the soil properties and climate factors were tested using Spearman correlation analysis,and the factors were discarded when the coefficients were >0.60.Consequently,the predictors of the multiple regression models were AH,AT,soil humidity,soil depth,and PC1 and PC2 scores.In constructing the models,we also included quadratic terms for the abiotic variables(that is,AT,AH,soil depth and soil humidity),as shown in previous studies(Garcia-Palacios et al.,2018;Sirami et al.,2019).To account for the nested structure of our sampling design(several sampling points nested within each elevation),all the models included a random intercept for elevation.
We used a model selection procedure based on the AICc(ΔAICc ≤2)to select the best predictors of the species importance value.This procedure was performed using the ‘dredge’ function in the R package MuMIn.Model averaging was performed based on AICc weights when multiple models were selected.The model residuals were inspected for normality,homogeneity and independence (Zuur et al.,2009),and the predictors were log transformed when necessary.All the predictor variables were standardized before analyses using the Z-score to interpret parameter estimates on a comparable scale.
We also evaluated the relative importance of the predictors as drivers of the species importance value.This method is similar to the approach of a variance decomposition analysis because we transformed all predictors to Z-scores before analysis.The following three identifiable variance fractions were evaluated:(1) climate,(2) soil properties,and (3) leaf traits.
All the calculations were performed with R software(v 4.0.4,R Core Team,2021),and significant effects were detected at the level ofP<0.05.
3.Results
3.1.Variation in leaf traits and soil properties along an elevation gradient
PCA showed that covariation among leaf traits could be represented by two independent dimensions,which accounted for 75%of all leaf trait variation (Fig.2,Table S1).The first principal component (PC1) represented the variation along the leaf economic spectrum(LES),with 59%of the total variation explained (Fig.2).The PC1 scores were negatively associated with LCP,LNP and LCN and positively associated with SLA,LN,and LP (Fig.2,Table S1).The second principal component (PC2)explained 17% of the overall variance and represented the variation along the leaf carbon content;PC2 was negatively associated with LC and LDMC(Fig.2,Table S1).
Soil temperature,soil rock exposure rate,soil pH,AT and AP were significantly negatively correlated with elevation (Fig.S3,P<0.05),whereas AH,litter depth and soil humidity were significantly positively correlated with elevation(Fig.S3,P<0.05).
With increasing PC1 scores,soil temperature,AP and elevation significantly decreased,while soil humidity significantly increased(Fig.S4 a-d).As the PC2 score increased,the elevation decreased significantly(Fig.S4 e).
3.2.Elevation distribution of woody species at each life stage
As elevation increased,tree density,species frequency,and species importance values of overall species increased significantly;when plotted,the trend exhibited a U-shaped pattern,with abundance at the bottom at 1000-1500 m a.s.l.(Fig.1 a,d,j),while the basal area of trees did not significantly change (Fig.1 g).There was an obvious difference between the saplings and adults in response to the elevation gradient.However,with increasing elevation,the four indices of the saplings increased significantly,the trends of which exhibited U-shaped patterns(Fig.1 b,e,h,k),while the same indices of the adults did not change significantly (Fig.1 c,k,m),with the exception that species frequency decreased significantly(Fig.1 f).
3.3.Predictors of species importance values at each tree size class
When using a selected threshold of ΔAICc ≤2,we identified four alternative models for all species (SI Appendix,Table S2) explaining a high proportion of the variance in species importance values(R2=0.54).Climate,soil,and leaf traits were responsible for 83.7%,4.7%and 11.6%of the explained variance in species importance values,respectively(Fig.3).Soil humidity and PC2 did not impact the species importance values of the overall species (Fig.2).However,PC1 was significantly negatively linked to species importance values (Fig.2),suggesting that species with conservative foliar traits had significantly higher species abundances than did species with without conservative foliar traits(Fig.S1).AH had a significant positive effect on species importance values (Fig.2),and AT had a negative impact on species importance values (Fig.2).AH also exhibited a quadratic effect on this variable(Fig.2).
