
高盐盐湖可分离嗜盐耐盐菌的种群多样性及四氢嘧啶产量评价
2022-03-07 00:55张田田李永臻沈国平王嵘朱德锐邢江娃
生物技术通报 2022年1期
张田田 李永臻 沈国平 王嵘 朱德锐 邢江娃
(青海大学医学院基础医学研究中心,西宁 810016)
柴达木盆地位于青藏高原东北部(90°00′-98°20′E,35°55′-39°10′ N),海拔 2 675-3 350 m,隶属高原大陆性干旱气候[1-3],盆地环境相对封闭,复杂的地质条件和独特的环境因素形成了地球上分布面积最大(2.0×106km2)、数量最多以及盐碱湖集中的特色高原盐湖群[4-5]。该区域内湖泊类型多样化,其总盐度(或离子组成)、水质营养(总碳、氮或磷)以及pH等环境因子存在显著差异。根据矿化度含量,盐湖可分为淡水湖(矿化度≤1.0 g/L)、微咸水湖(矿化度1.0-35.0 g/L)、咸水湖(矿化度35.0-50.0 g/L)和高盐盐湖(矿化度>50.0 g/L)4种类型[6];根据化学成分不同,又可分为碳酸盐型、硫酸盐型和氯化物型3种类型,其中碳酸盐湖又分为高碳酸盐型(>29.0%)、中碳酸盐型(8%-29.0%)和弱碳酸盐型(0.1%-8.0%)3个亚型,而硫酸盐型盐湖又分为硫酸钠亚型和硫酸镁亚型[7]。
盐湖微生物是湖泊生态系统的重要组成部分,其微生物多样性、群落结构组成和环境互作受到盐湖各类水化学因素的制约,如水化学类型、地理位置间距、总盐度、水质营养(总碳、氮或磷)、温度和pH等[8]。根据微生物对盐浓度的不同需求,可将其分为7类[9]:非嗜盐微生物(non-halophile,0-1.0 mol/L)、弱嗜盐微生物(slight halophile,0.2-2.0 mol/L)、中度嗜盐微生物(moderate halophile,0.4-3.5 mol/L)、边界极端嗜盐微生物(borderline extreme halophile,1.4-4.0 mol/L)、极端嗜盐微生物(extreme halophile,2.0-5.2 mol/L)、耐盐微生物(halotolerant,0->1.0 mol/L)和万能耐盐微生物(haloversatile,0->3.0 mol/L)。在应对高盐环境胁迫时,嗜盐微生物胞内可积聚多种代谢产物,如嗜盐菌素、抗生素、胞外多糖、相容溶质四氢嘧啶(ectoine)和甜菜碱等,这些特殊代谢物可广泛应用于农业生产[10]、食品加工[11]、环境治理[12-15]以及生物医药等方面[16-19]。国内学者曾以达布逊盐湖、青海湖、托素湖、尕海、茶卡盐湖、柯柯盐湖、察尔汗盐湖和小柴旦盐湖等为研究对象,通过高通量测序等手段发现大量的芽孢杆菌(Bacillus)、盐单胞菌(Halomonas)、大洋芽胞杆菌属(Oceanobacillus)、假单胞菌属(Pseudomonas)、乳球菌属(Lactococcus)和葡萄球菌属(Staphylococcus)等菌株资源广泛存在[2-3,20-22]。然而,目前自然环境中仅有1%左右的微生物获得纯培养,甚至更低[23]。本研究以茶卡盐湖、柯柯盐湖和小柴旦盐湖等三大硫酸镁亚型高盐盐湖为研究对象,采用RM培养基筛选分离可培养嗜盐和耐盐菌,分析其群落结构多样性及其与环境因子之间的关系,了解优势菌属的嗜盐特性和系统发育,并采用HPLC筛选四氢嘧啶高产野生菌株,为盐湖微生物资源的开发与利用提供重要的理论参考依据。
1 材料与方法
1.1 材料
1.1.1 样品采集 2018年7月采集茶卡盐湖(99°4′41.67′E,36°45′0.05′N)、柯柯盐湖(98°15′3.4′E,36°58′19.97′N)和小柴旦盐湖(95°32′58.38″E,37°28′0.65″N)的水泥混合样本共计12个,浑浊状态,水样采集深度10-25 cm,采样温度20-27.7℃,各样点间距大于100 m。样本储存于4℃车载冰箱,运回实验室备用。
