
非酿酒酵母产生的β-葡萄糖苷酶在发酵酒中的应用研究进展
2022-03-03 08:34覃秋杏韩小雨黄卫东战吉宬游义琳
食品科学 2022年3期
覃秋杏,韩小雨,黄卫东,战吉宬,游义琳,*
(1.中国农业大学食品科学与营养工程学院,北京市葡萄科学与酿酒技术重点实验室,北京 100083;2.中国农业大学(兴化)健康食品产业研究院,江苏 兴化 225700)
酿造过程中,为了控制不同批次酒的质量稳定,多使用单一菌种发酵剂来进行发酵,目前较为常用的酵母为酿酒酵母(Saccharomyces cerevisiae)。酿酒酵母的发酵能力极强,但这也导致发酵过程中酵母多样性下降,酵母产生的生化反应复杂程度降低,在一定程度上掩盖了不同发酵原料之间品种和地域的差异,造成发酵酒的香气和口感复杂度下降,带来了酒同质化严重、风味趋近的问题[1]。本实验室前期调研发现,随着技术的发展与进步,消费者已不再满足于传统发酵酒的单一风味,对发酵酒风味的多样性提出了更高的要求。消费者对于酒类多样化的需求和单一菌株发酵带来弊端之间的矛盾催生了关于非酿酒酵母(non-Saccharomyces cerevisiae)的研究。非酿酒酵母是发酵酒酿造中的非常规酵母(unconventional yeast),其发酵性能和逆境耐受能力都弱于酿酒酵母,但非酿酒酵母在发酵过程中可以分泌大量胞外酶,改善发酵酒的酒体、口感、色泽和泡沫等性质,其中非酿酒酵母产生的β-葡萄糖苷酶(β-glucosidase,BGL)(EC3.2.1.21)通过对糖苷结合态香气物质的水解作用可以提高发酵酒的香气复杂性。本文对发酵酒中影响β-葡萄糖苷酶活力的因素以及β-葡萄糖苷酶在不同发酵体系中的作用进行综述,为发酵酒酿造、高β-葡萄糖苷酶活力酵母筛选和相关酵母基因编辑等研究提供参考。
1 β-葡萄糖苷酶
β-葡萄糖苷酶是一类纤维素类水解酶,通过催化水解糖苷结合态物质中的糖苷键释放香气物质[2]。
1.1 β-葡萄糖苷酶来源
β-葡萄糖苷酶来源十分广泛[3],其编码基因BGL几乎存在于所有生物中[4]。食品中的β-葡萄糖苷酶主要来自于植物和微生物,植物源的β-葡萄糖苷酶主要位于浆果果皮中,与酿酒酵母产生的β-葡萄糖苷酶一样,在酒类发酵条件下(酸性(pH值3~4)、高糖含量以及一定浓度的乙醇)对于风味改善几乎没有作用[2,5-6]。因此目前研究的热点在于微生物产生的β-葡萄糖苷酶,商业使用的β-葡萄糖苷酶大多来自黑曲霉(Aspergillus niger)和哈茨木霉(Trichoderma harzianum),外源添加β-葡萄糖苷酶可以提高果汁的产量,改善成品酒的色泽、澄清度和品种香气[7-8]。真菌产生的β-葡萄糖苷酶也存在一定的不足,如外源酶制剂的纯度无法达到100%,因而可能引发安全性问题[9],β-葡萄糖苷酶对色素糖苷的水解作用可能会导致挥发性酚类物质的产生,使食品产生不良风味等[7,10]。基于以上的不足,研究者开始将目光转向其他来源的β-葡萄糖苷酶,研究发现非酿酒酵母产生的β-葡萄糖苷酶的活力较高,与霉菌产生的β-葡萄糖苷酶相比安全性高、获取方式简单且成本较低,因此非酿酒酵母产生的β-葡萄糖苷酶或许更适合用于发酵酒中[11]。