When using a ΔAICc of ≤2,we identified only two alternative models for saplings(SI Appendix,Table S2)explaining a high proportion of the variance in ecosystem stability(R2=0.63).Climate,soil,and leaf traits were responsible for 64.4%,1.9%and 33.7%of the explained variance in species importance values,respectively (Fig.3).Soil,climate and PC1 had no significant effects on species importance value,but PC2 was significantly correlated with sapling importance value (Fig.2),suggesting that,compared with species with a low abundance,those with a high abundance had a higher leaf carbon content.Soil and climate had no significant effects on species importance value,but PC1 was significantly negatively correlated with adult species importance value(Fig.2).
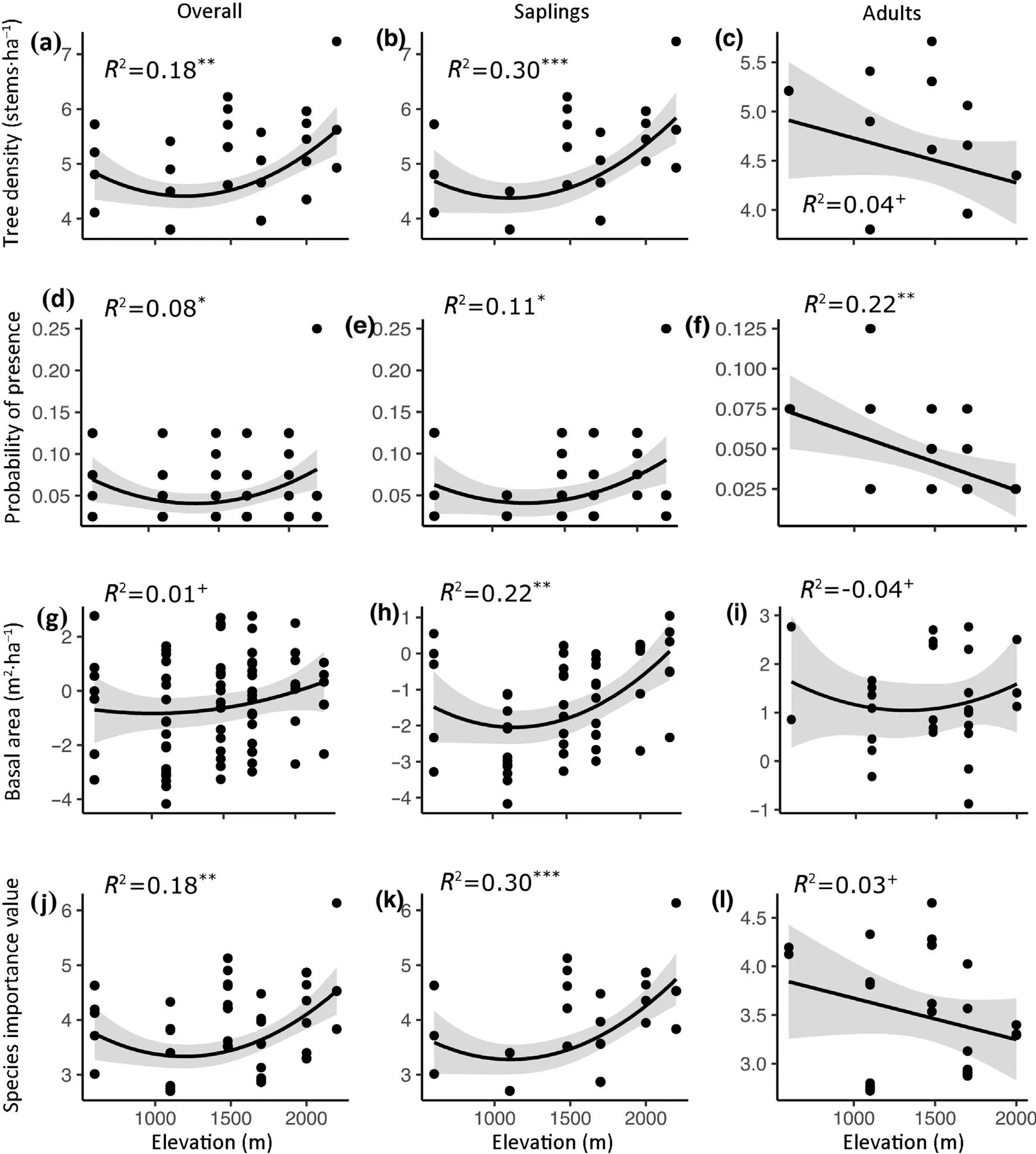
Fig.1.Elevational patterns of indices of tree population in the overall species (a,d,g,j),sapling (b,e,h,k) and adults (c,f,i,l) in the MT.Fanjingshan.The gray shadow indicates the upper and lower 95% confidence interval simulation limits.+P >0.05;*P <0.05;**P <0.01;***P <0.001.
