
基于TGF-β1/Smads信号通路探讨肩袖损伤肌腱止点处异常骨重塑的机制研究
2022-02-24 13:42:08孙孝月李亦丞孙学斌
创伤外科杂志 2022年2期
孙孝月 ,李亦丞,刘 阳,岐 飞 ,孙学斌
1.新疆医科大学第一附属医院运动医学科,乌鲁木齐 830054;2.新疆医科大学第一附属医院关节外科,乌鲁木齐 830054
肩袖损伤是一种常见的肩关节运动损伤疾病,其发病率占肩关节疾病17%~41%,其中冈上肌损伤最为常见[1]。目前,肩关节镜下肩袖修补术是修复肩袖损伤最常用、最有效的治疗方法[2]。然而,术区缝合锚钉在康复训练过程中易出现脱出以及置入位点处骨折等并发症,而且此类问题的处理极为棘手[3]。因此,深入探究肩袖损伤后肌腱止点处异常病理机制势在必行。
研究发现慢性肩袖损伤患者的肱骨大结节处出现异常骨囊肿或骨硬化病理表现[4],而这将严重影响肩袖修复锚钉的缝合强度,减少锚钉与骨组织的接触面积,进而导致脱锚等并发症发生。Wang等[5]发现当肌腱损伤后止点处出现异常骨重塑,该病理机制与TGF-β信号通路高度相关。
转化生长因子β1(transforming growth factor-β1,TGF-β1)是该家族中活性最强的蛋白多肽,超过90%,在全身广泛分布,在组织形成、维持组织内环境稳态方面发挥重要作用[6]。TGF-β1在调节骨代谢功能方面发挥不可替代作用,当关节受到异常机械应力刺激时,埋藏于骨基质中的TGF-β1释放增加,破坏软骨下骨原有的梯度导致异位成骨。异常增加的TGF-β1破坏正常生理状态下受到规律调控的骨重塑,使得骨的合成和吸收形成失偶联[7]。尽管描述了TGF-β1在异常骨重塑中的作用,但TGF-β1在肩袖损伤肌腱止点处异常骨重塑中所扮演的角色尚不清楚。
本实验研究采用SD大鼠动物制造冈上肌肌腱止点处断裂构建肩袖损伤模型,通过染色、免疫组织化学等组织学方法探讨TGF-β1信号通路是否参与肩袖损伤肌腱止点处异常的骨重塑的病理机制,为临床研究治疗肩袖损伤疾病提供新的靶点。
材料与方法
1 实验动物
SPF级雄性SD大鼠40只,12周龄,体质量指数(450±30)g,由新疆医科大学动物实验中心提供,动物合格证号:SCXK(新)2018-0002。实验内容、方法及动物福利等均已获得新疆医科大学第一附属医院实验动物伦理委员会批准(IACUC20191018-14)。
2 主要试剂与仪器
HE染料(中国索莱宝公司);番红固绿染液(美国Sigma公司);TGF-β1抗体(北京博奥森生物技术有限公司);Smad2/3抗体(美国Santa Cruz生物技术公司);成骨转录因子(Osterix)抗体(美国Abcam 公司);抗酒石酸酸性磷酸酶(tartrate-resistant acid phosphatase,TRAP)染色试剂盒、血小板-内皮细胞黏附因子(CD31)抗体均购自中国赛维尔生物科技有限公司;组织脱水机(型号:ASP300S)、石蜡包埋机(型号:EG1150)、病理切片机(型号:RM2135)均购自德国LEICA公司。
3 实验方法
3.1肩袖损伤模型制备 在动物实验平台上铺无菌手术单,SD大鼠采用腹腔注射给予戊巴比妥钠溶液(5mg/100g)进行麻醉,麻醉后进行右肩关节备皮,消毒皮肤。大鼠取左侧卧位,纵行切开右侧上肢皮肤2~4cm,暴露肩胛区上部及上臂肌肉,找到斜方肌附着部并切开此部肌肉,向上翻开即可看见冈上肌。分离冈上肌起止点间,切断少许三角肌充分暴露冈上肌肌腱止点与肱骨大结节连接处,钝性离断冈上肌肌腱在肱骨大结节附着部位。由于本实验是为了制造肩袖损伤模型,故不逐层缝合,标准条件下饲养动物。见图1。