1.2 β-葡萄糖苷酶的作用机制
得益于其催化糖苷键的能力,β-葡萄糖苷酶在挥发性香气物质的产生中发挥着重要的作用。香气化合物在发酵酒中的存在方式有游离态和结合态,游离态的物质可以直接挥发产生香气,而结合态的物质需水解为游离态物质后才能释放出香气物质[12-13]。糖苷结合态香气物质由苷元和糖基组成,其中苷元是风味活性物质,水解后可增加发酵酒中植物、水果型香气[14],在酒类原料中的风味活性苷元包括中链烷醇和烯醇、莽草酸衍生物、苯衍生物、C13-降异戊二烯、单萜烯和倍半萜醇等[14-15]。
糖苷结合态香气物质的水解方式包括酸水解和酶水解。在葡萄汁中,糖苷结合态香气物质可以自发进行酸水解,释放苷元[16-17],使得香气物质游离;而酶水解过程则需要酶类的参与(图1),首先糖苷外切酶α-鼠李糖苷酶、α-阿拉伯糖苷酶、β-木糖苷酶或β-洋芹糖苷酶等作用于糖苷基末端的糖苷键,释放β-葡萄糖苷,随后β-葡萄糖苷酶作用于β-葡萄糖苷,释放苷元和葡萄糖,游离态的苷元达到感官阈值后即可对酒的香气做出贡献[16,18-19]。在糖苷结合态香气物质的水解过程中,糖苷外切酶和β-葡萄糖苷酶具有协同作用,其中β-葡萄糖苷酶是关键的限速酶[20-21],发酵过程中该酶的含量与活力影响着发酵所得成品酒的风味复杂度。
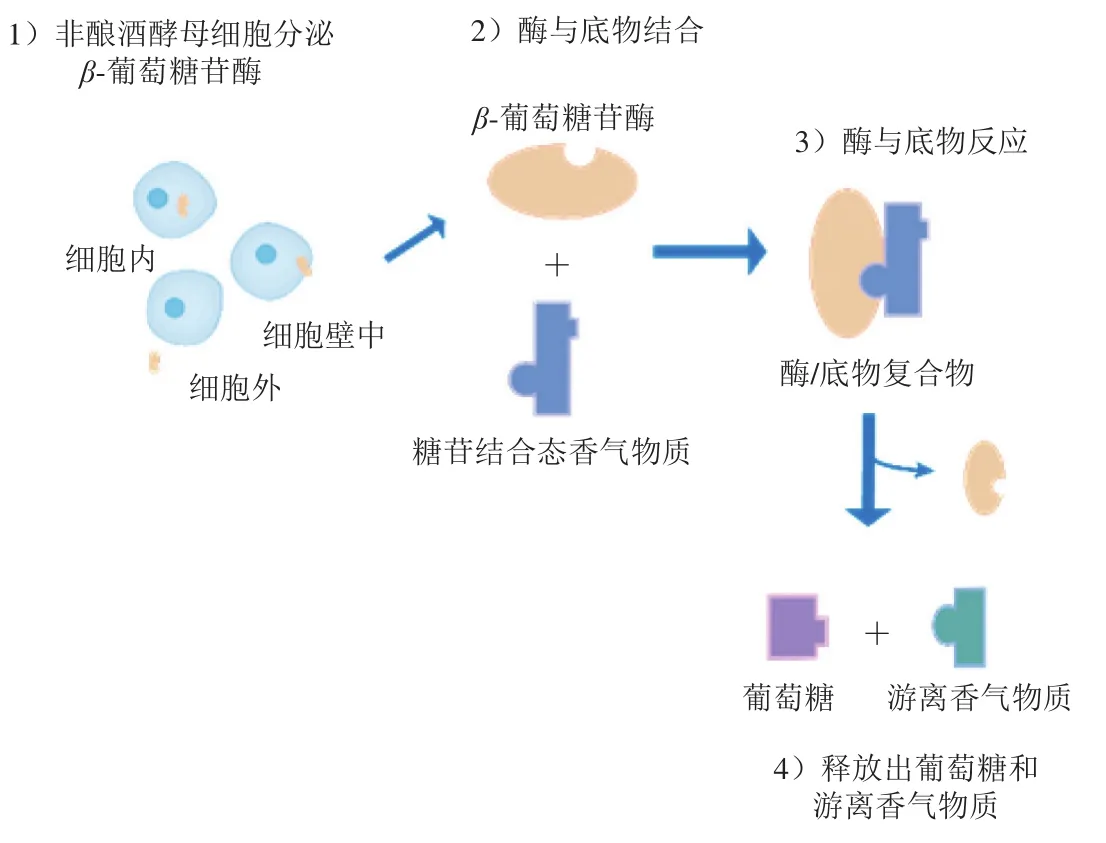
图1 β-葡萄糖苷酶作用机制示意图Fig.1 Schematic diagram of the action mechanism of β-glucosidase
不同的水解方式对酒的影响不同,酸水解导致苷元发生重新排列,反应速度慢,可能会产生不良风味[15,17];而酶水解所需的β-葡萄糖苷酶多来自原料及酵母,获取方式简单且酶水解反应速度快,并且非酿酒酵母产生的β-葡萄糖苷酶活力是所有酵母中最高的,可以在酿酒酵母无法产生β-葡萄糖苷酶或酶活力不足的情况下进行补充,从而提高发酵酒香气的复杂度[22]。
2 β-葡萄糖苷酶活力的影响因素
与其他种类的酶一样,β-葡萄糖苷酶活力也受到多种因素的影响,酵母合成和释放β-葡萄糖苷酶的能力、发酵环境中的温度、酸碱度和可发酵糖浓度都会对其酶活力产生影响[23]。
2.1 酵母合成和释放β-葡萄糖苷酶的能力对β-葡萄糖苷酶活力的影响
大部分非酿酒酵母都可以合成和释放β-葡萄糖苷酶,然而非酿酒酵母具有非常强的菌株特异性,导致菌株产生的β-葡萄糖苷酶活力存在着种间差异和种内差异,不同酵母细胞中β-葡萄糖苷酶的存在位置和聚合方式也有所不同。王佳等[24]发现不同的非酿酒酵母产生β-葡萄糖苷酶的能力不同,其中陆生伊萨酵母(Issatchenkia terricola)和季也蒙毕赤酵母(Pichia guilliermondii)产酶能力最强。对非酿酒酵母产生的β-葡萄糖苷酶进行酶活定位时发现,β-葡萄糖苷酶在酵母细胞中有细胞外、细胞内和细胞壁中3 种存在方式。