When using a ΔAICc of ≤2,we identified only three alternative models for adult trees (SI Appendix,Table S2) explaining a high proportion of the variance in ecosystem stability(R2=0.64).Climate and leaf traits were responsible for 55.0% and 45%,respectively,of the explained variance of the species importance value (Fig.3).Soil and climate had no significant effects on species importance value,but PC1 was significantly negatively correlated with the species importance value of adult trees(Fig.2).
4.Discussion
AT and AH are the main drivers affecting tree species elevation distribution (Wieczynski et al.,2019).In the current study,the species importance value significantly increased with elevation(Fig.1 j),which was related mainly to the decrease in temperature and increase in humidity (Fig.2,overall species).Previous studies have also shown that species abundance/density increased with decreasing temperature(Steinbauer et al.,2018),but other studies have proven the opposite results(Wason and Dovciak,2017).This inconsistence may be related to local climate factors.For example,when the temperature is very low,the relatively high humidity will cause freezing damage to trees;as such,it is disadvantageous for plants to use water and thus affects tree growth(Humagain et al.,2017).In contrast,when the temperature is suitable,plants can make full use of water,during which trees grow despite increasing humidity (Humagain et al.,2017).Therefore,species elevational migration is related to the interaction effects of temperature and humidity.When the temperature decrease at high altitudes does not exceed the temperature tolerance threshold of plants,increasing humidity is beneficial to plant growth.This opinion is reflected in our study.Although the AT decreases at high altitudes (Fig.S2),the AH increases(Fig.S2),which is beneficial to the survival of species(Fig.2).