1.1.2 培养基制备 菌种分离和富集培养采用RM固体与液体培养基[1]:NaCl 50 g或 100 g,MgSO4·7H2O 9.7 g,KCl 2.0 g,柠檬酸钠3.0 g,无水氯化钙0.2 g,酵母抽提物2.0 g,蛋白胨10.0 g,加入蒸馏水定容至1 L,调节pH 7.5。固体培养基加琼脂16.0 g,1×105Pa灭菌1 h,保存备用。
1.2 方法
1.2.1 菌株分离与耐盐实验 取适量水样100 μL直接涂布固体培养基平板,置入37℃恒温培养箱,培养18-24 h后取出。依据菌落的形态进行单一菌落挑选,重复划线纯化培养3-4次,最后-20℃保藏菌种(菌液含15%甘油)。耐盐度试验以1%接种量(V/V,1∶100)接种于不同NaCl浓度梯度的RM液体培养基中(0-3.0 mol/L,梯度0.5),3组平行,37℃,150 r/min振荡培养12、18和24 h。采用722分光光度计测定培养液光密度值(OD600),取其平均值,根据生长曲线计算最适盐度。
1.2.2 细菌DNA的提取、PCR扩增与测序 基因组DNA提取按照细菌基因组DNA提取试剂盒说明进行。细菌16S rRNA基因PCR扩增和测序采用通用引物 27F(5′-AGTTTGATCMTGGCTCAG-3′) 和 1492R(5′-GGTTACCTTGTTACGACTT-3′)进行。PCR 反应体系(25 μL):10×Buffer 2.5 μL,2.5 mmol/L dNTPs 1 μL,正、反向引物(浓度为 10 μmol/L)各 0.5 μL,DreamTaq-TM DNA聚合酶 0.2 μL,DNA模板(20-50 ng/μL)0.5 μL,ddH2O 补充至 25 μL。PCR 反应条件:94℃ 4 min,94℃ 45 s,55℃ 45 s,72℃ 60 s,30 个循环,最后72℃修复延伸10 min。PCR产物经凝胶电泳观察和纯化回收后用PCR引物直接测序,获得待测菌株的16S rRNA序列。
1.2.3 菌种鉴定与优势菌株系统发育分析 将测序得到的分离菌株16S rRNA基因序列在EzBioCloud(http://www.ezbiocloud.net/identify)和 NCBI GenBank(http:// blast.ncbi.nlm.nih.gov/Blast. cgi)数据库进行比对分析,确定分离菌株的分类学地位,两个数据库比对结果不同时,以相似度较高的结果为准(相似度≥98%)。通过NCBI数据库获取与菌株相似性较高的近缘物种参考基因序列,利用MEGA 7.0软件进行相似性比对,采用邻接法(neighbor joining,NJ)构建系统发育进化树(bootstrap为1 000)[24-25],以此分析优势分离菌株的系统发育地位。
1.2.4 四氢嘧啶的提取和细胞干重测定 将培养12 h的菌液扩大培养到100 mL锥形瓶中,放入摇床过夜培养24 h后测OD600,然后抽取10 mL于离心管中,12 000 r/min离心5 min,弃上清;加入纯水10 mL,用超声细胞破碎仪破碎7 min,12 000 r/min离心5 min,取其上清经0.22 μm水系微孔过滤器过滤,即四氢嘧啶提取液[26]。将剩余90 mL发酵液4℃12 000 r/min离心5 min,弃上清,再加10 mL的生理盐水快速洗涤,9 000 r/min 离心5 min,弃上清,将沉淀放于60℃烘干箱至恒重,冷却后称量,计算细胞干重(CDW g/L)。
1.2.5 高效液相色谱(HPLC)检测四氢嘧啶含量 配制1.0 mg/mL的四氢嘧啶标准品母液,HPLC检测四氢嘧啶浓度,并建立标准曲线(峰面积与浓度)。HPLC检测条件:流动相为水∶乙腈(V/V,20∶80),检测波长210 nm,流速为1.0 mL/min,柱压 3.486-4.761 MPa,柱温 30℃,上样量 15 μL[27].
