
Physiological Response to High Temperature and Evaluation of Heat
2022-03-07 00:03JiuyunWUGuixiangXUWeijiaLIANYaCHENHaifengLIYongxiangLIUJianfuJIANGJinghuiWEN
农业生物技术(英文版) 2022年1期
Jiuyun WU Guixiang XU Weijia LIAN Ya CHEN Haifeng LI Yongxiang LIU Jianfu JIANG Jinghui WEN


Tolerance of Different Grape Cultivars
Abstract [Objectives] This study was conducted to investigate the differences in the physiological responses of different grape cultivars to high temperature.
[Methods]The 19 tested cultivars were selected from the grape germplasm resources pool of Turpan Research Institute of Xinjiang Academy of Agricultural Sciences. Twelve physiological indexes including gas exchange parameters, chlorophyll content, antioxidant enzyme activity and proline content were determined in grape leaves under field conditions during the middle period of local natural high temperature period (July, daily maximum air temperature>35 ℃). The heat tolerance of different cultivars was evaluated by fuzzy membership function analysis and optimum partitioning clustering of ordered samples.
[Results] ① Under natural high temperature conditions in Turpan, the 19 tested grape cultivars responded differently to high temperature. ‘Red Globe’, ‘Fujiminori’ , ‘Beta’, ‘Hetianhuang’ had strong heat tolerance, while ‘Thompson Seedless’, ‘Hongqi Tezaomeigui’ , ‘Shuijing Wuhe’, ‘Victoria’ , ‘Yatomi Rosa’ and ‘Crimson Seedless’ had weak heat tolerance. ② Among the 12 physiological indexes, malondialdehyde content and antioxidant enzyme activity were mostly different among various grape cultivars. The grape cultivars with strong heat tolerance, ‘Red Globe’ and ‘Fujiminori’, had relatively lower malondialdehyde contents, while ‘Beta’ and ‘Hetianhuang’ had relatively higher malondialdehyde contents. But they had higher activity of antioxidant enzymes. ③ The results of fuzzy membership function analysis showed that the cumulative membership value (AR) of each physiological index was consistent with its apparent heat tolerance performance, suggesting that AR can be a potential index for the evaluation of heat tolerance of grape cultivars. Further cluster analysis classified the tested cultivars as strong, medium and weak. ‘Red Globe’, ‘Fujiminori’, ‘Beta’ and ‘Hetianhuang’ had strong heat tolerance.
[Conclusions]This study provides a reference for grape cultivation under high temperature and stress and breeding of heat-tolerant varieties.
Key words Grape; Physiological index; High temperature response; Heat tolerance
Received: September 6, 2021 Accepted: November 10, 2021
Supported by Youth Science and Technology Backbone Innovation Ability Cultivation Project of Xinjiang Academy of Agricultural Sciences (xjnkq-2021010); Tianshan Youth Project in Xinjiang Uygur Autonomous Region (2018Q093, 2019Q091); Tianshan Innovation Team of Xinjiang Uygur Autonomous Region (2020D14033); Natural Science Foundation of Xinjiang Uygur Autonomous Region (2019D01B33); Tianchi Plan of the Autonomous Region for Introduction of High-level Talents (2018).
Jiuyun WU (1988-), male, P. R. China, associate researcher, devoted to research about research on collection, preservation, identification and evaluation of grape germplasms.
*Corresponding author. E-mail: jianfu501@163.com; Wj51777@126.com.
High temperature will cause grape leaves to curl, lose water and dry up, and reduce photosynthetic efficiency, which in turn affects the physiological and biochemical reactions and the expression and regulation of stress-induced genes, severely impairs the ability to regulate their own stress, and causes heat damage and even malaise death[1-4]. With the continuous increase of global average temperature[5], the frequency and duration of high temperature events continue to increase, and the impact on grape production is becoming more and more significant. Turpan, Xinjiang has become an important grape-producing area in China due to its rich light and heat resources, and the area of viticulture has reached 36 800 hm2. However, the special geographical environment has caused Turpan to be affected by high temperature for a long time. The weather over 35 ℃ exceeds 100 d every year, and the weather over 40 ℃ averages more than 35 d per year[6-8]. The continuous high temperature stress seriously affects the photosynthesis of grapes, resulting in the reduction of grape yield and quality. Therefore, it is urgent to understand the effects of high temperature on the physiological characteristics of grapes, so as to provide a reference for alleviating high temperature stress, exploring stress-resistant cultivation methods and breeding heat-resistant grape varieties.
Scholars have done some research on the effects of high temperature on the physiological characteristics of grapes, and found that photosynthesis is the most sensitive physiological process of plants to high temperature stress[9]. High temperature stress will damage the photosynthetic apparatus of grape leaves, affect the photosynthetic efficiency of plants[10], and cause the Pn of grape leaves to decrease significantly[11]. High temperature stress can also disrupt the photochemical reaction and oxidation-reduction balance of grape leaves, causing grape leaves to lose water and chlorophyll fluorescence parameter values to be abnormal, and plants will show significant damage symptoms[12-14]. Meanwhile, high temperature will destroy the balance of active oxygen production and removal in plants, thereby producing a large amount of active oxygen and harmful substances such as malondialdehyde[15-18], but plants can reduce the damage of free radicals to cells caused by high temperature stress by improving their own antioxidant enzyme activity[19-20].
However, in general, there are few research reports on the effects of natural high temperature environmental conditions on the physiological characteristics of grapes, and previous studies have certain limitations. For example, adopting indoor simulation of high temperature stress conditions, or a small number of grape varieties, cannot accurately reflect the change characteristics of grape physiological characteristics under high temperature environmental conditions. Moreover, high temperature and strong light can synergistically inhibit the function of PSII[21-22], and only measuring the leaves indoors cannot fully reflect their heat tolerance in the natural environment[23]. Therefore, in this study, based on previous studies[24-31], under unique natural high temperature and strong light conditions in Turpan, the responses of physiological characteristics of different grape cultivars to high temperature stress were investigated, and their heat tolerance was comprehensively evaluated, with a view to providing a reference for the research on the heat tolerance mechanism of grape and the breeding of heat-tolerant grape cultivars in Turpan production area.
Materials and Methods
Experimental materials
The materials of 19 representative grape cultivars came from the grape germplasm resources pool of Turpan Research institute of Agricultural Sciences, Xinjiang Academy of Agricultural Sciences. The resource nursery is located at 89°18′E, 42°53′N, altitude -50 m, and the names, cultivar characteristics and parent sources of the 19 grape cultivars are shown in Table 1. The scaffolding adopts a dragon-trunk shape with a height of 1.5 m. The rows had a spacing of 1.0 m×4.5 m, and they were in the east-west direction. Ditch irrigation was adopted. The trees were at ages of 8-14 years, and had moderate tree vigor. The management was relatively extensive. The local annual average temperature was 17.6 ℃; the annual precipitation was 12.5 mm; the annual sunshine hours were 3 109. 2 h; and the frost-free period was about 210 d.
Determination indexes and methods
Temperature and humidity monitoring during the high-temperature period
The experiment was conducted during the high temperature period (June-August) in Turpan for two consecutive years from 2017 to 2018. MicroLabLite U disk thermometers and L92 digital temperature and humidity meters were used to monitor temperature and humidity, which were measured and recorded every half an hour, continuously in the entire high temperature period (June to August).