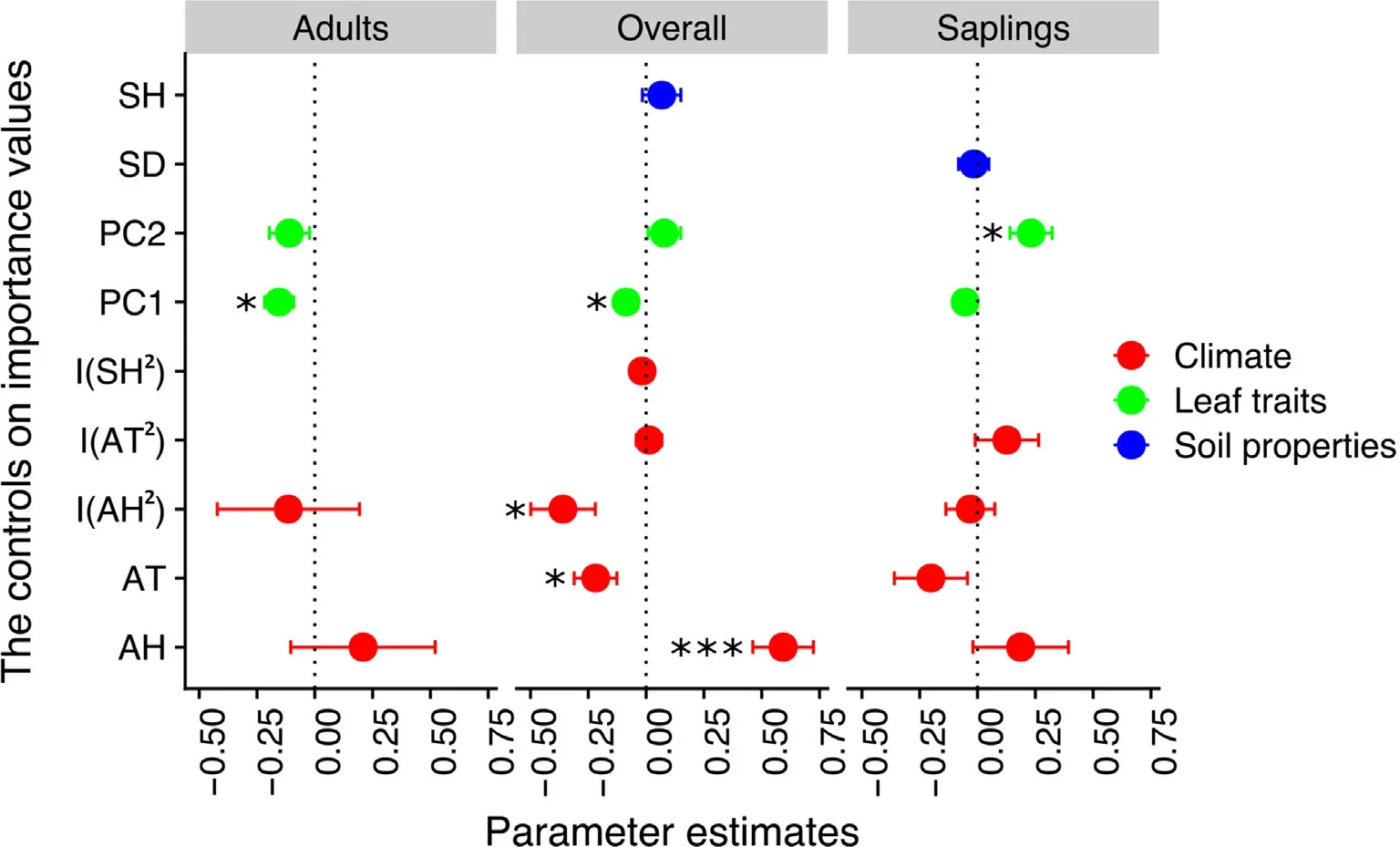
Fig.2.Effects of climate,leaf traits and soil properties on species importance values of overall species,saplings,and adults.Average parameter estimates(standardized regression coefficients) of model predictors associated 95% confidence intervals,and the P value of each predictor are given as:*P<0.05;**P<0.01;***P <0.001.See Supplementary Table 2 for model selection.AT,atmosphere temperature;AH,atmosphere humidity;SH,soil humidity;SD,soil depth;PC1,the first principal component scores of PCA;PC2,the first principal component scores of PCA.

Fig.3.The relative importance of each control expressed as the percentage of explained variance.The R2 of the averaged model for the overall species,saplings and adults were given on top of each bar,respectively.*P <0.05;**P <0.01;***P <0.001.
The elevation distribution of species is affected not only by abiotic factors (e.g.,climate and soil) but also by the leaf traits of trees (Soudzilovskaia et al.,2013).In the present study,woody plant species with nutrient conservation strategies(i.e.,a low SLA and leaf N)were located at high elevations on Mt.Fanjingshan (Fig.S1,Fig.2 overall species),whereas woody plant species at low elevations presented a nutrient foraging strategy (i.e.,a high SLA and leaf N;Fig.S1,Fig.2 overall species) (Read et al.,2014).This trend of the elevation distribution of woody plant species along the ecological axis of the LES has been reported in many previous studies (Read et al.,2014).This is due to environmental pressures such as low temperature,high irradiance,and ultraviolet rays at high elevations in the mountains;thus,species with nutrient conservation strategies are beneficial to for retaining nutrients and resisting disturbances (Read et al.,2014;Reich and Cornelissen,2014;Wright et al.,2004).