在对123 株季也蒙有孢汉逊酵母(Hanseniaspora guilliermondii)中β-葡萄糖苷酶活力进行测定时,发现细胞外β-葡萄糖苷酶活力最高,73%的菌株酶活力在80~120 pNP nmol/(mL·h)(pNP表示4-硝基苯酚),而在70 株葡萄汁有孢汉逊酵母(Hanseniaspora uvarum)中,细胞外、细胞内和细胞壁中的β-葡萄糖苷酶活力为30~40 pNP nmol/(mL·h)的菌株所占比例分别为37%、30%和29%,无论是同一菌属或是不同菌属,酶活定位的不同导致酶活力存在差异[13]。李爱华等[25]发现葡萄汁有孢汉逊酵母产生的β-葡萄糖苷酶为胞外酶,超声破碎后细胞上清液中的β-葡萄糖苷酶活力远高于其他样品,但也有研究发现胞内β-葡萄糖苷酶活力高于胞壁结合酶活力,因此细胞中酶活定位的不同会带来酶活力的不同,但是无法根据酶活定位来确定酶活力高低[26]。酵母细胞中β-葡萄糖苷酶有多种聚合方式,包括单聚体、二聚体、三聚体和四聚体[27],已有研究发现在不同菌属的细胞或同一菌属不同菌株的细胞中,都会存在β-葡萄糖苷酶聚合方式的差异,β-葡萄糖苷酶活力也存在不同[28-29]。酵母合成和释放β-葡萄糖苷酶的能力不同导致β-葡萄糖苷酶的活力不同再次验证了菌株特异性对于非酿酒酵母在啤酒中应用效果的影响,也提示着发酵前菌株筛选的必要性。
酿造过程中酿酒酵母和非酿酒酵母之间会产生相互作用,导致菌株β-葡萄糖苷酶活力发生改变[30]。非酿酒酵母的发酵能力较弱,为了获得较高的发酵度,通常会在接种非酿酒酵母的发酵液中同时或顺序接种酿酒酵母[31]。在美极梅奇酵母(Metschnikowia pulcherrima)和酿酒酵母共发酵葡萄酒的过程中,研究人员发现无论是混合接种还是顺序接种,共发酵组中的β-葡萄糖苷酶水解产生的橙花醇和香叶醇的含量都会显著低于美极梅奇酵母单菌发酵组[32],徐建坤[33]也得到了类似结果,虽然混合发酵组酶活力均高于酿酒酵母单菌发酵组,但混合接种时发酵液中最高β-葡萄糖苷酶活力远低于非酿酒酵母在最适条件下的酶活力,酿酒酵母与非酿酒酵母共发酵时产生的相互作用会影响β-葡萄糖苷酶的活力。
2.2 发酵环境对β-葡萄糖苷酶活力的影响
β-葡萄糖苷酶对环境的变化十分敏感,且不同的环境对酶活力的影响不同。
酒类的发酵温度通常低于25 ℃,而β-葡萄糖糖苷酶的最适温度为40~50 ℃,远高于发酵温度,导致β-葡萄糖苷酶在发酵温度时的活力低于最适温度时的活力[34]。β-葡萄糖苷酶对高温和低温的耐受性也并不一致,其对于高温的耐受能力弱于低温。本课题组前期研究发现4 株野生非酿酒酵母的最适温度为50 ℃,温度升高到70 ℃时β-葡萄糖苷酶的相对酶活力降低为44%,而当温度降低为10 ℃时相对酶活力大于54%,结果显示β-葡萄糖苷酶对低温的耐受性更好[16]。β-葡萄糖苷酶较好的低温耐受力可以使其在酒类的发酵温度下维持较高的活力,获得较好的发酵效果。
环境酸碱度的改变通过影响β-葡萄糖苷酶结构从而改变其酶活力。酸碱度改变时,酶的二级结构发生改变。β-葡萄糖苷酶的最适pH值为5.0~6.0,在最适pH值条件下,β-葡萄糖苷酶α-螺旋和β-折叠相对含量最高,无规卷曲相对含量最低(BGL1:26.7%,BGL2:23.8%,AS:24.2%),pH值降低,α-螺旋和β-折叠的相对含量减少,β-转角和无规卷曲相对含量增加,酶活力降低[35]。