
IL⁃17 诱导的p300 调控NSCLC 细胞迁移、侵袭和MMP2 表达的作用
2022-03-02 04:57张志伟蒋馨怡何庆玲阮玉婷吴宁霞王伟民王迎伟赵晨卉
南京医科大学学报(自然科学版) 2022年2期
李 雅,葛 文,张志伟,蒋馨怡,何庆玲,阮玉婷,吴宁霞,应 帅,王伟民,张 婧,邱 文,王迎伟*,赵晨卉
1南京医科大学免疫学系,江苏 南京 211166;2南京医科大学第一附属医院肿瘤科,江苏 南京 210029
肺癌是人类常见的恶性肿瘤,病理上分为小细胞肺癌和非小细胞肺癌(non⁃small cell lung cancer,NSCLC)两大类,后者约占临床病例的85%[1-2]。由于NSCLC发病率高,患者死亡又多与瘤细胞的侵袭转移有关,故研究NSCLC的转移机制极为重要。
NSCLC 发生发展与多种因素密切相关。近年来,关于肺部慢性炎症诱发NSCLC的文献日趋增多,其机制不仅涉及肿瘤局部促炎因子的表达上调[3],而且某些因子还能诱导瘤细胞的侵袭和转移[4-5]。已知IL⁃17 是一种典型的促炎因子,其在多种肿瘤组织中的表达常显著上调[4-5]。本课题组以往的研究表明,在NSCLC 的癌组织内IL⁃17 及IL⁃17 受体(IL⁃17RA)的表达明显升高。体外用IL⁃17 刺激NSCLC细胞后也能诱导其增生、迁移与侵袭[6]。不过,其诱导细胞迁移与侵袭的分子机制并不清楚。
基质金属蛋白酶(matrix metalloproteinase,MMP)是Zn2+依赖的蛋白水解酶,可分为6大类28个成员,多由成纤维细胞和肿瘤细胞等合成,通过降解细胞外基质而促进肿瘤转移[4-5]。有资料显示,MMP2广泛表达,在乳腺癌转移中起增强效应[4-5]。本课题前期体外实验已发现,受IL⁃17刺激的NSCLC细胞中MMP2的表达显著增加。
已知腺病毒E1A 相关300 kDa 蛋白(adenoviral E1A binding protein of 300 kDa,p300)是一种转录辅激活因子[7]。研究证实,p300能促进NSCLC的迁移与侵袭,其机制与p300辅助修饰某些转录因子促进了靶基因的表达有关[7]。本课题前期研究表明,用IL⁃17刺激NSCLC细胞后,其胞内p300和MMP2的表达明显上调。因此,IL⁃17上调的p300是否能通过调控MMP基因表达增强NSCLC细胞的侵袭和转移,值得进一步探究。
1 材料和方法
1.1 材料
人正常支气管上皮细胞系BEAS⁃2B 和NSCLC细胞系(A549、PC9、H1299)由武汉大学中国典型培养物保藏中心提供。
胎牛血清(南京诺唯赞生物公司);Lipofectamine 2000(赛默飞世尔科技公司,美国);兔p300 多克隆抗体(CST 公司,美国);兔MMP2 多克隆抗体(杭州华安生物公司);小鼠β⁃actin 单克隆抗体(上海碧云天生物公司);羊抗兔、鼠IgG⁃HRP 二抗(合肥Biosharp 公司)。siIL⁃17RA 购买于上海吉玛基因公司。
1.2 方法
1.2.1 BEAS⁃2B、A549、PC9和H1299细胞系的培养及IL⁃17RA的检测
将细胞接种于DMEM 完全培养液(10%胎牛血清)中,置于37 ℃、5%CO2孵箱中培养48 h。当细胞融合度达到90%时,按1∶5比例进行传代培养。
Western blot 实验:细胞裂解物离心3 min 取30 μg 蛋白行SDS⁃PAGE 电泳。电泳2 h 再湿转120 min。待蛋白转印到PVDF 后用脱脂牛奶封闭2 h,加IL⁃17RA一抗4 ℃过夜。用HRP 标记的二抗孵育1 h。最后行ECL化学发光试剂测定。
1.2.2 沉默IL⁃17RA基因后检测H1299细胞迁移和侵袭能力
培养H1299 细胞,待其生长融合度达80%时进行siIL⁃17RA质粒的转染。先用DMEM对4 μL lipo⁃fectamine 2000和2000 ng siIL⁃17RA 进行稀释,然后将二者混合后静置15 min 再加细胞,置37 ℃、5%CO2条件下继续培养。