Leaf morphology observation
The experiment was conducted in the middle of the high temperature period (July) in Turpan for two consecutive years from 2017 to 2018. Plant growth was observed in field, and grape leaves were also picked for observation. For each species, the fifth functional leaves of new shoots with basically the same growth were selected and quickly brought back to the laboratory to take pictures and observe.
Photosynthetic gas exchange parameters
The photosynthetic characteristics were determined for two consecutive years in Turpan in the middle of the high temperature period (July). The fifth functional leaves of new shoots with the same growth irradiated by the sun directly were randomly selected for each species, and the photosynthetic rate (Pn), stomatal conductance (Gs), intercellular CO2 concentration (Ci), transpiration rate (Tr) and other indexes were measured. For each species, three leaves were measured, and three replicates were set. The measurement conditions were as follows photon flux density 1 800 μmol/(m2·s), CO2 concentration of the reference chamber (360±20) μmol/L, and leaf chamber temperature (25±0.5) ℃. The day of the measurement was clear and windless, and the measurement period was 13:00-16:00, during which the average temperature was 41.20 ℃ and the relative humidity was 36.30%.
Chlorophyll content
The relative content of chlorophyll was measured for two consecutive years in Turpan in the middle of the high temperature period (July) by a TYS-B chlorophyll meter, and expressed as SPAD value. The fifth functional leaves of new shoots with the same growth irradiated by the sun directly were randomly selected for each species, and the data displayed by the chlorophyll meter were recorded. Each cultivar was measured for five times, and three replicates were set. The measurement period was 11:00-13:00, during which the average temperature was 38.01 ℃ and the average relative humidity was 35.31%.
Physiological indicators of stress resistance
Sampling was conducted from 11:00 to 13:00 on July 14, 2017, during which the average temperature was 35.66 ℃ and the relative humidity was 36.66%. The 5th functional leaves of new shoots were picked and added into a sampling bag, and three leaves were taken for each cultivar. The samples were placed in an incubator, covered with ice cubes, and quickly brought back to the laboratory. Next, 1.0 g of grape leaf tissues was immediately weighed accurately, added with 9.0 ml of normal saline in portions according to the ratio of weight (g)∶volume (ml)=1∶9, and added into a mortar, into which a little quartz sand was then added. The leaf tissues were mechanically grinded into a homogenate under ice-water bath conditions, poured into a 10 ml centrifuge tube, and stood in an ice-water bath for 15 min. Centrifugation was then performed at 3 500 r/min, 4 ℃ for 10 min, and the supernatant was taken for testing. Proline (Pro) content was determined by the proline determination kit (colorimetric method); protein concentration was determined by the BCA protein quantification kit; and SOD, POD, CAT and APX activity were determined using superoxide dismutase assay kit (colorimetric method), peroxidase assay kit (colorimetric method), catalase assay kit (colorimetric method) and ascorbate peroxidase assay kit (colorimetric method), respectively. TBA malondialdehyde determination kit (colorimetric method) was used to determine the content of MDA.
Heat resistance evaluation
The fuzzy membership function method[31-35] was adopted for the analysis on the physiological indexes of different varieties of grapes, and for the calculation of the membership value of each index separately. The formulas were as follows:
R(Xi)=Xi-XminXmax-Xmin (1)
R(Xi)=-1Xi-XminXmax-Xmin (2)
AR=∑ni=1Xi (3)
In the formulas: Xi is the measured value of each index, and Xmax and Xmin are the maximum and minimum values of a certain index, respectively. If an index trait is positively correlated with heat resistance, formula (1) can be used, and if it is negatively correlated, formula (2) can be used; and formula (3) was used to accumulate the membership values of various indexes, and heat resistance evaluation was performed combining cluster analysis[31].
Data processing
Analysis of variance was performed on the test data. Multiple comparisons were performed by LSD, and the significance of differences was detected with P<0.05 as the significant level of difference, and P<0.01 as the extremely significant level of difference. DPS 7.05 and SPSS 19.0 were used for correlation analysis and cluster analysis.
Results and Analysis
Dynamics of temperature changes during the high temperature period of the experimental site
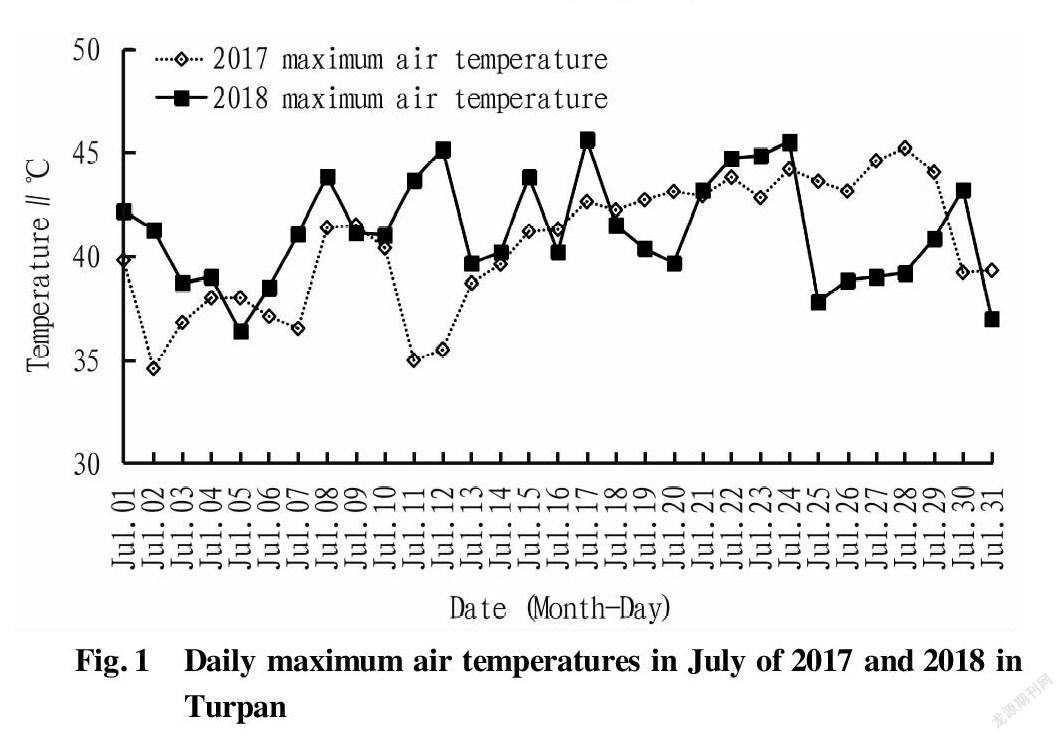
From 2017 to 2018, the temperature in the experimental area was monitored during the high temperature period (June to August) for two consecutive years, and the physiological indexes were measured in the middle of the high temperature period (July). In terms of the temperature, the average temperature in July 2017 was 33.15 ℃, the average daily high temperature was 40.60 ℃, and the highest temperature was 45.20 ℃. In July 2017, Turpan had a daily maximum temperature over 35 ℃ for up to 30 d, and the weather with a temperature over 35 ℃ lasted for 29 d, including 18 d for weather over 40 ℃ and the continuous period of 15 d with a weather over 40 ℃ (Fig. 1). The average temperature in July 2018 was 34.23 ℃, the average daily high temperature was 41.19 ℃, and the highest temperature was 45.61 ℃. In July 2018, the daily maximum temperatures of Turpan were all higher than 35 ℃, and 20 d had a weather over 40 ℃ (Fig. 1).