对巴斯德毕赤酵母(Pichia pastoris)Y8产生的多种酶的pH值耐受性进行检测,发现在pH 5.0时β-葡萄糖苷酶活力最高,而当pH值为2.0时,酶活力会降低至pH 5.0时的19%,并且β-葡萄糖苷酶对于酸性环境的敏感性远高于其他酶类[2]。在常见的发酵酒体系中,常规啤酒的pH值在4.0~5.0之间[36-37](酸啤酒由于含有乳酸,pH值可降至3.3[38]),葡萄汁和果汁由于含有较多的果酸,pH值一般为2.0~4.0[39-40],β-葡萄糖苷酶的高酸敏感性可能会导致其在一些pH值较低的果酒(如葡萄酒)中的作用效果较差。
发酵液中还含有大量的可发酵糖(葡萄糖、果糖和蔗糖等),其中葡萄糖对于β-葡萄糖苷酶活力有较大的影响[41]。Lopez等[18]对4 种非酿酒酵母产生的β-葡萄糖苷酶的糖耐受能力进行了研究,发现实验菌株的β-葡萄糖苷酶活力随着葡萄糖浓度升高而逐渐下降,当葡萄糖浓度达到200 mmol/L后活力仅为未添加葡萄糖时的20%,而果糖和蔗糖对酶活力几乎没有影响。葡萄糖是非酿酒酵母酒精发酵过程中不可缺少的重要糖类,但其对于β-葡萄糖苷酶活力的影响可能会导致成品酒的风味物质含量发生改变。
乙醇是酵母酒精发酵的产物,发酵过程中可发酵糖的消耗可以缓解糖类对β-葡萄糖苷酶活力的抑制作用,但乙醇含量的增加会抑制酶活力,并且乙醇对β-葡萄糖苷酶的抑制作用随乙醇浓度的增大而增强。果酒和啤酒的酒精度较低,乙醇体积分数一般为2%~5%,此条件下对β-葡萄糖苷酶活力的抑制作用较弱[42-43],而葡萄酒的乙醇体积分数一般大于12%,在此条件下β-葡萄糖苷酶的活力受到强烈抑制[11,44]。乙醇对β-葡萄糖苷酶活力的抑制作用随浓度增加而增强的情况或许提示高β-葡萄糖苷酶活力的非酿酒酵母对于低酒精度酒的增香更为有效。
β-葡萄糖苷酶活力的影响因素众多,除上述因素外,发酵时间[17]、金属离子[16]等也会导致酶活力发生改变,因此在筛选酵母或对酵母进行改造时,除了对出发菌株自身特性进行考察,还需要考虑所得实验菌株产生的β-葡萄糖苷酶对发酵环境的适应能力。表1对部分非酿酒酵母产生的β-葡萄糖苷酶的性质进行了总结。
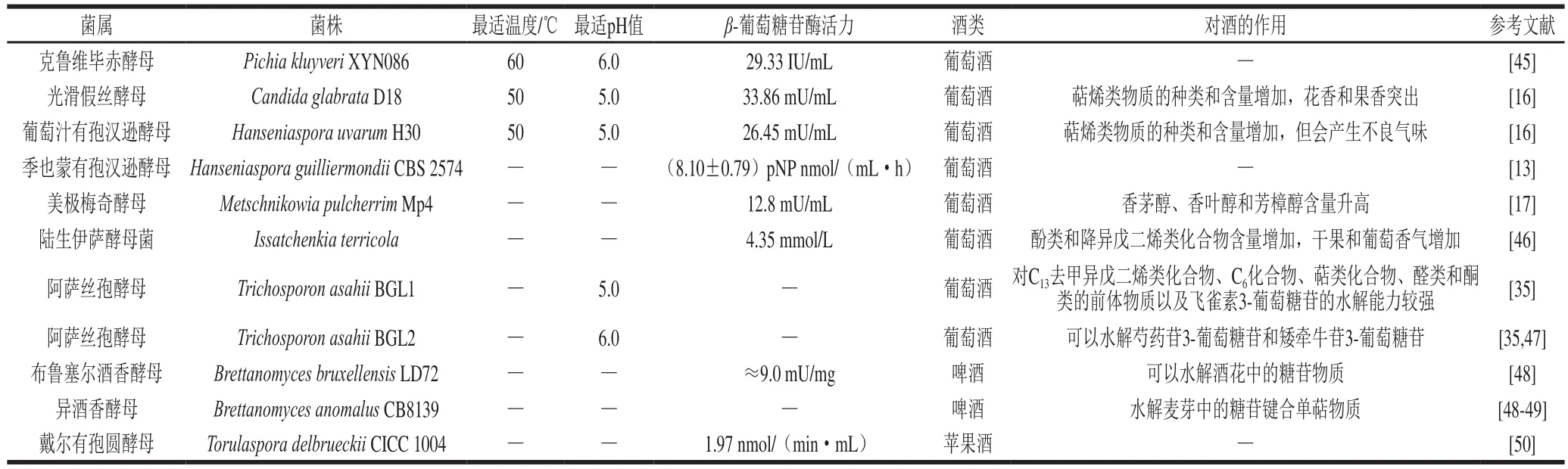
表1 部分非酿酒酵母来源的β-葡萄糖苷酶的性质Table 1 Properties of β-glucosidases from non-Saccharomyces yeasts
3 β-葡萄糖苷酶在发酵酒中的应用
β-葡萄糖苷酶广泛存在于非酿酒酵母中,其在假丝酵母属(Candidasp.)、耐热克鲁维酵母(Lachancea thermotolerans)、毕赤氏酵母(Pichiasp.)、有孢汉逊酵母(Hanseniasporasp.)、梅氏酵母(Metschnikowia zizyphicola)、东方伊萨酵母(Issatchenkia orientalis)和酒香酵母(Brettanomycessp.)等菌属的某些菌株中具有较高的活力[45,51-53],可用于发酵酒以提高香气复杂度,并对酒体的颜色稳定性产生影响。
3.1 β-葡萄糖苷酶在葡萄酒中的应用