In the present work,climate tended to be the most important predictor of sapling and adult distributions(both constituted more than 50%of the variation in species distribution,Fig.3).However,there was a significant difference in elevation distribution between the adults and saplings(Fig.1),in which tree density,basal area and species importance values of the saplings frequently increased significantly with increasing elevation(Fig.1 b,e,h,k).However,the same indices of the adult trees did not change significantly (Fig.1 c,i,l),and even the adult tree frequency decreased significantly with elevation(Fig.1 f).Previous studies have proven that saplings are more sensitive to climate than adult trees are(Wason and Dovciak,2017).However,in our study,climate(i.e.,AT and AH) had a similar influence on both saplings and adults (Fig.3),although the effects of individual climate factors were not significant(Fig.2).Interestingly,we found that saplings and adults employed different mechanisms to adapt to the differences in environments at different elevations,in which the elevation distribution of saplings was related mainly to the leaf carbon content(Fig.S1,Fig.2),while the adults adapted mainly to these environments via a nutrient conservation strategy(Fig.S1,Fig.2).We know that the increase in sapling abundance at high elevations is due mainly to the migration of low-elevation species and/or the regeneration of species at originally high elevations.Previous studies have shown that species abundance significantly increases with increasing elevation (Steinbauer et al.,2018),and appropriately low temperatures can improve species regeneration.In the current study,the AT decreased with increasing elevation(Fig.S2)and played a critical role in the distribution pattern of saplings(Fig.2).However,it was difficult to determine whether the migration of low-elevation species or the regeneration of species at original elevations led to an increase in sapling abundance at high elevations;saplings encounter low temperatures and low light levels in the overstorey,which limits the photosynthesis of sapling trees and may have caused them to die because of a lack of carbohydrates (Rowland et al.,2015).Therefore,to survive at high elevations,saplings need a large amount of carbon to maintain their growth and metabolism.Consequently,the elevation distribution of saplings was positively correlated with the principal component axis of the leaf carbon content (Fig.S1,Fig.2) but was not significantly related to the LES(Fig.S1,Fig.2),which indicated that carbohydrates,not nutrients,were the main limiting factor for the growth of saplings at high elevations in the present study.Therefore,where does this carbon mainly originate from?This is a question that needs to be answered.According to previous studies,we speculate that this portion of carbon is mainly transported from large trees near saplings through underground mycorrhizal network systems (Gorzelak et al.,2020;Pec et al.,2020;Simard et al.,1997),and/or saplings reduce their energy supply and their consumption of absorbed nutrients(which is not significantly related the LES)to supply more C to saplings(Pec et al.,2020).However,these inferences need to be verified in future studies.
Unlike that of saplings,the elevation distribution of adults was related to the LES,and their leaves mainly adopted a nutrient conservation strategy (Fig.S1,Fig.2).Woody plant species are anchoring organisms that regenerate and migrate mainly through seed dispersal.However,with environmental changes,adults have no chance to ‘move from their position’;thus,they adopt nutrient conservation strategies only to maintain their nutrients and resist external disturbances(Wright et al.,2004).At the same time,the nutrient conservation strategy of adults is also beneficial for transferring additional nutrients,carbohydrates,and biological information to saplings for their regeneration and migration.In addition,we found that leaf traits and climate factors have a significant impact on the overall species distribution,while the distribution of saplings and adults only were significant correlated with leaf traits(Fig.2).These results indicate that the leaf traits of species have a certain impact on the distribution of species in the present study.This has been reported in the previous studies (Soudzilovskaia et al.,2013;Vesk et al.,2020).Because the distribution of woody plants is affected by seed dispersal,colonization,and survival ability (May et al.,2013).Among these three factors,leaf traits are closely related to their colonization and survival ability (Reich and Cornelissen,2014),and therefore also affect the distribution trend of species(Vesk et al.,2020).In other words,only species with the ability to plasticize specific traits can colonize and survive in a specific environment,and then determines the distribution pattern of species.Therefore,the elevational adaptation strategies of woody plant species can be explained in depth from the perspective of leaf functional traits.