Jiuyun WU et al. Physiological Response to High Temperature and Evaluation of Heat Tolerance in Different Grape Cultivars
Phenotypic changes in grape leaves of different cultivars under high temperature conditions
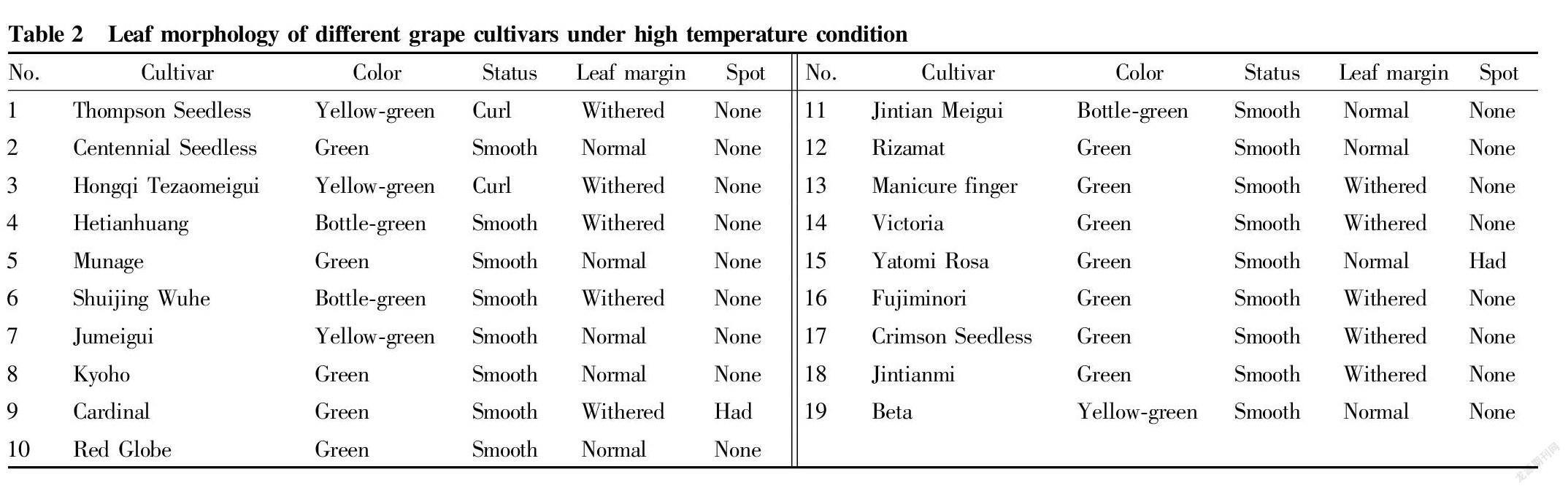
Under the natural high temperature conditions in Turpan, only the leaves of ‘Red Globe’, ‘Centennial Seedless’, ‘Munage’ and ‘Rizamat’ had no obvious heat damage symptoms, and the leaves of other cultivars had different degrees of heat damage symptoms. Among them, the leaves of ‘Thompson Seedless’, ‘Jumeigui’ and ‘Beta’ were yellow-green; the leaves of ‘Hetianhuang’, ‘Shuijing Wuhe’ and ‘Jintian Meigui’ were dark green; the leaves of ‘Thompson Seedless’ and ‘Hongqi Tezaomeigui’ were curled; ‘Thompson Seedless’, ‘Hongqi Tezaomeigui’, ‘Hetianhuang’, ‘Shuijing Wuhe’, ‘Cardinal’, ‘Manicure finger’, ‘Fujiminori’, ‘Crimson Seedless’, ‘Jintianmi’ and ‘Victoria’ had different degrees of dry leaf margin, and the leaf margins of ‘Hetianhuang’, ‘Cardinal’, ‘Crimson Seedless’ and ‘Jintianmi’ were severely dry, and the leaf margins of ‘Hongqi Tezaomeigu’ , ‘Shuijing Wuhe’, ‘Manicure finger’, ‘Fujiminori’ and ‘Victoria’ were slightly dry; and the leaves of ‘Cardinal’ and ‘Yatomi Rosa’ showed spots. The results of morphological observation showed that the heat damage symptoms of ‘Thompson Seedless’, ‘Shuijing Wuhe’ and ‘Hongqi Tezaomeigui’ were more obvious, and they were preliminarily judged to be high-temperature sensitive cultivars, and ‘Red Globe’ and ‘Rizamat’ and ‘Centennial Seedless’ were cultivars tolerant to high temperature.
Comparison of photosynthetic gas exchange parameters of different grape cultivars under high temperature conditions
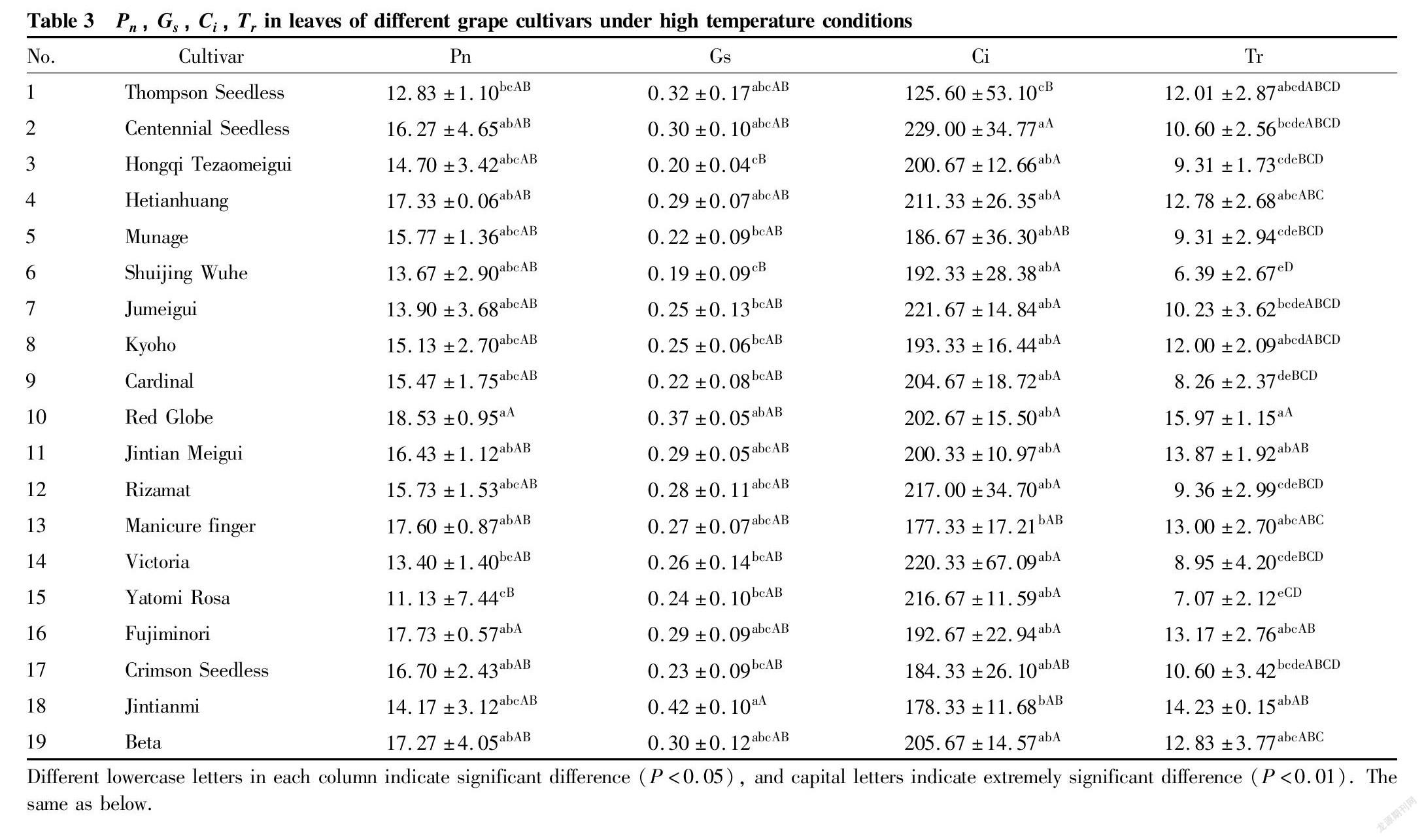
Photosynthesis is the basis for the formation of grape yield and is closely related to factors such as net photosynthetic rate, stomatal conductance, intercellular CO2 concentration, and transpiration rate. It can be seen from Table 3 that under natural high temperature conditions in Turpan, the net photosynthetic rate (Pn) was the highest in ‘Red Globe’ [18.53 μmol/(m2·s)] and the lowest in ‘Yatomi Rosa’ [11.13 μmol/(m2·s)] among the 19 grape cultivars. Except for ‘Yatomi Rosa’, which was extremely significantly lower than ‘Fujiminori’ and ‘Red Globe’ (P<0.01), there were no significant differences in Pn between other varieties (P>0.05). The stomatal conductance (Gs) was the highest in ‘Jintianmi’ [0.42 mol/(m2·s)] and the lowest in ‘Shuijing Wuhe’ (0.19 mol/(m2·s)) among various grape cultivars, and except for the Gs of ‘Jintianmi’, which was extremely significantly higher than those of ‘Shuijing Wuhe’ and ‘Hongqi Tezaomeigui’ (P<0.01), there were no significant differences in Gs between other cultivars. Among various grape cultivars, ‘Centennial Seedless’ had the highest intercellular