1.2.6 环境因子相关性分析 基于三大盐湖的水质离子特征数据和分离菌株菌种鉴定结果,采用多元直接梯度回归法分析菌群与环境因子之间的相关性。采用R语言Vegan包进行典范对应分析(canonical correlation analyses,CCA)和作图,通过bioenv函数判断环境因子与样本群落分布差异的最大Pearson相关系数,通过permutest分析判断CCA分析的显著性。
1.2.7 NCBI数据提交 本研究所得序列均提交NC BI(http://www.ncbi.nlm.nih.gov/)数据库,细菌登录 号 MT982223-MT982264;MT982266- MT982321;MT982324- MT982326;MT982332-MT982343。
2 结果
2.1 分离培养细菌的分子鉴定与统计
本研究筛选获得茶卡盐湖、柯柯盐湖和小柴旦盐湖的可培养细菌共计113株(表1)。其中,RM中盐培养基(5% NaCl)共筛出菌株70株,高盐培养基(10% NaCl)共筛选出菌株43株。统计分析表明:所有分离菌株来自厚壁菌门(Firmicutes)和变形菌门(Proteobacteria)2个门(表2)。中盐培养基筛选出的优势菌属为Firmicute门Bacillus属(32株);高盐培养基筛选出的菌株主要分布在Firmicutes门的Staphylococcus属(15株)和Bacillus属(10株),以及Proteobacteria门的Halomonas属(15株)。其中,茶卡盐湖共分离获得47株可培养细菌,优势菌株为Bacillus属(20株,约占42.6%)和Halomonas属(10株,约占21.3%);柯柯盐湖共分离获得40株可培养细菌,优势菌株为Staphylococcus属(17株,约占42.5%)和Bacillus属(16株,约占40%);小柴旦盐湖共分离获得26株可培养细菌,优势菌株为Halomonas属(13株,约占50%)。

表1 分离菌株数据统计Table 1 Statistics of isolated strains
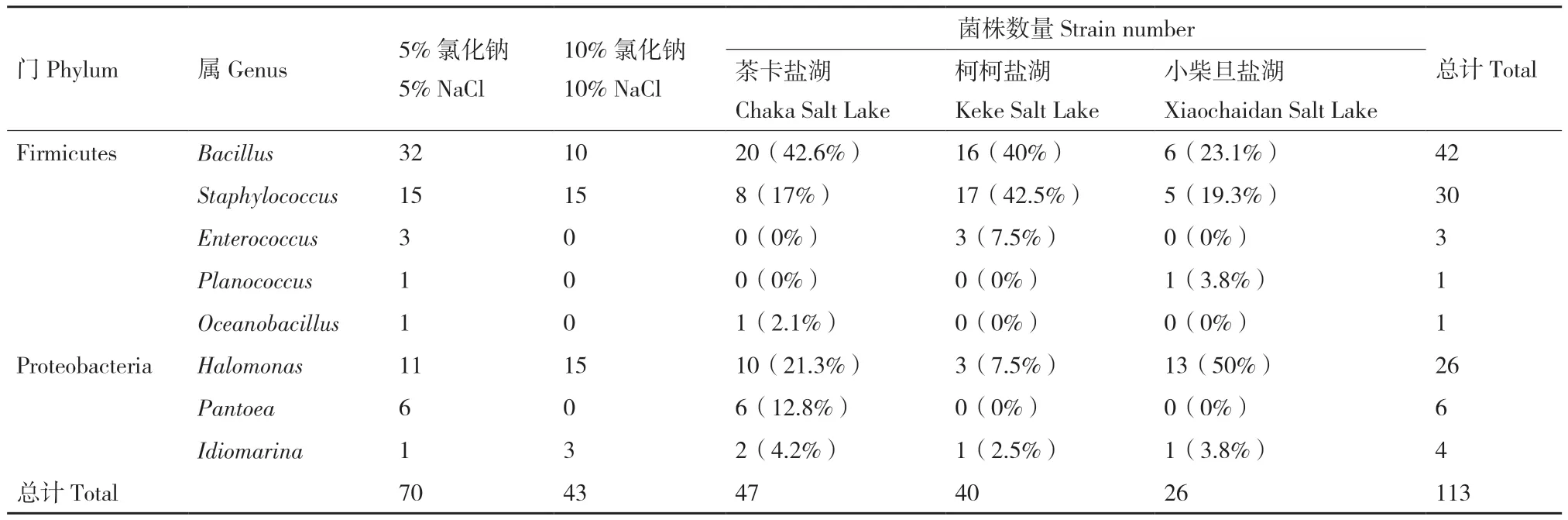
表2 柴达木盆地典型盐湖分离细菌的分子鉴定与丰度统计Table 2 Molecular identification and abundance statistics of the isolated bacteria from typical salt lakes in Qaidam Basin
2.2 环境因子CCA分析
茶卡盐湖、柯柯盐湖和小柴旦盐湖为柴达木盆地典型的硫酸镁亚型高盐盐湖,总盐度均值分别为 292.64 g/L、256.95 g/L和 110.82 g/L,以 Na+、Mg2+、Cl-、SO42-离子为主[2,22,28]。CCA 分析结果显示(图1):由于3个盐湖的理化性质相似,各环境因子对样本的影响程度基本相当,没有显著的环境影响因子。各环境因子之间总盐度(TS)、有机碳(TOC)、有机氮(TN)与 Na+、Cl-、K+、Mg2+、CO32-离子成正相关,与pH、Ca2+和SO42-离子成负相关。优势菌属Bacillus的丰度主要与Mg2+和K+成正相关,Staphylococcus主要与TN和CO32-成正相关,Halomonas主要与pH和SO42-离子成正相关。
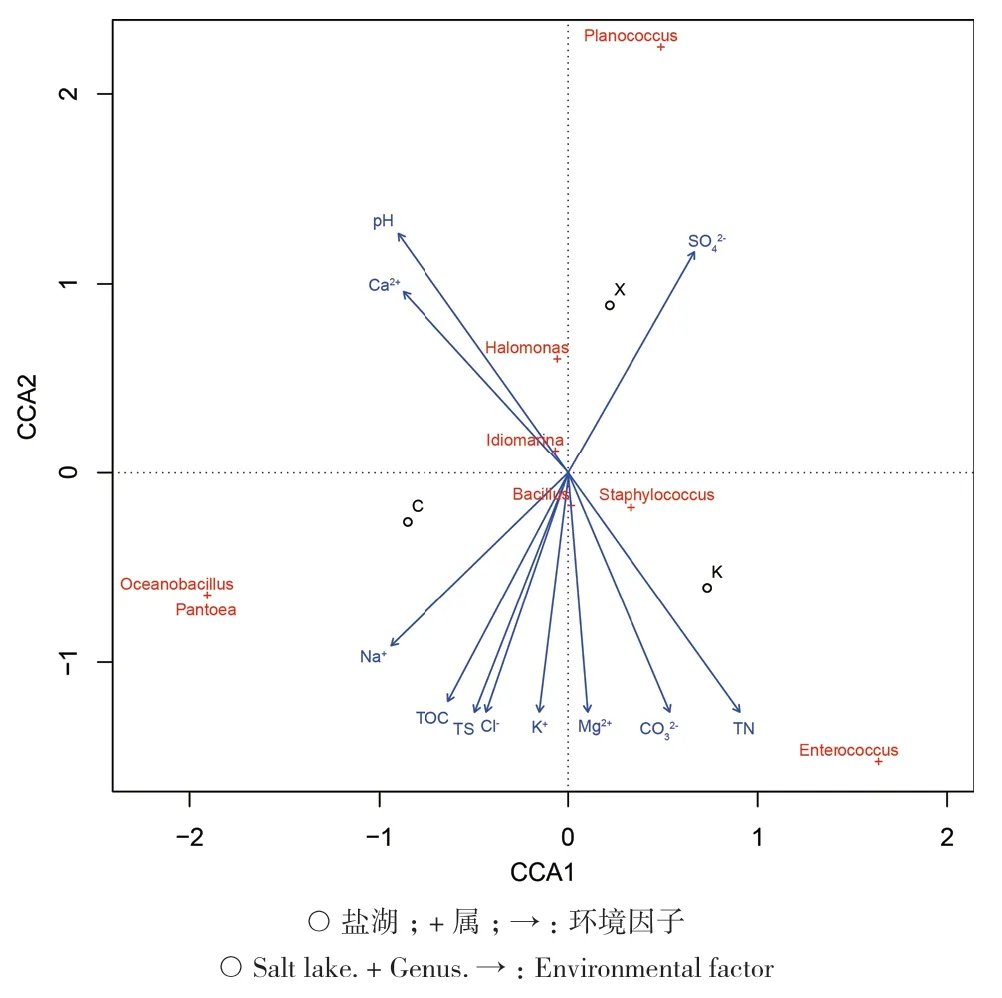
图1 三大盐湖分离细菌环境因子CCA分析Fig. 1 CCA analysis between environmental factors and bacteria isolated from the three salt lakes
2.3 优势菌株菌落形态、嗜盐特性与分类统计
