
濒危孑遗植物珙桐在不同水分条件下对增温和CO2浓度升高的响应
2022-03-07 12:27赵怀敏刘丽萍陶泉洪
绵阳师范学院学报 2022年2期
胡 钰,李 芯,赵怀敏,刘丽萍,陶泉洪,李 艳
(绵阳师范学院生态安全与保护四川省重点实验室,四川绵阳 621006)
0 引言
珙桐(DavidiainvolucrateBaill.)又名鸽子树、水梨子,隶属蓝果树科(Nyssaceae)珙桐属(Davidia),是我国特有的单型属植物[1].珙桐起源古老,为第三纪古热带植物区系的孑遗种,被誉为植物界的“活化石”[2].珙桐曾广泛分布在世界许多地区,但第四纪冰川致使国外珙桐已绝迹,现仅残存于我国,且仅分布在我国四川、云南、贵州、湖南、湖北、陕西与甘肃7省地形复杂的山地[3-4].珙桐分布区的气候条件相差较大,年平均气温8.9~15 ℃,平均相对湿度80%,年降水量600~2 600.9 mm[5].其多生长在山地沟谷及两侧山坡中、下部的小生境,对环境要求严苛,喜欢温凉、湿润、多雨、多雾的气候.
近年来,由于人类活动造成的温室气体排放不断增加,导致极端高温、干旱事件频繁发生.目前有关气候变化导致的极端天气,如高温、干旱等对珙桐的影响已有报道.研究发现,高温会对珙桐的光合作用PSI系统和PSII系统造成损伤,随着胁迫程度和胁迫时间的增加抗氧化酶清除氧自由基的能力也会减弱[6-7].珙桐对干旱也很敏感[8],干旱胁迫后其生长、光合叶面积、光合速率、气孔导度、蒸腾速率、叶绿素荧光、渗透调节物质积累、抗氧化酶活性等均会受到影响[9-13].
据预测,珙桐适宜的分布区域将受到气候变化的影响,分布范围将向我国西部及西南部地区扩展,同时目前的适宜分布区将破碎化[14-15].但有关气候变化的关键因子是如何影响珙桐生长的报道还较少.王宁宁等[9-16]发现,CO2浓度升高除了会影响珙桐分布外还会引起一些生理特性的改变,如叶片栅栏组织比例、叶绿素含量、光合作用效率等.
大气CO2浓度升高,增温以及全球温度升高导致干旱加剧都属于全球变化中的主要环境问题[8],而这些环境因子的共同作用对珙桐生长及生理特性的影响还不明确.本研究通过人工模拟大气CO2浓度升高和增温以及土壤干旱,以期分析在这些因子单独作用及共同作用下珙桐生长和光合特性的变化,为珙桐生理生态研究提供理论基础,也有助于认知珙桐对环境变化的响应机理和生物多样性保护.
1 材料与方法
1.1 材料与试验处理
试验材料是利用四川省绵阳市平武地区的珙桐种子繁殖的珙桐幼苗.实验在绵阳市平武县王朗国家级自然保护区(103°55′~104°10′E,32°49′~33°02′N)进行.于3月12日选取在苗圃中培育的健康无病虫害,30 cm左右,大小和高度均一的珙桐幼苗移栽到10 L的塑料花盆中,以土质均匀的当地土壤作为培养土,每盆栽1株.在大棚中缓苗2个月待长势稳定后,5月11日,选取144盆幼苗放入人工气候室中进行试验.
试验采用3因素完全随机设计:2种温度梯度×2种CO2浓度×2种水分梯度.采用人工气候室控制温度和CO2浓度,通过控制浇水时间来进行土壤干旱处理.其中2种温度梯度包括在环境温度下增温0 ℃ 和增温2.5 ℃;2种CO2浓度包括环境CO2浓度(350 ppm)和CO2浓度加倍(700 ppm);2种水分梯度包括每天浇水(非干旱)和隔5 d浇水(干旱)处理(干旱天数依据前期预实验结果,据称重法,干旱5 d土壤含水量达到田间持水量的50%).每个处理设3个重复,每个重复6株苗,试验从5月持续到8月底.
1.2 生长特征及生物量积累的测定
在试验结束时测定珙桐植株的株高和基径,并将所有植株按根、茎、叶分开收获,在105 ℃杀青1 h,75 ℃烘干至恒重后称重,然后计算叶片干重、茎干重、根干重、总生物量及根冠比.总生物量=根干重+茎干重+叶干重;根冠比=根干重/(茎干重+叶干重).