CO2 concentration (Ci) (229.00 μmol/mol), ‘Thompson Seedless’ showed the lowest value (125.60 μmol/mol), which was significantly lower than other 18 cultivars (P<0.05); ‘Jintianmi’ and ‘Manicure finger’ were significantly lower than ‘Centennial Seedless’ (P<0.05); and there were no significant differences in Ci between other cultivars. In addition, the transpiration rate (Tr) was the highest in ‘Red Globe’ [15.97 mol/(m2·s)] and the lowest in ‘Shuijing Wuhe’ [6.39 mol/(m2·s)] among various grape cultivars, and the values of ‘Shuijing Wuhe’ and ‘Yatomi Rosa’ were extremely significantly lower than those of ‘Red Globe’, ‘Jintianmi’ and ‘Jintian Meigui’ (P<0.01). It showed that under the natural high temperature conditions in Turpan, ‘Red Globe’, ‘Hetianhuang’, ‘Fujiminori’ and ‘Beta’ among the 19 varieties still maintained relatively high photosynthetic efficiency, while ‘Shuijing Wuhe’ and ‘Thompson Seedless’ and ‘Yatomi Rosa’ had a low photosynthetic efficiency.
Comparison of chlorophyll and malondialdehyde contents in leaves of different grape cultivars under high temperature conditions
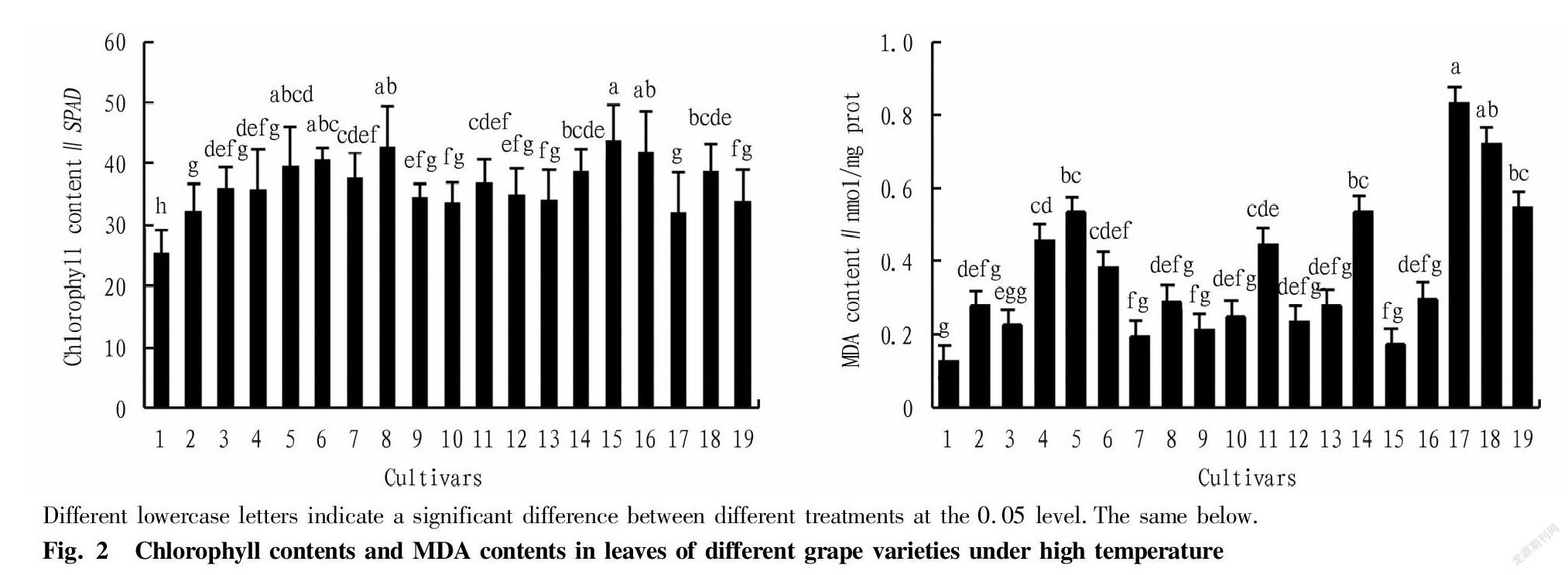
Chlorophyll content directly reflects the photosynthetic efficiency of plants, and is one of the important indexes of changes in their photosynthetic physiological activity. Fig. 2 shows that among the 19 grape cultivars under natural high temperature conditions in Turpan, ‘Yatomi Rosa’, ‘Kyoho’ and ‘Fujiminori’ had higher chlorophyll contents, which were significantly higher than those of other cultivars except ‘Victoria’, ‘Cardinal’ and ‘Centennial Seedless’ (P<0.05). Among them, ‘Yatomi Rosa’ had the highest chlorophyll content of 43.85 SPAD, and ‘Thompson Seedless’ was the lowest with only 25.44 SPAD, which was extremely significantly lower than other 18 cultivars (P<0.01), reflecting that its photosynthetic efficiency significant was lower than other cultivars under high temperature conditions.
It can also be seen from Fig. 2 that under natural high temperature conditions in Turpan, "Crimson Seedless", "Jintianmi" and "Beta" among different grape cultivars had higher leaf malondialdehyde (MDA) contents, which were extremely significantly higher than those of other cultivars except ‘Victoria’ and ‘Munage’ (P<0.01). Among them, the content of MDA in ‘Crimson Seedless’ was the highest (0.833 nmol/mg), and the content of "Thompson Seedless" was the lowest (0.127 nmol/mg), and significantly lower than those of ‘Crimson Seedless’, ‘Jintianmi’, ‘Beta’, ‘Victoria’, ‘Munage’, ‘Hetianhuang’, and ‘Jintian Meigui’ (P<0.01), indicating that different cultivars had different degrees of membrane lipid peroxidation under high temperature conditions, and the degree of membrane lipid peroxidation of ‘Thompson Seedless’ among them was significantly lower than above seven cultivars (Fig.2).