细胞划痕实验:将转染siIL⁃17RA质粒的H1299细胞饥饿12 h 再行培养。当融合度达90%时,用200 μL 移液器吸头产生细胞单层的线性伤口,PBS清洗后加含1%FBS 的DMEM,同时加50 ng/mL 的IL⁃17。继续培养并在0 h和24 h观察其生长情况。
Transwell实验:将转染siIL⁃17RA的H1299细胞饥饿12 h 再接种至含1%FBS DMEM 的Transwell 小室中(5×104个/孔)。另下室加600 μL 相同的培养液。培养24 h 去除上清,并洗去未穿透膜的细胞。迁移到膜下的细胞行100%甲醇固定和0.1%结晶紫染色。显微镜下观察后拍摄5个随机选择的视野。
1.2.3 IL⁃17 刺激H1299 细胞后检测不同时间p300和MMP2的表达
收集IL⁃17 刺激后0、1、2、3、6、12 h 的H1299 细胞蛋白,行Western blot 检测p300 和MMP2 的表达,方法同前。
1.2.4 沉默IL⁃17RA基因后p300和MMP2蛋白表达的测定
将siIL⁃17RA质粒转染H1299细胞后48 h,再用IL⁃17 刺激3 h。用Western blot 检测p300 和MMP2的表达,实验分组及检查方法同前所述。
1.2.5 过表达或沉默p300 对细胞迁移、侵袭及MMP2表达的影响
1.2.5.1 p300 过表达和shp300 小干扰质粒的来源及验证
将p300 过表达质粒用Lipofectamine 2000 转染H1299 细胞48 h,实验分组为:①pcDNA3.1 组;②pcDNA3.1/p300 组。收集细胞蛋白行Western blot检查。
同样,将shp300 小干扰质粒转染细胞48 h,之后用IL⁃17 刺激3 h。实验分组为:①shCTR+IL⁃17组;②shp300⁃1+IL⁃17 组;③shp300⁃2+IL⁃17 组;④shp300⁃3+IL⁃17组。收集细胞蛋白行Westernblot检查。
1.2.5.2 过表达或沉默p300 对H1299 细胞迁移和侵袭的影响
实验分为:①pcDNA3.1 组;②pcDNA3.1/p300组;③shCTR+IL⁃17组;④shp300+IL⁃17组。检查时间点为0 h和24 h。具体方法同前。
1.2.5.3 过表达或沉默p300 影响H1299 细胞表达MMP2的测定
过表达p300 的分组为:①pcDNA3.1 组;②pcD⁃NA3.1/p300 组。沉默p300 的分组为:①shCTR 组;②shCTR+IL⁃17 组;③shp300+IL⁃17 组。用Western blot检查MMP2的表达,时间点选择见前所述。
1.3 统计学方法
所有实验均独立重复3次,数据以均数±标准差()表示。采用SPSS 19.0 统计软件进行统计学分析,多组间比较则采用单因素方差分析,Bonfferoni法进行两两比较,P<0.05为差异有统计学意义。
2 结果
2.1 NSCLC细胞系IL⁃17RA的表达
为了证实NSCLC 细胞表面存在IL⁃17RA,检测了正常支气管上皮细胞BEAS⁃2B 及NSCLC 的3 种细胞系(A549、PC9和H1299)。结果显示,这些细胞均有IL⁃17RA 的表达,但NSCLC 细胞系表达更为明显,尤以H1299 细胞显著(图1),故后续选择H1299细胞系开展实验。
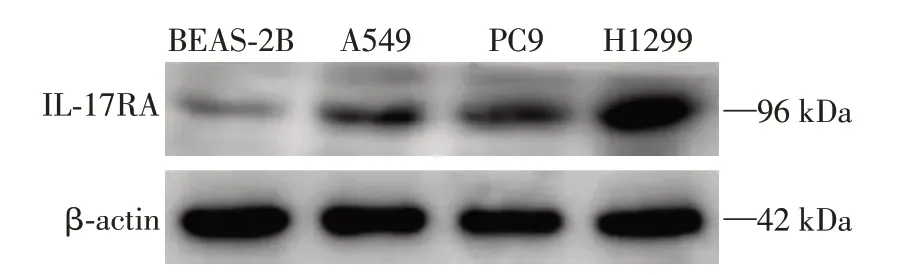
图1 IL⁃17RA在支气管上皮细胞(BEAS⁃2B)和NSCLC 细胞系(A549、PC9和H1299)中的表达Figure 1 The expression of IL⁃17RA in bronchi epithelial cell(BEAS⁃2B)and NSCLC cell line(A549,PC9 and H1299)