β-葡萄糖苷酶在葡萄酒中的作用不仅包括水解糖苷键增加香气复杂度,部分非酿酒酵母产生的β-葡萄糖苷酶对花色苷的水解作用也会对葡萄酒的颜色产生影响。研究发现高β-葡萄糖苷酶活力的非酿酒酵母可以将葡萄果实中的糖苷结合态香气物质如单萜烯、C13-降异戊二烯和芳香族化合物等释放出来。在本课题组前期研究中,将葡萄汁有孢汉逊酵母H30(Hanseniaspora uvarumH30)和光滑假丝酵母D18(Candida glabrataD18)与酿酒酵母顺序接种于葡萄汁中进行发酵,发现这两株非酿酒酵母都可以增加成品酒中的萜烯类物质和C13-降戊二烯类物质的含量,并且光滑假丝酵母D18可以使发酵酒中品种香种类增多[16]。高β-葡萄糖苷酶活力的季也蒙有孢汉逊酵母(Hanseniaspora guilliermondii)BF1被用于无味菲亚诺葡萄(Fiano)的发酵中,无论是BF1单菌发酵还是与酿酒酵母混菌发酵,成品酒中芳樟醇和松油烯-4-醇的含量均较高,感官评价显示两款酒中水果味和玫瑰花香最为突出[13]。β-葡萄糖苷酶还与葡萄酒中萜烯类物质的产生有关,王兴凯[17]发现葡萄模拟汁中萜烯类物质(主要是芳樟醇、香茅醇和香叶醇)的变化与β-葡萄糖苷酶活力的变化趋势一致,而从陆生伊萨酵母菌(Issatchenkia terricola)中分离得到的β-葡萄糖苷酶用于葡萄酒酿造时,则可以使葡萄酒中酚类物质含量增加83%,降异戊二烯化合物含量增加65%[46]。多种高β-葡萄糖苷酶活力的非酿酒酵母已被证明可以用于葡萄酒的发酵,且在增加香气物质特别是萜烯类物质和C13-降戊二烯类物质等含量方面有重要作用。
除了香气特征外,颜色也是葡萄酒重要的属性之一,花青素是葡萄酒中重要的色素化合物,大部分的花青素是单葡萄糖苷,部分β-葡萄糖苷酶可使糖苷键断裂,游离的花青素迅速降解成无色化合物,导致葡萄酒色泽不佳甚至失色[54-56]。不同菌株产生的β-葡萄糖苷酶对花色苷的作用不同,研究者将阿萨丝孢酵母(Trichosporon asahii)所产的β-葡萄糖苷酶加入至蛇龙珠葡萄汁中进行发酵,发现β-葡萄糖苷酶对花色苷特别是飞燕草素葡萄糖苷具有较强的降解作用[35],而在Vernocchi等[57]的研究中,将含有高β-葡萄糖苷酶活力的酿酒酵母BV12和BV14接种于桑娇维塞(Sangiovese)葡萄汁时,并未发现这两株酵母对葡萄酒颜色的不良影响。β-葡萄糖苷酶对于花色苷的分解作用的差异可能与底物的结构特征和β-葡萄糖苷酶的性质有关,β-葡萄糖苷酶对花色苷的分解作用也可以作为一种改变葡萄酒颜色的方法加以利用[58]。
3.2 β-葡萄糖苷酶在啤酒中的应用
啤酒中的糖苷结合态香气物质主要来自啤酒花,酒花中含有的糖苷结合态香气物质包括脂肪醇、萜烯醇和C13-降异戊二烯类化合物等[14,59],这些物质在发酵过程中被水解释放,从而改善啤酒的香气复杂性[60]。其中,萜烯类糖苷物质的香气阈值较低,水解时产生的萜烯类物质(如月桂烯、香叶醇、佛手柑醇和芳樟醇等)具有浓郁的花香和水果香气[61],单萜烯还可以进一步转化为β-香茅醇或α-萜烯醇,增强花香味和柠檬味[62]。C13-降异戊二烯包括β-大马士酮和β-紫罗兰酮等,是β-葡萄糖苷酶水解产生的重要风味物质,可以产生令人愉悦的香气[63]。