Recent studies have shown that species distributions shift in varying directions,with some shifting upward,some shifting downward,and some not changing(O'Sullivan et al.,2020).Therefore,it is important to determine the elevation distribution of specific woody plants for regional species protection and management.However,because of the steepness of Mt.Fanjingshan,it was difficult to design a large-scale plot survey.Therefore,this study adopted the point-centred-quarter community survey method to record species elevation distributions.Based on the existing data,it is difficult to disentangle the direction and magnitude of specific species along the elevation gradient on Mt.Fanjingshan.Consequently,it is necessary to overcome these difficulties and generate large-scale plots to answer these questions.Nevertheless,the point-centred-quarter survey method is a classic plant community survey method and has been widely used in the past(Wason and Dovciak,2017).Therefore,the results of this study show that mountains are the main refuge for species in the context of climate warming,and the elevational adaptation mechanism of different species of trees of different sizes of can be strongly explained based on plant functional traits (such as leaves),which play a critical role in species protection and management in nature reserves(i.e.,Mt.Fanjingshan).
5.Conclusions
The elevation distribution of woody plant species on Mt.Fanjingshan was driven mainly by AT,AH,and leaf traits,but the trends of saplings and adults were different;sapling abundance increased significantly with increasing elevation,while adults showed no significant change in abundance.These results suggest that the adaptation mechanisms to environment at different elevations differ between saplings and adults,in which the elevation distribution of saplings was related mainly to the leaf carbon content,while the elevation distribution of adults was related to the LES.Therefore,when analysing the elevation distribution pattern of woody plant species,researchers cannot ignore the role of plant functional traits.Future studies should disentangle the patterns and mechanisms of climate,soil,and plant functional traits on the elevational migration of specific species and explain the relative importance of lowelevation species migration and original elevational species regeneration on the elevation distribution of saplings,which is highly important for species protection and management in high-mountain regions.Consequently,more data from large plots are needed to determine the driving factors and mechanisms of the elevation distributions and migration patterns of specific species.
Ethics approval and consent to participate
The subject has no ethic risk.
Consent for publication
Not applicable.
Availability of data and material
Data available on request from the authors.
Competing interests
The authors declare that they have no conflict of interest.
Funding
This research was financially supported by PhD Research Start-up Foundation of Tongren University (trxyDH1807,trxyDH1826,trxyDH1806),the Natural Science Foundation of Guizhou Provincial Department of Education ([2019]075),the Science and Technology Project of Guizhou Province (ZK-[2021]221),the National Nature Science Foundation of China(31900271),and the Key Laboratory Project of Guizhou Province([2020]2003).
Authors' contributions
ZHW and ML convinced conceptualization.ZHW analyzed data.ZHW led the writing of the manuscript.ML and LLY led the review and editing of the manuscript.All authors made contributions to data collection via fieldwork or lab work.All authors read and approved the final manuscript.
Acknowledgements
We sincerely thank Zongkun Shi,Zhongfei Pan,Chaohui Ran,Qian Luo,Fangyi Wei,Jiayan Jiang,Huiqing Yang,Jinyan Zhou,Gen Li and Ling Tian for help in the field and laboratory.
Appendix A.Supplementary data
Supplementary data to this article can be found online at https://do i.org/10.1016/j.fecs.2022.100012.

- Forest Ecosystems的其它文章
- A better carbon-water flux simulation in multiple vegetation types by data assimilation
- Sensitivity analysis of Biome-BGCMuSo for gross and net primary productivity of typical forests in China
- Environmental and biological factors affecting the abundance of Prosopis flexuosa saplings in the central-west Monte of Argentina
- Modifying regeneration strategies classification to enhance the understanding of dominant species growth in fire-prone forest in Southwest China
- Climate-driven variations in productivity reveal adaptive strategies in Iberian cork oak agroforestry systems
- Conifer establishment after the eruption of the Paricutin volcano in central Mexico