Comparison of soluble protein and proline contents of different grape cultivars under high temperature conditions
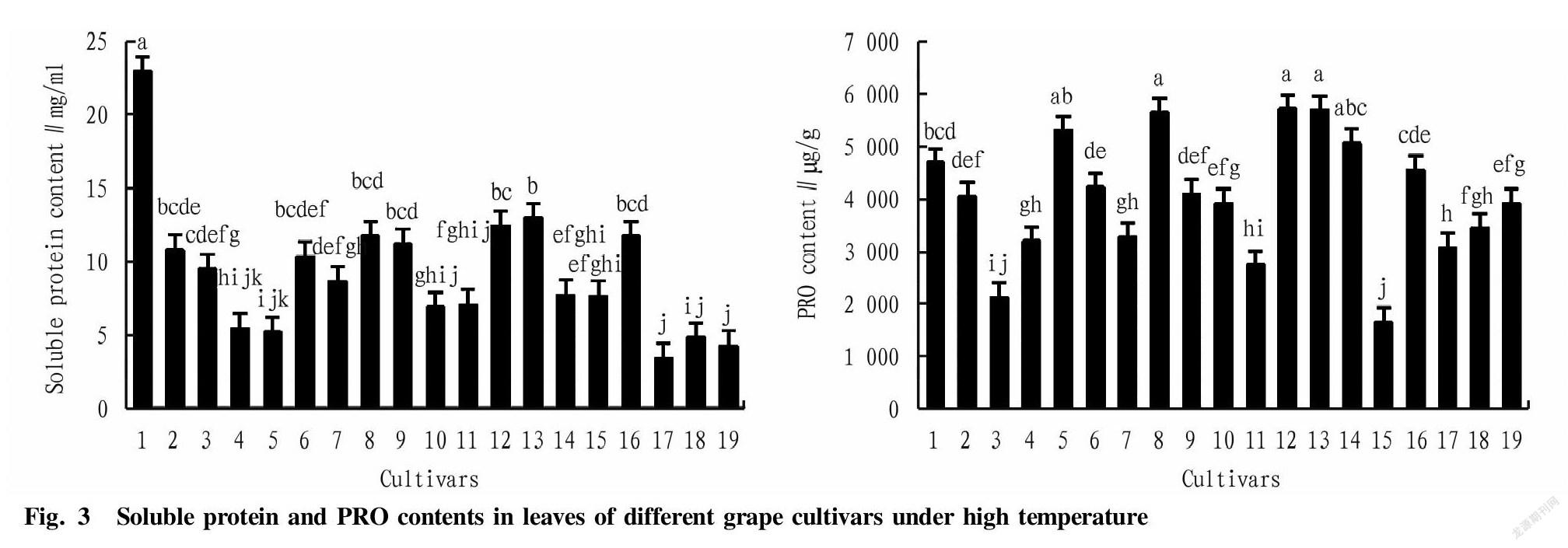
It can be seen from Fig. 3 that under natural high temperature conditions in Turpan, ‘Thompson Seedless’, ‘Manicure finger’ and ‘Rizamat’ had higher leaf soluble protein contents, which were extremely significantly higher than those of ‘Crimson Seedless’, ‘Beta’, and ‘Jintianmi’ (P<0.01). Among them, ‘Thompson Seedless’ had the highest protein content (22.903 mg/ml), which was significantly higher than other varieties (P<0.01), and ‘Crimson Seedless’ had the lowest content of soluble protein (3.418 mg/ml), which was extremely significantly lower than other varieties. Meanwhile, ‘Rizamat’, ‘Manicure finger’, and ‘Kyoho’ among various grape cultivars showed higher proline contents, which were significantly higher than those of other varieties except ‘Munage’ and ‘Victoria’ (P<0.05). Among them, ‘Rizamat’ had the highest proline content (5 725.0 μg/g), and ‘Yatomi Rosa’ had the lowest content (1 650.0 μg/g), which was extremely significantly lower than other varieties (P<0.01). It showed that the osmotic adjustment ability of cultivars such as ‘Thompson Seedless’, ‘Manicure finger’ and ‘Rizamat’ was relatively strong under high temperature conditions, while the osmotic adjustment ability of cultivars ‘Yatomi Rosa’, ‘Crimson Seedless’ and ‘Hongqi Tezaomeigui’ was weak.
Comparison of antioxidant enzyme activity in leaves of different grape cultivars under high temperature conditions
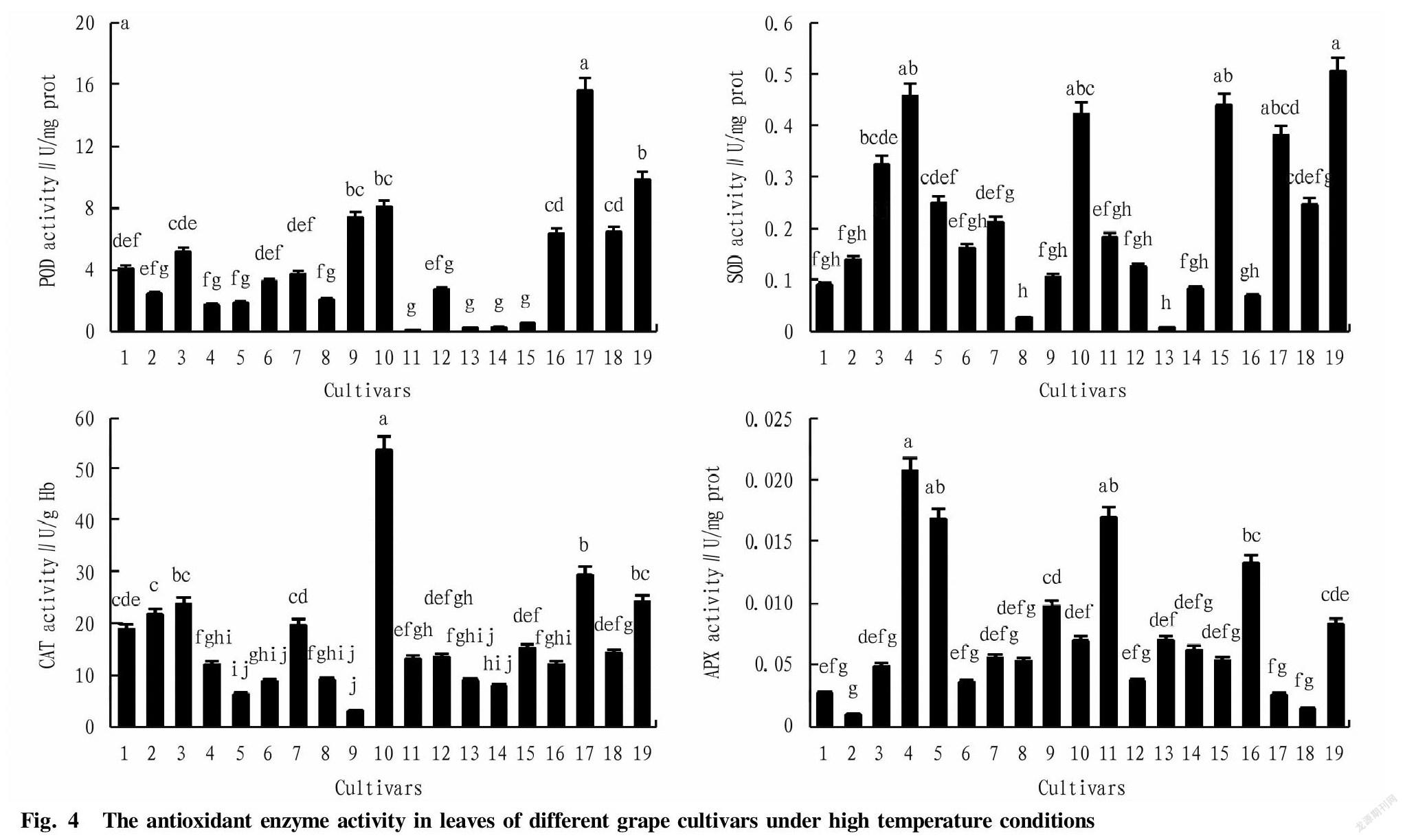
Fig. 4 shows that among the different grape cultivars under natural high temperature conditions in Turpan, ‘Crimson Seedless’ and ‘Beta’ had higher leaf POD activity, which was significantly higher than that of other cultivars except for ‘Red Globe’ and ‘Cardinal’ (P<0.05). Among them, the activity of ‘Crimson Seedless’ was the highest (15.635 U/mg), and was significantly higher than that of other cultivars (P<0.01), and ‘Jintian Meigui’, ‘Manicure finger’, ‘Victoria’ and ‘Yatomi Rosa’ had lower POD activity, which was