1.3 光合作用测定
在收获前3周,在每个处理中随机选择3株植株,每株植株选择3片完全伸展叶片用于测定叶片气体交换.采用LI-6400便携式光合作用测定系统(LI-CORInc. Lincoln, Nebr.)进行测定.测定时将叶面温度控制为25 ℃,光照强度控制为1 200 μmol m-2s-1,相对湿度控制为50%左右,CO2浓度控制为(350±5) μmolmol-1.测定时间选择晴朗上午08∶00-11∶30期间进行.测定指标包括Pn(净光合速率)、E(蒸腾速率)、gs(气孔导度)、Ci(胞间CO2浓度)等.
1.4 数据统计分析
数据采用SPSS16.0进行统计分析.采用两因素方差分析增温和CO2加倍及其共同作用分别在水分充足和干旱情况下对珙桐生长和光合作用的影响,用Duncan法在0.05水平上进行多重比较.采用成对T检验分析各气候因子处理下的生长和光合指标分别在水分充足和干旱下的差异.
2 结果与分析
2.1 增温和CO2加倍分别在非干旱和干旱条件下对珙桐生长的影响
株高在各处理下均无显著差异;水分充足条件下增温和CO2加倍的共同作用显著促进了基径增加,但在干旱条件下,其影响不显著(见表1).
2.2 增温和CO2加倍分别在水分充足和干旱条件下对珙桐生物量积累的影响
从表1可以看出,增温对各器官生物量及根冠比无显著影响,但CO2加倍能显著促进根系和总生物量的增加,并且在增温和CO2加倍的共同作用下两者增加也达到了显著水平,根细生物量的增加幅度在CO2加倍处理和增温与CO2加倍共同作用下分别为47.4%和65.1%,总生物量增加幅度分别为34.6%和46.5%;干旱条件下总生物量的增加在CO2加倍处理及两因子共同作用下分别显著提高了67.3%和47.1%,但干旱条件下根系生物量仅在CO2加倍处理下显著增加了77.7%,而在两因子共同作用下有增长趋势但未达到显著水平.

表1 珙桐生长特征在干旱和非干旱条件下对增温和CO2浓度加倍的响应
2.3 增温和CO2加倍分别在水分充足和干旱条件下对珙桐光合特性的影响
从表2可以看出,水分充足条件下,与对照相比,增温、CO2加倍及其共同作用均会显著增加净光合速率,并且CO2加倍后净光合速率最高,显著高于增温和交互处理;增温和CO2加倍处理后气孔导度会显著降低,但交互处理下又显著升高;胞间二氧化碳浓度和蒸腾速率在3种处理下均显著下降;干旱条件下,各处理对净光合速率均无显著影响,增温和CO2加倍显著降低、共同作用显著增加了气孔导度、胞间二氧化碳浓度和蒸腾速率.

表2 珙桐光合特性在干旱和非干旱条件下对增温和CO2浓度加倍的响应
2.4 生长特征在干旱与非干旱处理下的差异
由图1可知,各处理下珙桐的株高生长在干旱和非干旱条件下无显著差异,基径生长在单独干旱,增温、CO2加倍下干旱和非干旱处理也无显著差异,仅在增温和CO2加倍共同作用下,干旱处理显著抑制了基径生长.

图1 干旱和非干旱处理下珙桐生长特征差异
2.5 生物量积累在干旱与非干旱处理下的差异
由图2可知,单独干旱处理以及增温下的干旱处理不会对根系生物量产生显著影响,但在CO2加倍和增温与CO2加倍共同作用这两种条件下,干旱会显著抑制根系生物量积累.单独的干旱处理以及增温和CO2加倍的交互效应会显著影响叶片干重和总生物量的积累.根冠比在各种处理下,干旱和非干旱处理的差异都不显著.
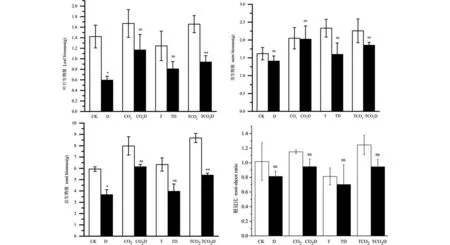
图2 干旱和非干旱处理下珙桐生物量积累差异
2.6 光合特性在干旱与非干旱处理下的差异
由图3可知,光合特性受水分胁迫影响较大,净光合速率,气孔导度,胞间二氧化碳浓度,蒸腾速率在单独干旱、增温、CO2加倍、增温与CO2共同处理下,水分胁迫的影响都达到了显著水平,且气孔导度和蒸腾速率达到极显著水平.