图1 SD大鼠动物肩袖损伤模型制备示意图。a.暴露冈上肌肌腱止点;b.切断冈上肌肌腱止点;c.心脏取血;d.样本取材
3.2动物实验分组 按照随机数字表法分为正常组、模型组术后14、30、60d组,各10只。正常组肩关节不作任何手术操作正常饲养,模型组术后14、30、60d右肩关节在冈上肌肌腱止点与肱骨大结节连接处行冈上肌肌腱切断术建立肩袖损伤模型,造模后于术后14、30和60d人道处死所有动物取右肩关节保留关节囊。见图1。
所有动物正常饲养,可饮水摄食,笼内正常活动。各组动物均先采用镇静和麻醉剂辅助实施安乐死注射,处死前需心脏取全血。从腹部剪开,注意不要剪到脏器,剪破横隔后暴露心脏,输液皮条针头从心尖快速插入,随后依次完成快速全身灌注,灌注液为生理盐水和4%组织固定液,待SD大鼠尾巴僵直后,取材。
4 观察指标
4.1HE染色观察软骨组织病理改变情况 取大鼠右侧保留关节囊的肩关节置于组织固定液中,固定24h,使用10%EDTA-2Na溶液脱钙28d,标本脱水、石蜡包埋;于冈上肌与肱骨大结节冠状面切片机连续切片,厚度为4μm;带有标本的载玻片放置烤箱中常规脱蜡烤片至过夜,以备日后使用。取各组切片行常规HE染色,放置烤箱烘烤1h,脱蜡梯度乙醇分化,HE染色7min,水洗,1%盐酸溶液分色10s,水洗返蓝;伊红染色1min,梯度乙醇脱水,浸入透明液,晾干后中性树胶封片。完成HE染色后,光学显微镜下观察关节软骨组织的形态及结构,测量钙化软骨和透明软骨的厚度,判断潮线是否上移,和病理性改变的程度。
4.2SO-FG染色观察软骨组织退变程度 取每组部分切片,常规烤箱烘烤后浸入脱蜡液中,梯度乙醇分化至水,HE染色1min,水洗:0.2%固绿染液5min,1%冰乙酸10s,0.1%番红O染液5min,水洗:随后置于95%乙醇10s,透明液5min,晾干后中兴树胶封片。完成SO-FG染色后,光学显微镜下观察关节软骨组织破坏情况。
4.3TRAP染色观察软骨下骨破骨细胞活化程度 取每组载玻片放入烤箱烤片,常规脱蜡乙醇梯度,根据TRAP染色试剂盒生产说明书处理4组大鼠切片,然后用甲基绿染液进行复染,晾干后中性树胶封片。通过TRAP试剂盒染色后,光学显微镜下观察软骨下骨中破骨细胞表达情况并进行定量分析。
4.4免疫组织化学法检测TGF-β1、p-Smad2/3、Osterix及CD31表达量 按照免疫组织化学试剂盒生产说明进行实验流程,取带有组织标本的中杉阳离子防脱载玻片,过夜烤片,载玻片浸入脱蜡液梯度乙醇最后至水;3%过氧化氢(H2O2)溶液室温静置15min,阻断内源性过氧化酶的活性,TBST洗3遍,每次5min;0.4%胃蛋白酶放置37℃孵育箱30min抗原修复,TBST洗3遍,每次5min;移液枪滴加山羊血清封闭液40μL,放置37℃孵育箱25min,滴加TGF-β1(1∶100)、p-Smad2/3(1∶40)、Osterix(1∶400)、CD31(1∶400)抗体40μL,4℃冰箱过夜;次日常温40min,TBST洗3次,每次5min;滴加山羊抗兔二抗37℃孵育箱25min,TBST洗3次,每次5min;移液枪滴加DAB显色剂40μL,显微镜下观看组织轮廓由无色渐变为棕黄色立即放入蒸馏水中终止显色;HE染色3min后置于水中清洗,盐酸乙醇溶液1s,PBS溶液中返蓝;梯度乙醇后浸入透明液,晾干后中性树胶封片。完成免疫组织化学法后,采用GraphPad Prism 8软件对软骨下骨中TGF-β1、p-Smad2/3、Osterix及CD31表达量数值进行处理。
5 统计学分析
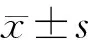
结 果
1 软骨组织病理改变
正常组:关节软骨结构正常,细胞排列分层整齐有序,形态规则,软骨细胞核呈蓝紫色;透明软骨厚度>钙化软骨厚度,潮线清晰;模型组术后14d:关节软骨结构较正常,细胞排列紊乱部分分布不均,透明软骨厚度>钙化软骨厚度;模型组术后30d:关节软骨内细胞紊乱且部分密度不均匀,透明软骨厚度<钙化软骨厚度,潮线上移;模型组术后60d:关节软骨内细胞排列紊乱,钙化软骨明显增厚,透明软骨厚度<钙化软骨厚度,潮线显著上移。与正常组相比,模型组术后30、60d透明软骨厚度均减小,钙化软骨厚度均增加,潮线上移(P<0.05);模型组术后14d透明软骨厚度和钙化软骨厚度与正常组结果比较,差异无统计学意义(P>0.05)。见图2、表1。