significantly lower than that of other cultivars except ‘Hetianhuang’, ‘Munage’, ‘Kyoho’, and ‘Centennial Seedless’ (P<0.05), and ‘Jintian Meigui’ had the lowest activity (0.110 U/mg). Meanwhile, the SOD activity of grape leaves was higher in ‘Beta’, ‘Hetianhuang’, and ‘Yatomi Rosa’, which were significantly higher than other cultivars except ‘Red Globe’, ‘Jintianmi’, ‘Hongqi Tezaomeigui’ (P<0.05). Specifically, the SOD activity of ‘Beta’ was the highest (0.505 U/mg), and ‘Manicure finger’ had the lowest SOD activity (0.008 U/mg), which was significantly lower than most cultivars. In addition, the CAT activity of grape leaves was higher in ‘Red Globe’ and ‘Jintian Meigui’, of which ‘Red Globe’ had the highest CAT activity (53.527 U/g), which was significantly higher than that of other cultivars (P<0.01), and ‘Cardinal’ had the lowest CAT activity (3.082 U/g), which was extremely significantly lower than most cultivars (P<0.01). In terms of the APX activity of grape leaves, ‘Hetianhuang’, ‘Jintian Meigui’and ‘Munage’ were higher, extremely significantly higher than other cultivars except ‘Fujiminori’. ‘Hetianhuang’ had the highest APX activity (0.021 U/mg), and ‘Centennial Seedless’ showed the lowest value (0.001 U/mg), which was extremely significantly lower than ‘Hetianhuang’, ‘Jintian Meigui’, ‘Munage’, ‘Fujiminori’ and ‘Cardinal’. The above results indicated that under natural high temperature conditions in Turpan, ‘Red Globe’, ‘Beta’, ‘Crimson Seedless’ and ‘Hetianhuang’ had higher antioxidant enzyme activity and stronger ability to withstand high temperature stress.
Analysis and comprehensive evaluation of heat tolerance in grape cultivars
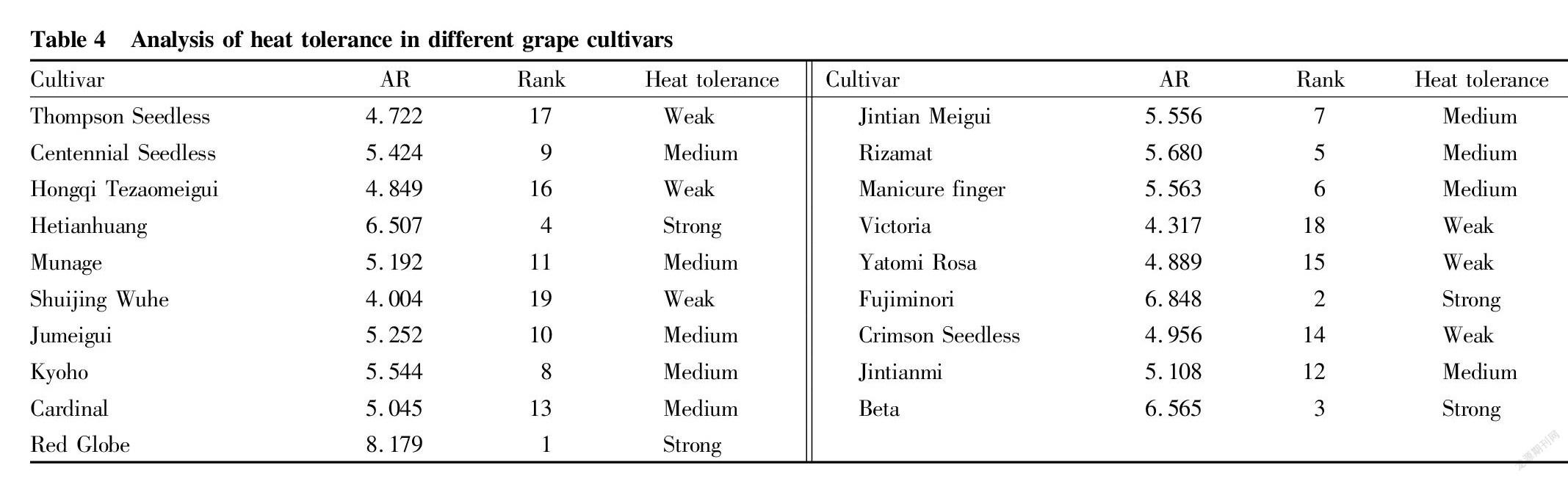
The 12 indexes were analyzed by the fuzzy membership function method, and the comprehensive AR values were sorted (Table 4). The result was ‘Red Globe’﹥‘Fujiminori’﹥‘Beta’﹥‘Hetianhuang’﹥‘Rizamat’﹥‘Manicure finger’﹥‘Jintian Meigui’﹥‘Kyoho’﹥‘Centennial Seedless’﹥‘Jumeigui’﹥‘Munage’﹥‘Jintianmi’﹥‘Cardinal’﹥‘Crimson Seedless’﹥‘Yatomi Rosa’﹥‘Hongqi Tezaomeigui’﹥‘Thompson Seedless’﹥‘Victoria’﹥‘Shuijing Wuhe’.