目前在啤酒发酵体系中利用非酿酒酵母产生β-葡萄糖苷酶的性质进行增香的研究较少,以酒香酵母的研究最为透彻。酒香酵母因其可以产生大量的酚类物质从而导致异味,在过去通常被认为是导致腐败的酵母[64],但随着对酒香酵母研究的不断深入,酒香酵母可以产生β-葡萄糖苷酶的特性逐渐被开发,并且发现部分菌株在发酵过程中并未产生不良风味,这也加速了酒香酵母在啤酒中的应用研究[65-66]。布鲁塞尔酒香酵母(Brettanomyces bruxellensis)和异酒香酵母(Brettanomyces anomalus)是啤酒发酵中较为常见的两类酒香酵母,常用于贵兹啤酒和兰比克啤酒的发酵中。酒香酵母产生的β-葡萄糖苷酶可以作用于酒花和麦芽,当酒香酵母作用于酒花时,可以水解释放出酒花中含有的苷元,如芳樟醇、水杨酸甲酯、顺-3-己烯-1-醇和1-辛烯-3-醇等,且水解所得苷元物质的质量浓度大于0.005 mg/L[48]。当酒香酵母作用于麦芽时,可以使得麦芽中的糖苷键断裂,使麦芽中的萜烯类物质游离出来[49]。酒香酵母能否产生β-葡萄糖苷酶与其是否含有BGL1和BGL2基因有关,同时含有这两个基因,或者仅含BGL2基因的酒香酵母具有较高的胞外β-葡萄糖苷酶活力,并且在发酵过程中酶活力高于仅含BGL1或不含BGL1和BGL2基因的酵母[67]。部分酿酒酵母中编码外切葡聚糖酶的EXG基因也被证实具有非特异作用于葡萄糖聚合物和较小的糖基结合底物的活力[68],高EXG活力的酵母菌株在发酵过程中对酒花糖苷也有一定的分解能力,但不同菌株之间的活力相差较大[69],且其对于酒花糖苷的分解能力远低于非酿酒酵母[48],基因组成决定了酿酒酵母和非酿酒酵母是否具有产生高活力β-葡萄糖苷酶的能力。酒香酵母目前多用于特种啤酒的发酵,但随着对其基因组学和代谢组学研究的不断深入,其在啤酒发酵中的应用也会大大扩宽。
耐热克鲁维酵母具有较高β-葡萄糖苷酶活力的特性也被应用于啤酒发酵中,可以促进酒花中糖苷前体物质的释放[70-71],耐热克鲁维酵母还是一种潜在的啤酒发酵剂,在啤酒发酵中可产生乳酸降低pH值[72],与酸啤酒发酵常用的乳酸菌相比,耐热克鲁维酵母耐受异α-酸的能力更高,更适合用于酸啤酒的发酵。耐热克鲁维酵母在发酵过程中可以产生甘油,对于改善酒体和口感有重要作用[71]。耐热克鲁维酵母在产β-葡萄糖苷酶、发酵能力、产乳酸、产甘油等方面优于其他非酿酒酵母,因此其在啤酒中的应用前景也更广阔。
高β-葡萄糖苷酶活力的非酿酒酵母种类众多,并且大部分都在葡萄酒增香中有较好表现,但其在啤酒体系中的研究较少,布雷特酵母和耐热克鲁维酵母在啤酒增香中的作用表明高β-葡萄糖苷酶活力的非酿酒酵母对于啤酒风味具有改善作用,这也侧面说明其他高β-葡萄糖苷酶活力的酵母在啤酒发酵中也具有广阔的应用前景。
3.3 β-葡萄糖苷酶在其他发酵酒中的作用