2.2 敲低IL⁃17RA 对IL⁃17 诱导的H1299 细胞迁移和侵袭的影响
为了确定IL⁃17 可通过结合IL⁃17RA 来影响H1299 细胞的迁移和侵袭,在转染了siIL⁃17RA 的H1299细胞中进行划痕和Transwell实验。结果发现,转染siIL⁃17RA质粒48 h后再用IL⁃17(50 ng/mL)刺激24 h,siIL⁃17RA+IL⁃17组的细胞迁移和侵袭能力均明显降低(图2)。

图2 敲低IL⁃17RA表达对IL⁃17诱导细胞迁移和侵袭的影响Figure 2 Influence of IL⁃17RA knockdown on IL⁃17⁃induced cell migration and invasion
2.3 IL⁃17 刺激H1299 细胞不同时间p300 和MMP2蛋白表达的变化
用前述剂量的IL⁃17刺激H1299细胞0、1、2、3、6、12 h,采用Western blot 检测p300和MMP2蛋白表达。IL⁃17 刺激后2 h,p300 和MMP2 的表达显著升高,3 h 时达到高峰(图3)。因此,后续选择了IL⁃17刺激细胞3 h作为蛋白检测的时间点。

图3 Western blot 检测IL⁃17 刺激H1299 细胞不同时间p300和MMP2的表达Figure 3 Expression of p300 and MMP2 at different time in H1299 cell stimulated with IL⁃17 by Western blot
2.4 敲低IL⁃17RA 表达对IL⁃17 诱导的H1299 细胞p300和MMP2表达的影响
为了明确p300 和MMP2 蛋白的表达确为IL⁃17刺激及IL⁃17结合IL⁃17RA所致,将H1299细胞转染siIL⁃17RA 48 h 再用IL⁃17 刺激3 h。结果显示,IL⁃17刺激细胞后3 h能显著上调p300和MMP2的蛋白表达,而siIL⁃17RA+IL⁃17组p300和MMP2的蛋白水平则显著减少(图4)。

图4 Western blot 检测敲低IL⁃17RA 影响p300 和MMP2的表达Figure 4 IL⁃17RA knockdown affects p300 and MMP2 expression by Western blot
2.5 过表达或敲低p300 对H1299 细胞迁移、侵袭及表达MMP2的影响
2.5.1 p300 过表达和shp300 质粒验证及shp300 有效沉默靶点的确定
将pcDNA3.1/p300过表达质粒和对照质粒分别转染H1299 细胞48 h(转染效率约70%),收集细胞蛋白检测p300 的表达。结果显示,转染pcDNA3.1/p300的细胞中,p300的表达显著增加(图5)。
将构建的3 个针对不同靶点的shRNAp300⁃1、shRNAp300⁃2、shRNAp300⁃3 质粒和其对照shCTR质粒分别转染H1299 细胞48 h 发现,shp300⁃2 能显著下调p300 蛋白的表达(图5),故后续选取了shp300⁃2进行实验,并统一简化命名为shp300。

图5 Western blot检测pcDNA3.1/p300和shp300质粒表达的验证Figure 5 Expression verification of pcDNA3.1/p300 and shp300 plasmids by Western blot
2.5.2 过表达或敲低p300 对H1299 细胞迁移和侵袭的影响
为了检查p300 基因对H1299 细胞迁移和侵袭的影响,在细胞转染p300 表达质粒和shp300 质粒后进行了划痕和侵袭实验。结果表明,转染pcDNA3.1/p300 质粒可增加细胞的迁移和侵袭能力,而转染shp300质粒48 h后再行IL⁃17刺激24 h,其迁移和侵袭能力均明显降低(图6)。