分离的Bacillus属菌株(42株)多为圆形或椭圆形,形态大小稍有差异,菌落颜色多呈白色或乳白色,部分呈乳黄色或土黄色,极少数呈粉色(如菌株C43、C18和X20)和浅褐色(如菌株C37和C10)。在生长盐度范围上,大多数菌株在0-3.0 mol/L范围都可以生长,仅个别菌株在0-2.0 mol/L或更低的盐度下才能生长(表3)。参考Kushner的分类标准[9],21株Bacillus属菌株为中度嗜盐菌,约占50%;16株为耐盐菌,约占38.1%;5株为弱嗜盐菌,约占11.9%。Halomonas属菌株(26株)多为圆形或椭圆形,菌落颜色呈白色或乳白色,少量呈现褐色(如菌株C9)和黄色(如C39、C40和C44)。大多数Halomonas菌株的生长盐度范围为0-3.0 mol/L,生长范围宽泛且最适盐度较高,易于培养。其中,24株隶属中度嗜盐菌,约占92.3%,2株隶属耐盐菌范畴,约占7.7%。
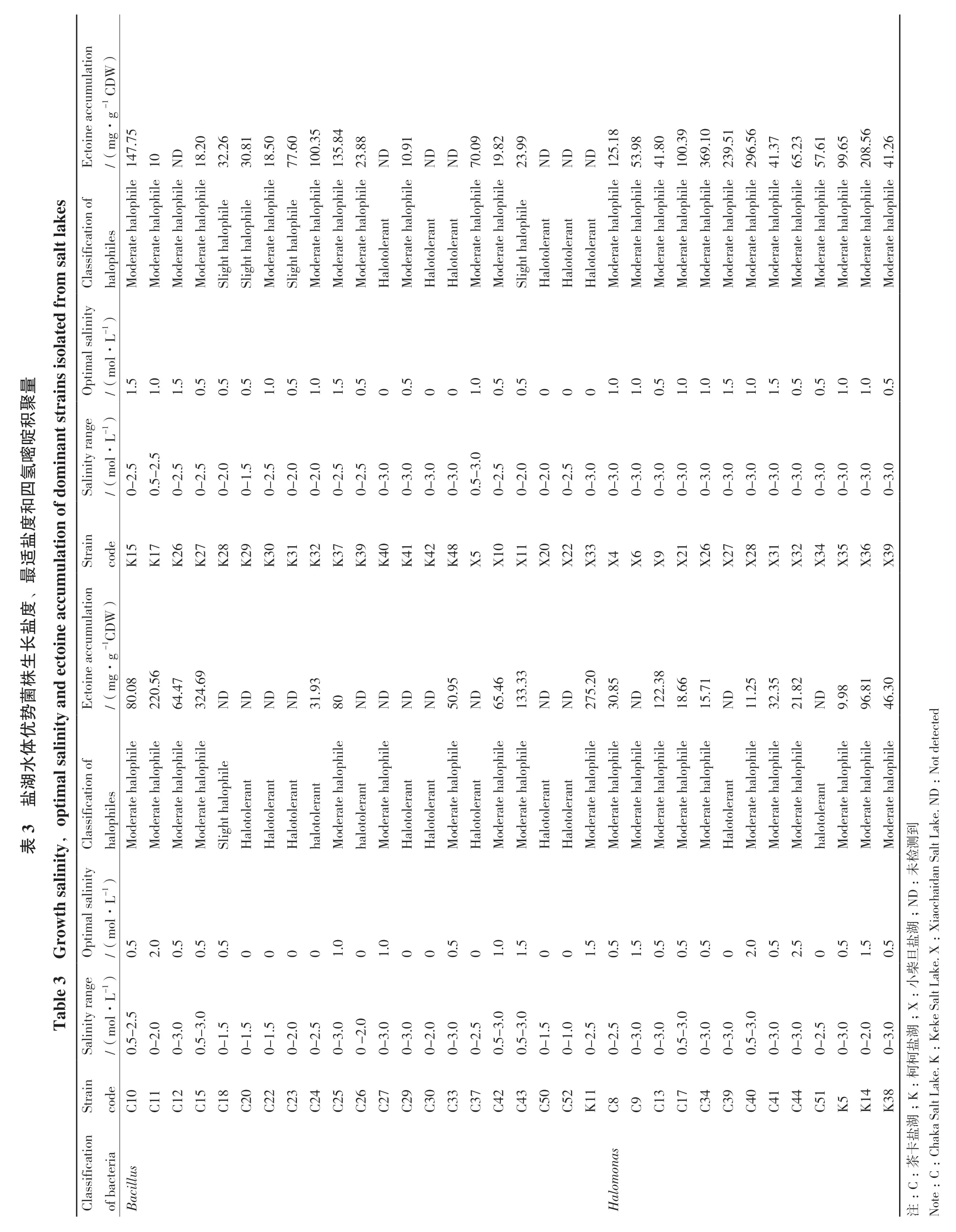
ulation )Ectoine accum mg·g -1 C DW/(147.75 10ND 18.20 32.26 30.81 18.50 77.60 100.35 135.84 23.88 ND 10.91 ND ND 70.09 19.82 23.99 ND ND ND 125.18 53.98 41.80 100.39 369.10 239.51 296.56 41.37 65.23 57.615 99.6 208.566 41.2 es ted from salt lak Classification of hile hile hile halophiles Moderate halophile Moderate halophile Moderate halophile Moderate halophile Slight halop Slight halop Moderate halophile Slight halop Moderate halophile Moderate halophile Moderate halophile Halotolerant alophile Moderate h Halotolerant Halotolerant alophile Moderate h alophile Moderate h Slight halophile Halotolerant Halotolerant Halotolerant Moderate halophile Moderate halophile Moderate halophile Moderate halophile Moderate halophile Moderate halophile Moderate halophile alophile Moderate h alophile Moderate h alophile Moderate h alophile Moderate h alophile Moderate h alophile Moderate h积t strains iso la聚L-1)mol·量Optimal salinity/(1.51.01.50.50.50.51.00.51.01.50.500.5001.00.50.50001.01.00.51.01.01.51.01.50.50.51.01.00.5啶an ge Salinity ranL-1)嘧in mol·/(0-0.5-2.5氢om 2.52.5 0-0-0-0-0-0-0-0-0-0-0-0-0-0.5-2.5 2.52.01.52.52.02.02.52.53.03.03.03.03.0 2.02.02.53.03.03.03.03.03.03.03.03.03.03.03.03.03.0四0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-0-和度 适ulatio n of d盐最Strain code K15 K17 K26 K27 K28 K29 K30 K31 K32 K37 K39 K40 K41 K42 K48 X5X10 X11 X20 X22 X33 X4X6X9X21 X26 X27 X28 X31 X32 X34 