图3 干旱和非干旱处理下珙桐光合特性的差异
3 讨论
已有研究表明气候变化会对珙桐分布产生影响.吴建国等[15]利用分类和回归树生态位模型模拟分析了气候变化对珙桐分布的影响,认为随气候变化珙桐的适宜分布范围将减小且被破碎化.因此气候变化有可能改变珙桐目前分布区的环境条件,进而影响珙桐的生存.但是气候变化导致的干旱、温度和CO2浓度升高等对珙桐生长和生理特征的影响研究还较少.胡晓健等[17]发现,干旱胁迫会抑制马尾松株高的增加和生物量的积累;但本文中珙桐的株高和生物量在干旱条件下与非干旱条件下无显著差异.这可能是由于本研究中的干旱胁迫程度较轻.李娜等[18]发现樟子松的株高随着温度的升高而降低;而本研究发现升温对珙桐的株高没有明显的影响;这可能是由于在本研究中的增温梯度还较小.王宁宁等[16]发现,CO2浓度升高会显著抑制珙桐株高的增长,但显著促进了生物量的增加;提高了净光合速率.本研究结果中株高受CO2浓度升高的影响不明显,生物量的变化趋势与以往研究一致.同时本研究还发现总生物量的增加主要由于根系生长的增大.这与陈明涛等[19]关于干旱对油松,刺槐,侧柏和沙棘的研究结果是一致的,干旱胁迫会增加根生物量的积累.光合能力是植物生长和物质积累的重要影响因素[20-21],本研究中,增温、CO2浓度升高以及两者的共同作用均会显著提高珙桐叶片的净光合速率,这与CO2单独处理及与增温共同作用下珙桐生物量的增加趋势是一致的.增温对植物生长的影响以往研究结果并不一致[22-24].如增温会促进植物地上部的生长,促进枫树根系的生物量积累,但对农作物却有抑制作用[25-26].本研究中增温对珙桐生长和生物量积累的影响不显著,这可能与增温方式对土壤环境影响或者与植物间碳积累和分配方式的差异有关[22,27-28]
据预测,由于大气CO2浓度的升高引发的一些非生物环境胁迫,如热胁迫、干旱等会与CO2浓度升高一起产生联合生态效应[29-30].本研究中,单独的干旱处理会显著抑制珙桐的光合作用,这与以往研究一致[16],即干旱会通过影响气孔导度,蒸腾速率以及其他与光合相关的特性如光合色素等来对净光合速率产生影响[31-32].从生长和生物量积累来看,单独的干旱对珙桐的影响主要体现在对地上部分叶片生物量的积累上,对地下根系和茎干生物量积累以及生长上影响都不显著.但是在气候因子作用下,即增温和CO2升高的联合作用下,干旱对珙桐的影响会增加,如根系、叶片、总生物量、基径都会受到显著影响.其中,在气候因子影响下,如增温,干旱影响加重,可能是因为温度升高影响土壤的理化性质和土壤酶的活性,进而加重了干旱的胁迫[33].同时有研究证明,温度会通过改变植物体内代谢状况进而影响植物生长发育,温度过高会导致光合作用下降,使得呼吸作用消耗更多的碳水化合物,进而碳水化合物含量减少,促使干旱胁迫加重[34-36].因此,在单独干旱条件下,通过降低光合产物向叶片分配来抑制叶片生长从而减少地上部分水分蒸腾可能是珙桐应对水分胁迫的主要策略.但是当气候因子发生变化后,珙桐的地上和地下应对干旱的策略会发生变化.从各个气候因子分别作用下珙桐的生长反应来看,增温和CO2升高的影响与水分条件无关,但增温和CO2的联合作用对生长和光合的影响程度不同.与水分充足条件相比,干旱环境下增温和干旱的联合促进效应会降低,即干旱会在一定程度上影响气候因子对珙桐生长和光合的调控作用.
4 结论
据预测,珙桐的适宜分布区在未来气候变化下会逐渐往半干旱地区迁移,珙桐可能面临气候变化及由此带来的极端气候如干旱等对其生存能力的挑战.本文研究不同水分条件下,增温和CO2浓度升高对珙桐生长和光合作用的影响.结果表明,增温对珙桐幼苗生长和生理的影响不明显,且与水分条件无关;水分充足时,CO2浓度升高主要影响地上部叶片物质积累,但在增温和CO2浓度升高共同作用下,显著促进珙桐幼苗的地上部和地下部分生长;而在干旱条件下,CO2浓度升高及其与增温共同作用对珙桐生长的促进作用会降低.另外,在增温和CO2升高共同作用下干旱与非干旱对珙桐光合特性、根干重、叶片干重、总生物量以及基径生长的影响存在显著差异.
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
农业工程学报(2022年10期)2022-08-22
中原商报·科教研究(2022年1期)2022-05-13
农业工程学报(2022年4期)2022-04-24
现代园艺(2022年6期)2022-04-08
农业与技术(2020年12期)2020-07-04
安徽农学通报(2020年7期)2020-05-26
新课程·下旬(2017年7期)2017-08-14
红岩(2015年3期)2015-11-28
高中生学习·高三版(2014年3期)2014-04-29