β-葡萄糖苷酶在发酵酒中的应用并不局限于葡萄酒和啤酒,在一些果酒如苹果酒[50]、毛桃果酒[73]等发酵体系中也有相关的研究,β-葡萄糖苷酶在这些酒中的作用不仅包括增香,还可以改善酒的品质。在苹果汁中单独或混合接种戴尔有孢圆酵母(Torulaspora delbrueckii)与酿酒酵母,发现混合发酵时酵母产生的β-葡萄糖苷酶、α-淀粉酶和果胶酶的活力均高于单菌发酵时的活力,并且酶活力越高对苹果酒香气的改善效果越好[50]。在毛桃果酒酿造过程中由于果实颜色不一,产品会发生变色和浑浊,在发酵过程中加入β-葡萄糖苷酶后,花色素苷发生水解,毛桃果酒颜色稳定性提高[73]。在果酒发酵中使用非酿酒酵母,可以筛选和驯化适用于果酒的非酿酒酵母,随着研究的深入,非酿酒酵母的性能开发也将更为多样。
3.4 不同发酵体系对非酿酒酵母要求的比较
β-葡萄糖苷酶对于香气的改善作用在葡萄酒、啤酒、苹果酒等体系中都得到了验证,并且不同菌株产生的β-葡萄糖苷酶活力不同,不同的发酵汁对β-葡萄糖苷酶活力的影响也不同。葡萄酒等果酒发酵汁中,含有较多的酸性物质,pH值较低,并且葡萄酒的酒精度通常较高,因此筛选用于果酒发酵的非酿酒酵母时,需要对其酒精耐受力和耐酸能力进行测试。啤酒发酵汁与以上酒类的不同之处在于其在发酵过程中加入了啤酒花,啤酒花在煮沸过程会产生异α-酸,异α-酸在酸性环境中可以抑制酵母的生长[74],导致非酿酒酵母的延滞期延长[75],因此用于啤酒增香发酵的非酿酒酵母除了高产β-葡萄糖苷酶外还需要具有一定的耐受异α-酸等的能力。不同发酵体系中酵母的适应性不同,因此在发酵液中接种非酿酒酵母时需先根据发酵体系中对菌株进行筛选。
3.5 获取具有β-葡萄糖苷酶活力菌株的方法
目前获得高产β-葡萄糖苷酶的酵母的方法有很多种,包括直接筛选、诱变育种和基因工程等(表2)。直接筛选法指从含有β-葡萄糖苷酶的基质中,通过一定的方法筛选出可以产生β-葡萄糖苷酶的酵母,牟含[76]通过七叶苷显色法从白酒酒窖中筛选得到了多株高产β-葡萄糖苷酶的非酿酒酵母,并将所得非酿酒酵母用于葡萄酒增香酿造中。产β-葡萄糖苷酶的菌株来源十分广泛,除了酒类发酵醪,各种细菌和真菌[77]以及茶树类植物[78]也是其重要来源,因此通过直接筛选法获得高产β-葡萄糖苷酶的菌株是一个极其普适的方法。诱变育种指利用物理或化学的方法使酵母的遗传物质和性能发生改变的一种方法,研究人员通过常压室温等离子体与紫外复合诱变技术对两株酿酒酵母进复合诱变,所得菌株产生的β-葡萄糖苷酶活力远高于出发菌株[79]。诱变育种方法具有快速、突变率高等特点,可以在短时间内形成大量突变,因此也是获得高β-葡萄糖苷酶活力菌株的较好方式[80]。基因工程技术是指直接在出发菌株中导入目的基因从而获得高产所需物质菌株的方法,目前较为常用的技术为CRISPR/Cas9技术,研究人员利用该技术将酵母细胞壁蛋白Cwp2p编码基因破坏后,使得酵母分泌的β-葡萄糖苷酶活力提高[81]。通过基因工程技术寻找与β-葡萄糖苷酶相关的靶点基因,可以达到提高出发菌株酶活力的目的。以上方法都是目前较为常用的获取新性能酵母的方法,但是也不可避免地存在一定的不足,这也导致在进行高酶活力酵母的筛选和改造时,需要根据所需特性及不同方法的优缺点进行选择。

表2 获取产β-葡萄糖苷酶的酵母菌株方法比较Table 2 Comparison of methods for obtaining yeast strains with β-glucosidase
4 结 语