X35 X36 X39度、盐mg·g -1CD e accum ulation e accum W)长in株生ecto 8菌/(80226432ND ity and Ectoin.00.56.474.69.8.66.71势ND ND ND.93 3180ND ND ND ND 50.95 ND 65.46 133.33 ND ND5.205 2730ND2.38 121815ND.25 1132.35 21.82 ND 9.98 96.81 46.30优体水湖al salin hile alophile alophile alophile alophile alophile alophile alophile alophile Not detected 3 盐ptim D:salinity,o Classification of halophiles Moderate halophile Moderate halophile检Salt L Moderate halophile Moderate halophile表Slight halop Halotolerant Halotolerant Halotolerant halotolerant Moderate halophile halotolerant Moderate halophile Halotolerant Halotolerant Moderate h Halotolerant Moderate h Moderate h Halotolerant Halotolerant Moderate h Moderate halophile Moderate halophile Moderate halophile Moderate halophile Moderate halophile Halotolerant Moderate halophile Moderate halophile Moderate h halotolerant Moderate h Moderate h Moderate h测ake. N到ity 未1)th D:L-Grow timal salin;N湖iaochaidan Op /(0.5 mol·2.00.50.50.500001.001.0000.501.01.5001.50.51.50.50.50.502.00.52.500.51.50.5盐:X旦Table 3 Salinity range L-1)柴ake. X小mol·/(0.5-2.02.5:3.0;X 3.03.0 e Salt L 0.5-1.5 0-0-0-1.5 0-1.5 0-2.0 0-2.5 0-3.02.0 0-0 -3.0 0-0-0-0-0-0.5-2.03.0 3.02.53.0 0.5-1.5 0-0-0-2.5 1.02.5 0-3.0 0-3.03.0 0-0.5-3.0 0-3.03.0 0-0.5-3.0 0-0-0-0-0-0-2.5 3.03.02.03.0湖ek盐:K柯Strain 柯code C11 C158023456790372302:ake. K C12 C2 C1C2C2C2C2C2C2C2C3C3C3C4C4C5C5K11 C8C9C137490141 BacillusC10 C1C3C3C4C4C4C5K5K148 K3;K湖Classification 盐of bacteria lomonas 卡茶:C haka Salt L:Ha :C:C注Note
2.4 构建优势菌株系统发育树