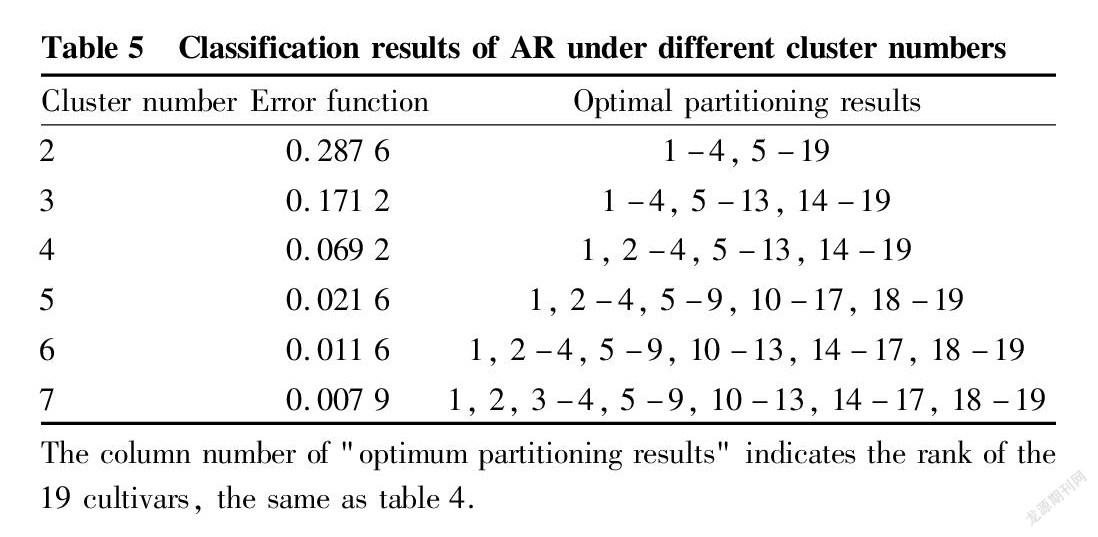
The optimum partitioning clustering method of ordered samples was further used to classify the AR values, so as to obtain the optimum partitioning error function and classification results of all cultivars for heat tolerance. As the number of classifications increased, the error function tended to be stable, and the samples were initially divided into three levels, and F-test is performed (Table 5). The results showed that the differences between various levels were extremely significant (P<0.01), so the optimum three-level partitioning was determined. The heat tolerance of the samples was divided into three types: weak, medium, and strong, with AR thresholds corresponding to <4.0, 4.0-6.0, and >6.0, respectively. According to this standard, the 19 cultivars were screened and evaluated. Finally, 4 cultivars had strong heat tolerance, 9 cultivars had medium heat tolerance, and 6 cultivars had weak heat tolerance. Among them, ‘Red Globe’, ‘Fujiminori’, ‘Beta’ and ‘Hetianhuang’ were cultivars with strong heat tolerance, which provides a theoretical reference for the selection and breeding of heat-tolerant grape cultivars.
Conclusions and Discussion
The response of plants to high temperature stress is affected by many factors, and the heat tolerance mechanisms of different types of plants are not the same[32]. Studies have shown that the optimal growth temperature for grape is 25-30 ℃[12, 36]. Some scholars believe that the temperature of 30-35 ℃ during the day has no significant effect on photosynthesis of grape, and only when the temperature exceeds 35 ℃ will it have a significant effect on photosynthesis of grape[36]. Therefore, in this study, we carried out the evaluation of high temperature response of physiological indexes and heat tolerance in different grape cultivars under natural high temperature conditions (>40 ℃) in Turpan, which has reference value for the research of grape heat-tolerant physiological defense mechanisms and the breeding of heat-tolerant varieties. The results showed that the photosynthetic efficiency and chlorophyll content of the four cultivars of ‘Red Globe’, ‘Hetianhuang’, ‘Fujiminori’ and‘Beta’ were relatively high, and they were evaluated comprehensively to have stronger heat tolerance, and were all cultivars with mid-late maturity and strong adaptability, providing reference for the selection and breeding of heat-tolerant grape varieties in Turpan.
Changes in the morphological characteristics of plants can directly reflect the degree of high temperature damage and resistance to high temperature stress[14]. Photosynthesis is the most sensitive physiological process of plants to high temperature[23-24], while chlorophyll is the main pigment for photosynthesis in plants, and its content directly reflects the nutritional status and photosynthetic capacity of plants. This study found that under natural high temperature conditions in Turpan, the photosynthetic efficiency and chlorophyll content were basically consistent with the apparent morphology of leaves, similar to our previous research results[8]. In this study, it was also found that the net photosynthetic rate and chlorophyll content of the leaves of ‘Thompson Seedless’ under high temperature conditions were significantly lower than other cultivars, but it can still be grown and promoted in Turpan for a long time. We speculate that it might be related to the high electron transfer rate, osmotic adjustment ability and strong self-protection of ‘Thompson Seedless’ under high temperature conditions[8, 30], which is worthy of further exploration and research.
Soluble protein and proline have the functions of osmotic regulation and cytoplasmic dehydration prevention, which can reflect the stress resistance of plants[23, 28]. MDA content represents the degree of membrane lipid peroxidation and indirectly reflects the antioxidant capacity of plant tissues[23]. High temperature stress will increase the peroxidation of plants and cause the increase of MDA content, and plants maintain the stability of the membrane system by enhancing the activity of antioxidant enzymes and increasing the content of soluble protein and proline[14, 23]. The results of this study showed that under the unique natural high temperature conditions in Turpan, the antioxidant enzyme activity of the leaves of the 19 grape cultivars was basically consistent with their photosynthetic efficiency, but it was inconsistent with their MDA contents. We believe that it might be related to protein concentration and enzyme protein inactivation, which is consistent with the results of Cha et al.[14].
The response of different physiological indexes to high temperature stress is very different, and the reflected information has a certain relevance to the evaluation of heat tolerance. Li[34] and Jiang[31] adopted fuzzy membership function and cluster analysis to evaluate the heat tolerance of different varieties of plants, and all obtained satisfactory results. Therefore, in this study, we adopted fuzzy membership function combined with the optimum partitioning clustering method to comprehensively evaluate the heat tolerance of different grape cultivars[31-36], and the results were basically consistent with field morphology of the plants. Other studies have shown that strong light and high temperature can synergistically inhibit the function of plant PS II[21-22], and only measuring the leaves indoors cannot fully reflect their heat tolerance in the natural environment[27]. The data of this study were obtained under natural high temperature and strong light environment, so the methods and the results obtained have certain reference significance for the evaluation, identification and variety breeding of grape.
References
[1] LIU JZ, FENG LL, GU XT, et al. An H3K27me3 demethylase-HSFA2 regulatory loop orchestrates transgenerational thermomemory in Arabidopsis[J]. Cell Research, 2019(29): 379-390.
[2] ZHANG SF, LING JC. Effects of high temperature stress on physiological property of grape leaf in greenhouse[J]. Qinghai Agricultural and forestry technology, 2017(2): 18-20. (in Chinese).
[3] ZHA Q, XI XJ, JIANG AL, et al. Influence of heat stress on the expression of related genes and proteins in grapevines [J]. Scientia Agricultura Sinica, 2017, 50(9): 1674-1683. (in Chinese).
[4] ZHANG RJ, LI Y, YU XM, et al. Effects of heat stress and exogenous brassinolide on photosynthesis of leaves and berry quality of ‘Kyoho’ grapevine[J]. Journal of Fruit Science, 2015, 32(4): 590-596. (in Chinese).
[5] IPCC. Climate change 2014: The physical science basis∥contribution of working group I to the fifth assessment report of the intergovernmental panel on climate change[M]. Cambridge: Cambridge University Press, 2014.
[6] CHEN H. Xinjiang statistical yearbook[M]. Beijing: China Statistical Publishing House, 2018. (in Chinese).
[7] Turpan Government Network. Access Turpan: Climate characteristics[EB/OL]. 2017-09-14 [2018-04-15].http:∥www.tlf.gov.cn/info/375/171173.htm. (in Chinese).