图6 过表达和敲低p300影响细胞的迁移与侵袭Figure 6 Effects of p300 overexpression and knockdown on cell migration and invasion
2.5.3 过表达或沉默p300 基因对H1299 细胞表达MMP2的影响
细胞转染质粒同前。Western blot 检测结果显示,转染pcDNA3.1/p300 组,MMP2 蛋白表达明显增加(图7A),但在shp300+IL⁃17组,MMP2的表达则显著降低(图7 B)。

图7 Western blot检测过表达或敲低p300对MMP2表达的影响Figure 7 Effects of p300 overexpression and knockdown on MMP2 expression by Western blot
3 讨论
文献报道,无论是SCLC还是NSCLC患者,其肿瘤组织多有炎症微环境的存在[8],而炎症微环境中生成的促炎因子(如IL⁃17)不仅可使支气管上皮细胞发生改变,而且还能加剧细胞的损害,促进肺癌的发生与发展[4-5]。
已知IL⁃17 主要的生物学特征是加剧炎症反应,如诱导更多促炎因子和介质的生成。不过,近年来深入研究IL⁃17 后发现,IL⁃17 既能参与炎症性疾病[9],又可在促进某些肿瘤血管生成、细胞增殖和转移等方面发挥作用[9]。
本研究体外用IL⁃17刺激NSCLC的H1299细胞后进行细胞划痕和Transwell实验发现,IL⁃17刺激能增加细胞的迁移和侵袭,而敲低IL⁃17RA后再行IL⁃17 刺激,其迁移和侵袭能力未见提升。提示,NSCLC 受IL⁃17 刺激时可与其受体结合,最终引发了细胞的迁移和侵袭。
众所周知,细胞行为的变化除了受外界刺激原作用外,还与刺激原诱导的某些转录因子、转录辅激活因子和靶分子的表达紧密相关[10⁃11]。本研究测定受IL⁃17 刺激的H1299 细胞内一些因子变化后发现,转录辅激活因子p300 和MMP2 的表达明显上调,且两者的表达时相基本一致。提示p300 和MMP2升高之间可能存在某种联系。
已知p300 是一含有多个功能结构域的蛋白分子,能作为辅激活因子与转录因子结合,参与靶基因表达的调控[10⁃11]。另MMP2(也称明胶水解酶)是MMP家族的重要成员,它不仅可水解Ⅳ型胶原蛋白和其他生物活性分子,而且还能激活MMP9[10⁃11],故它是肿瘤侵袭与转移重要的调节因子。
为了进一步探讨p300 表达对NSCLC 细胞迁移与侵袭及MMP2 表达的影响,本研究在H1299 细胞内进行了p300 过表达和敲低的相关实验。结果表明,过表达p300后既可促进NSCLC细胞的迁移和侵袭,也能上调MMP2 的表达,而敲低p300 后由IL⁃17诱导的迁移和侵袭能力及MMP2 的表达均显著降低。鉴于IL⁃17诱导NSCLC细胞p300和MMP2的表达高峰时间明显早于细胞的迁移和侵袭。因此推测,p300 的表达上调或许是通过增加MMP2 的生成,最终促进了细胞的迁移和侵袭。不过,这一推测还有待后续的实验加以证实。
猜你喜欢
中国交通信息化(2022年8期)2022-10-28
中学生物学(2022年8期)2022-10-13
中国种业(2022年9期)2022-10-13
——一道江苏高考题的奥秘解读和拓展
中学生物学(2022年7期)2022-09-07
科学与生活(2021年16期)2021-11-25
青岛大学学报(医学版)(2021年5期)2021-11-17
中学生数理化·高一版(2021年12期)2021-09-05
三农资讯半月报(2020年11期)2020-06-21
江苏农业学报(2019年1期)2019-09-10
现代农业科技(2018年13期)2018-10-20