系统发育分析结果显示柴达木盆地极端盐湖Bacillis属主要有5个进化分支,属于6个不同的种(图2-A)。分离菌株 C10、C11、C15、C24、C25、C27、C29、C30、C33、C42、C43、C50、K11、K29、K30、X5、X11、X20和X22 与菌株B. zhangzhouensis(MG547767.1)聚类在同一进化分支,基因相似性为99.87%-99.93%,表明进化同源;菌株C37、K17、K26、K32、K37、K39和 K42与 菌 株B. pumilus(CP027116.1)聚类在同一进化分支,基因相似性为99.93%-100%,表明进化同源;菌株C12、C18、C20、C23、C52、K27、K28、K31、K40、K41、K48、X10和X33与菌株B. atrophaeus(NR024689.1)聚类在同一进化分支,基因相似性为99.93%-100%,表明进化同源;C22、C26和K15分别与菌株B. paralicheniformis、B. flexuss和B. subtili基因相似性为100%,表明进化同源。Halomonas属主要有6个进化分支(图2-B),属于7个不同的种分类单元,此外3株菌株仅能鉴定到属,进一步分类学地位尚需鉴定。菌株C9、C13、C34和X4与菌株H. zhaodongensis(NR 125612.1)基因相似性为98.79%-99.86%,表明进化同源;菌株K5和 K14与 菌 株 Halomonas sp. TC19(EU308353.1)基因相似性为100%,表明进化同源;菌株C8、K38、X6、X9、X21、X26、X31、X32、X34、X35与 X39与 H. alkaliphila(CP024811.1) 基 因 相 似度为99.73%-100%,表明进化同源;C39和C44分别与菌株H. arcis和H. salicampi基因相似性为99.22%和98.72%,表明进化同源;菌株 C17与H.gomseomensis 基因相似性为98.14%,但两者并未聚类在一起,可能为潜在的新种;C40与H. lutescens的基因相似性为98.14%,但两者并未聚类在一起,可能为潜在的新种;C51与H. zhaodongensis的基因相似性为98.16%,但两者并未聚类在一起,可能为潜在的新种。

图2 优势菌株系统发育进化树(A:芽孢杆菌属,B:盐单胞菌属)Fig. 2 Phylogenetic tree of dominant strains(A :Bacillus. B :Halomonas)
2.5 四氢嘧啶含量测定
硫酸镁亚型三大高盐盐湖优势分离菌株之间的四氢嘧啶含量存在较大差异(如K5和X26)(表3)。其中,检测到的Bacillus属单位细胞干重四氢嘧啶积聚量低于50 mg/g的菌株共计10株;积聚量在50-100 mg/g的菌株共计7株;积聚量高于100 mg/g的菌株共计7株。Halomonas属四氢嘧啶积聚量低于50 mg/g的菌株共计11株;积聚量在50-100 mg/g的菌株共计5株;积聚量高于100 mg/g的菌株共计7株。两个属中共有21株菌株未检测到四氢嘧啶的积聚。Halomonas属 的 X26、X28、X27和 X36与 Bacillus属的C15、K11和C11等7株菌株的四氢嘧啶积聚量显著高于其他菌株(大于200 mg/g),具有高效积聚生产四氢嘧啶的潜力。
3 讨论
高盐盐湖环境中广泛存在不同的嗜盐微生物[28]。诸多研究表明:高盐盐湖的细菌优势属群主要有假单胞菌属(Pseudomonas)、芽孢杆菌属(Bacillus)、希瓦式菌属(Shewanella)、盐单胞菌属(Halomonas)、不动杆菌属(Acinetobacter)、梭状芽孢杆菌(Clostridium)、泛菌属(Pantoea)、分枝杆菌属(Mycobacterium)、交替单胞菌属(Alteromonas)、嗜盐杆菌属(Halobacterium)、海杆 菌 属(Marinobacter)、 肠 球 菌(Enterococcus)、盐红菌属(Halorubrum)、嗜盐碱单胞菌属(Natronomonas)、极嗜盐菌属(Haloferax)、红球菌属(Rhodococcus)、硝化细菌(Nitrobacter)、深海弯曲菌属(Thalassolituus)、栖苏打菌属(Nitrincola)、盐杆菌属(Salinibacter)、海茎状菌属(Maricaulis)、罗丹杆菌属(Rhodanobacter)、玫瑰杆菌(Roseobacter)和脱硫杆菌属(Desulfobacterium)等,并因各盐湖水化学特征和环境因子的不同而存在明显差异[29-35]。