[8] WU JY, LIAN WJ, LIU ZG, et al. High temperature response of chlorophyll fluorescence parameter and heat tolerance evaluation of different grape cultivars[J]. Journal of Northwest A&F University (Natural Science Edition), 2019, 47(6): 80-88. (in Chinese).
[9] WALBOT V. How plants cope with temperature stress[J]. BMC Biol, 2011, 9 (1): 79.
[10] WANG LJ, FAN L, LOESCHER W, et al. Salicylic acid alleviates decreases in photosynthesis under heat stress and accelerates recovery in grapevine leaves[J]. BMC Plant Biol, 2010, 10(1): 34.
[11] LUO H B, MA L, DUAN W, et al.Influence of heat stress on photosynthesis in Vitis vinifera L. cv. Cabernet Sauvignon[J]. Scientia Agricultura Sinica, 2010(13): 2744-2750. (in Chinese).
[12] ZHA Q, XI XJ, JIANG AL, et al. Effects of heat stress on chlorophyll fluorescence characteristics and antioxidant activity in grapevines[J]. Plant Physiology Journal, 2016, 52(4): 525-532. (in Chinese).
[13] ZHA Q, XI XJ, JIANG AL, et al. Changes in the protective mechanism of photosystem II and molecular regulation in response to high temperature stress in grapevines[J]. Plant Physiol Biochem, 2016(101): 43-53.
[14] ZHA Q, XI XJ, JIANG AL, et al. Research on high temperature stress response of different table grape cultivars[J]. Acta Agriculturae Shanghai, 2018, 34(5): 77-83. (in Chinese).
[15] YANG XF, GUO FQ. Research advances in mechanisms of plant leaf senescence under heat stress[J]. Plant Physiol J, 2014, 50(9): 1285-1292. (in Chinese).
[16] WU HY, SHOU SY, ZHU ZJ, et al. Effect of high temperature stress on photosynthesis and chlorophyll fluorescence in sweet pepper (Capsicum fructescens L.)[J]. Acta Horiculturae Sinica, 2001, 28(6): 517-521. (in Chinese).
[17] SUN JL, ZHAO BL, YU SL. Effects of exogenous salicylic acid (SA) on membrane lipid peroxidation and the activity of antioxidase in grape seedlings under high temperature stress[J]. Journal of Shihezi University: Natural Science, 2015, 33(3): 275-280. (in Chinese).
[18] LI XH, LI HX, REN JP, et al. Effect of cadmium stress on photosynthesis, lipid peroxidation and antioxidant enzymes activities of grapevine with different scion-rootstock combinations[J]. Journal of Henan Agricultural Sciences, 2018, 47(3): 100-104. (in Chinese).
[19] SUN LX, XIE YL, ZHANG YH. Effect of temperature variation within the facilities on antioxidant properties grape leaves under early and dlayed cultivation[J]. Northern Horticulture, 2014(1): 38-43. (in Chinese).
[20] WANG BL, XU M, SHI QH, et al. Effects of high temperature stress on antioxidant systems, chlorophyll and chlorophyll fluorescence parameters in early cauliflower leaves[J]. Scientia Agricultura Sinica, 2004, 37(8): 1245-1250. (in Chinese).
[21] SUN YJ, FU YD, DU YP, et al. Effects of different temperature and light treatments on photosynthetic system II in Vitis vinifera L. cv. cabernet sauvignon[J]. Scientia Agricultura Sinica, 2013, 46(6): 1191-1200. (in Chinese). (in Chinese).
[22] SUN YJ, DU YP, ZHAI H. Effects of different light intensity on PSII activity and recovery of Vitis vinifera cv. cabernet sauvignon leaves under high temperature stress[J]. Plant Physiology Journal, 2014, 50(8): 1209-1215. (in Chinese).
[23] WANG T, TIAN XY, XIE YF, et al. Research advance on heat-stress tolerance in plants[J]. Journal of Yunnan Agricultural University, 2013, 28(5): 719-726. (in Chinese).
[24] JIN J, YANG L, FAN D Y, et al. Advances in the studies on high temperature tolerance of fruit trees[J]. Molecular Plant Breeding: 1-12. [2018-10-12].http://kns.cnki.net/kcms/detail/46. 1068.S.20180817.1643.008.html. (in Chinese).
[25] ZHA Q, XI XJ, JIANG AL, et al. Study on leaf phenotype, soluble sugar and chlorophyll fluorescence characteristics response to heat stress in ‘Jumeigui’ grapes[J]. Chinese Agricultural Science Bulletin, 2015, 31(25): 118-123. (in Chinese).
[26] STEFANOV D, PETKOVA V, DENEV ID. Screening for heat tolerance in common bean (Phaseolus vulgaris L.) lines and cultivars using JIP-test [J]. Scientia Horticulturae, 2011, 128(1): 1-6.
[27] SUN JL, ZHAO BL, YU SL. Study of exogenous salicylic acid (SA) on the heat tolerance in grape seedlings under high temperature stress [J]. Journal of Soil and Water Conservation, 2014, 28(3): 290-294. (in Chinese).
[28] XU HG. Evaluation of grape heat tolerance and study on the transcriptome of grape with different heat tolerance[D]. Beijing: China Agricultural University, 2014. (in Chinese).
[29] JIA YANG. Study on the microclimate difference of Turpan grape canopies and the photosynthetic characteristics of several grape varieties[D]. Urumqi: Xinjiang Agricultural University, 2015. (in Chinese).
[30] JIA Y, LIAO K, NIU YY, et al. Analysis on difference of photosynthetic characteristics and fluorescence parameters of different grape varieties in Turpan area[J]. Journal of Xinjiang Agricultural University, 2015, 38(1): 12-17. (in Chinese).
[31] JIANG JF, MA YF, FAN XC, et al. Evaluation of 196 Vitis germplasm resources to heat tolerance [J]. Journal of Plant Genetic Resources, 2017, 18(1): 70-79. (in Chinese).
[32] ZHA Q, XI XJ, HE YN, et al. Comprehensive evaluation of heat resistance in 68 Vitis germplasm resources[J].Vitis, 2018(57): 75-81.
[33] ZHANG Hl, YUAN C, ZHU L, et al. Comprehensive evaluation of salt-tolerance of maize at mature period using subordinate function value analysis[J]. Journal of Northwest A&F University: Natural Science Edition, 2018, 46(2): 47-55. (in Chinese).
[34] LI H, SU J L, LI C, et al. Physiological response and heat tolerance of different Rhododendron cultivars under high temperature stress[J]. Journal of Jiangsu Forestry & Technology, 2018, 45(5): 1-9. (in Chinese).
[35] WU JY. Evaluation of the heat tolerance of different grape varieties in Turpan[D]. Shihezi: Shihezi University, 2018. (in Chinese).
[36] LI C, LI XJ, HAN Z, et al. Research advances on effects of abiotic stress on photosynthesis of grape[J]. Shandong Agricultural Sciences, 2017, 49(12): 144-148. (in Chinese).
Editor: Yingzhi GUANG Proofreader: Xinxiu ZHU

- 农业生物技术(英文版)的其它文章
- Expression Analysis of Heat Shock Protein 70 Gene in Rice (Oryza sativa L.)
- Changes in Physiological and Biochemical Characteristics of Floral Organ
- Research Progress on Lonicera japonica Thunb. Affected by Environmental Stress
- Research Progress on Genetic Diversity of Snap Bean
- Allelopathic Effects of Cedrus deodara Needle Extracts on Seed
- Development Status and Countermeasures of Passiflora spp. Seedling Industry in Qinzhou, Guangxi