高β-葡萄糖苷酶活力的非酿酒酵母在发酵酒中的应用已经在葡萄酒、啤酒和其他发酵酒等多个发酵体系中得到验证(图2)。在我国传统的酒精饮料白酒的酒曲和黄水以及黄酒的酒曲中,也有关于高产β-葡萄糖苷酶菌株的发现,研究人员将其用于葡萄酒的发酵中可以增加酒的柑橘类、甜果类、香料类香气[86-89]。大量研究表明非酿酒酵母产生的β-葡萄糖苷酶对于发酵酒的作用包括水解糖苷结合态香气物质,释放C13-降异戊二烯、单萜烯、倍半萜醇和脂肪醇等增加酒类香气的复杂度。β-葡萄糖苷酶对花色苷的水解作用也影响着酒类的颜色稳定性。部分高β-葡萄糖苷酶活力的非酿酒酵母在发酵中还可以产生乳酸和甘油,改善酒的口感和风味。

图2 β-葡萄糖苷酶活力的影响因素及其在不同发酵体系中的应用Fig 2 Factors affecting β-glucosidase activity and its application in different fermentation systems
虽然高产β-葡萄糖苷酶的非酿酒酵母在发酵酒中的应用研究已经取得一定的进展,但由于菌株特异性以及β-葡萄糖苷酶活力的影响因素众多等原因,导致其在实际应用中仍存在一些问题。商业酿酒酵母是一种常用的发酵剂,在多种发酵环境中都具有较好的发酵能力,而非酿酒酵母并非普适性的发酵剂,其在不同发酵体系和发酵条件的适用性还有待验证。对于大部分非酿酒酵母而言,由于其本身对于发酵环境更为敏感,并且发酵能力较弱,因此在发酵体系和发酵条件发生改变时,适用于新环境的非酿酒酵母需要进行改造或重新筛选。非酿酒酵母虽然已经被广泛研究,但其并不是被长期使用的公认安全的酵母,由于其会产生异味等性质,在过去甚至被认为是腐败酵母[42],部分非酿酒酵母本身是一种病原体,在发酵中可能会导致食品安全问题[49],因此非酿酒酵母本身以及产生的物质的生物安全性还需要进一步验证。目前除了自然发酵酒外,大部分非酿酒酵母在发酵酒中的应用处于实验室阶段,而高β-葡萄糖苷酶活力的非酿酒酵母在发酵酒中的应用研究才刚刚起步,实现产业化还有漫长的发展历程。
微生物产生的β-葡萄糖苷酶不仅可以应用于酒类的增香,在速溶茶增香[21]、食品脱苦[90]、白藜芦醇生成[29]、异黄酮糖苷转化为异黄酮苷元[91]、土壤改善[92]、病虫害防治[93]等方面都有着良好的应用前景,无论是通过从自然基质中筛选获得含有β-葡萄糖苷酶的菌株,或是通过诱变育种和基因工程的方法改造从而获得高酶活力菌株,随着代谢组学和全基因组测序研究的不断深入,将β-葡萄糖苷酶用于工业生产是其未来发展的重要方向。
猜你喜欢
中草药(2022年20期)2022-11-15
现代食品科技(2022年9期)2022-10-09
酿酒科技(2021年8期)2021-12-06
军事文摘·科学少年(2021年1期)2021-02-04
化工设计通讯(2021年2期)2021-01-07
酿酒科技(2020年7期)2020-12-19
中国畜牧业(2019年22期)2019-12-30
名人传记·财富人物(2017年9期)2017-11-02
名人传记·财富人物(2017年9期)2017-11-02
故事作文·低年级(2016年7期)2016-05-14