柴达木盆地盐湖众多,细菌类型丰富,优势物种组成多样化。沈硕[1]采用16S rRNA基因测序分析察尔汗盐湖(310.01-332.82 g/L)的细菌多样性,结果显示优势菌属以Bacillus(33%)、Oceanobacillus(25%)和 Halomonas(17%)为主;赵婉雨等[20]采用高通量测序与传统培养技术结合的方法分析达布逊湖(371 g/L)微生物多样性,表明优势菌属以Pseudomonas和Bacillus为主;课题组前期基于16S rRNA基因高通量测序分析发现硫酸镁型高盐盐湖茶卡盐湖[22]、柯柯盐湖[2]和小柴旦盐湖[28]的优势细菌属群主要是Bacillus(8.24%-58.3%),次为Pseudomonas(1.4%-10.5%)、Oceanobacillus(1.5%-10.3%)、Lactococcus(1.1%-10.5%) 和 Halomonas(0-7.4%)等。本研究采用RM中盐和高盐培养基分离出茶卡盐湖、柯柯盐湖和小柴旦盐湖的可培养嗜盐耐盐菌共计113株,优势菌属为Bacillus属(42株)、Staphylococcus属(31株)和Halomonas属(26株),其中Bacillus和Halomonas的丰度与盐湖类型呈明显的正相关。分离出的优势细菌菌属与前期的高通量测序分析结果大体一致,表明选用的RM中盐和高盐培养基比较适合这两类优势菌群的筛选和分离培养,且Bacillus和Halomonas很可能是硫酸镁亚型高盐盐湖的典型优势菌群。前期高通量测序得到的优势菌属Pseudomonas、Oceanobacillus和Lactococcus在本研究中尚未分离到,需要后期优化相应的菌株分离和筛选培养基的类型。同时,本研究中分离到的嗜盐菌主要为中度嗜盐菌,并未分离到极端嗜盐菌,可能与分离培养基中NaCl最高盐度设置偏低有关。
优势分离菌属Bacillus属分类学定位于芽孢杆菌纲芽孢杆菌科,是一类产芽孢的好氧、或兼性厌氧性革兰氏阳性细菌[36],广泛存在于盐湖、沙漠、海洋或火山谷等特殊环境之中[37]。截止2020年,LPSN数据库(http://www.bacterio.net/)已收录的Bacillus属共计95个种,代表性菌株如B. atrophaeus、B. mycoides和B. cereus等。本研究共计获得优势纯培养Bacillus属菌株42株,主要有萎缩芽孢杆菌(B. atrophaeus)、短小芽孢杆菌(B. pumilus)、张州芽孢杆菌(B. zhangzhouensis)、枯草芽孢杆菌(B. subtilis)、弯曲芽孢杆菌(B. flexus)和地衣芽孢杆菌(B. paralicheniformis)等,细菌分类以中度嗜盐菌(约占50%)和耐盐菌(约占38.1%)为主。优势分离菌属Halomonas属分类学定位于γ-变形菌纲盐单胞菌科,能在含有0-5.1 mol/L NaCl环境中生长,是一类需氧、杆状革兰氏阴性细菌,广泛分布于盐湖、盐场或海洋等极端盐环境[38]。截止2020年,LPSN数据库已收录Halomonas属菌株共计108个种。Halomonas属是嗜盐微生物重要的代表菌群,也是盐环境优势属群之一[39],代表菌株如H. elongata、H. alkaliphile和H. marina等。本研究中,共计获得纯培养Halomonas属菌株26株(其中3株菌种分类未确定),主要有嗜碱性盐单胞菌(H. alkaliphile)、尖顶盐单胞菌(H. arcis)以及肇东盐单胞菌(H. zhaodongensis)等,细菌分类以中度嗜盐菌(约占92.3%)为主。沈硕[1]通过可培养方法发现察尔汗盐湖Bacillus种属主要为:甲基营养芽孢杆菌(B. methylotrophicus)、枯草芽孢杆菌(B.subtilis)、短小芽孢杆菌(B. pumilus)、暹罗芽孢杆菌(B. siamensis)和萎缩芽孢杆菌(B. atrophaeus)等,Halomonas种属主要为:尖顶盐单胞菌(H.arcis)、H. zhaodongensis和泰坦尼克号盐单胞菌(H.titanicae)等。李坤珺等[40]通过可培养方法发现运城盐湖Bacillus种属主要为:甲基营养芽孢杆菌(B.methylotrophicus)、沙地芽孢杆菌(B. aquimaris)、花津滩芽孢杆菌(B. hwajinpoensis)和盐芽胞杆菌(B.salarius)等,Halomonas种属主要为:尖顶盐单胞菌(H. arcis)、硫化物单胞菌(H. sulfidaeris)、艾丁湖盐单胞菌(H. aidingensis)、脱氮芽孢杆菌(H.denitrificans)和韩国芽孢杆菌(H. koreensis)等。与以上盐湖菌株多样性相比,本研究分离得到的优势菌属同为Bacillus和Halomonas属,但具体的菌种略有不同,可能与各盐湖环境与理化性质的差异有关。
嗜盐微生物因具有特殊的嗜盐和耐盐机制而具有广泛的应用。为了在高盐环境下适应生存,嗜盐或耐盐微生物能够合成并积累大量的相容物质,如甜菜碱、四氢嘧啶、羟基四氢嘧啶、谷氨酰胺、海藻糖和蔗糖等,以此维持细胞内外的渗透平衡[41]。四氢嘧啶是一种环状氨基酸衍生物,具有亲水性和两性离子特征,是典型的微生物相容溶质之一。研究证明四氢嘧啶具有强大的生物大分子(胞外多糖、酶、DNA和抗体等)和细胞稳定作用,能够保护细胞免受高渗透压及高温等极端环境的影响[42],因此在精细化工、环境治理、农业生物技术和生物医学等领域具有良好的应用潜力[10,43-47]。在盐激条件下,不同种属的嗜盐菌能生物合成四氢嘧啶,如 Marinococcus sp. M AR 2[48]、Stenotrophomonas sp. WMA-LM19[49]、 芽 孢 杆 菌 B. clausii NIOTDSB04[50]、盐单胞菌 H. salina[51]、H. hydrothermalis Y2[52]和 H. elongata[53]等。艾丽等[54]通过高效液相色谱-质谱法(HPLC-MS)测定Chromohalobacter S2在最适盐浓度下的合成量为238. 201 mg/g,姚倩倩等[55]通过HPLC测定Halomonas W2在高盐度下四氢嘧啶合成量为161 mg/g,王慧敏等[16]通过HPLC-MS测定Halomonas sp. Y在最适生长盐浓度下其四氢嘧啶合成量为175.5 mg/g。本研究采用普通摇瓶发酵初步测定柴达木盆地优势菌株的四氢嘧啶积聚量,发现X26、C15、X28、K11、X27、C11和X36等菌株四氢嘧啶积聚量明显高于其他菌株,其 含 量 分 别 为 369.10、324.69、296.56、275.20、239.51、220.56和208.56 mg/g。与以上菌株相比,本研究所筛选出的高产菌株四氢嘧啶积聚量相对较高,具有后续应用生产四氢嘧啶的潜力。
4 结论
本研究通过RM中盐和高盐培养基从三大硫酸镁亚型高盐盐湖中筛选分离获得嗜盐耐盐菌113株,16S rRNA基因测序显示优势种属为Bacillus、Staphylococcus和Halomonas。基于HPLC测定优势细菌胞内的四氢嘧啶积聚量发现菌株X26、X28、X27、X36、C15、K11和C11等菌株具有高效积聚生产四氢嘧啶的潜力,可用于四氢嘧啶的发酵模式菌株。本研究为柴达木盆地高盐盐湖微生物资源的开发和利用提供了一定的参考依据。
猜你喜欢
昆明医科大学学报(2021年8期)2021-08-13
昆明医科大学学报(2021年3期)2021-07-22
科学导报(2020年80期)2020-12-21
科学导报(2020年75期)2020-12-21
中国现代医药杂志(2020年10期)2020-12-14
农药科学与管理(2019年6期)2019-11-23
科学导报(2019年21期)2019-09-03
中成药(2017年9期)2017-12-19
中华老年口腔医学杂志(2016年2期)2017-01-15
环境(2016年12期)2017-01-12