表1 各组透明软骨和钙化软骨厚度比值比较

图2 各组软骨组织厚度改变图(HE,×100)。a.正常组:关节软骨结构正常,透明软骨厚度大于钙化软骨厚度;b.模型组术后14d:关节软骨内细胞排列紊乱部分分布不均,透明软骨厚度大于钙化软骨厚度;c.模型组术后30d:关节软骨内细胞紊乱且密度分布不均,透明软骨厚度小于钙化软骨厚度;d.模型组术后60d:关节软骨内细胞排列紊乱,钙化软骨明显增厚,潮线显著上移(图中HC为透明软骨,CC为钙化软骨)
2 软骨组织退变程度
SO-FG染色结果显示:正常组大鼠关节表面结构完整,软骨细胞整体排列有序,番红染色着色较深;模型组术后软骨细胞排列紊乱,番红着色逐渐较浅,表明软骨基质丢失,异常骨质形成。见图3。

图3 各组软骨组织病理学改变图(SO-FG,×200)。a.正常组:关节软骨表面结构完整,番红着色较深;b.模型组术后14d:关节软骨内仅有少量番红染色缺失;c.:模型组术后30d:关节软骨内部分番红着色较浅;d.模型组术后60d:关节软骨内大面积番红染色缺失
3 肩袖止点处破骨细胞活化程度
TRAP染色结果显示:软骨下骨中破骨细胞表达量在模型组14d显著升高,并维持较高水平于术后30d,最终在术后60d恢复至正常水平。模型组术后14、30d破骨细胞表达量高于正常组(P<0.05);与正常组比较,模型组术后60d破骨细胞表达量比较差异无统计学意义(P>0.05)。见图4、5。

图4 肩袖止点处破骨细胞活化程度图(TRAP,×50)。a.正常组:软骨下骨中破骨细胞表达较少;b.模型组术后14d:软骨下骨中破骨细胞表达显著增多;c.模型组术后30d:软骨下骨中破骨细胞表达较正常组明显增多;d.模型组术后60d:软骨下骨中破骨细胞表达量较少(黑箭头所示为破骨细胞)

图5 各组肩袖止点处破骨细胞定量分析图。与正常组比较:*P<0.05
4 肩袖止点处参与异常骨重塑的TGF-β1/Smad2/3信号通路的变化
免疫组织化学染色显示:TGF-β1、p-Smad2/3阳性细胞表达量在模型组术后14、30d中均显著高于正常组(P<0.05);与正常组比较,TGF-β1、p-Smad2/3阳性细胞表达量在模型组术后60d TGF-β1、p-Smad2/3阳性细胞表达量比较差异无统计学意义(P>0.05)。Osterix阳性细胞表达量在模型组术后14、30、60d中呈现递增形式,与正常组比较差异有统计学意义(P<0.05)。CD31阳性细胞表达量在模型组术后14、30、60d显著高于正常组,差异有统计学意义(P<0.05)。见图6、7。

图6 各组软骨下骨中TGF-β1、p-Smad2/3、Osterix及CD31表达情况(免疫组织化学法,×50)。a.正常组:TGF-β1、p-Smad2/3、Osterix及CD31生物学因子阳性细胞表达最少;b.模型组术后14d:TGF-β1和p-Smad2/3生物学因子阳性细胞表达最多;c.模型组术后30d:TGF-β1、p-Smad2/3、Osterix及CD31生物学因子阳性细胞表达较正常组阳性细胞表达较多;d.模型组术后60d:Osterix和CD31生物学因子阳性细胞表达最多

图7 各组软骨下骨中TGF-β1、p-Smad2/3、Osterix及CD31因子定量分析图。a.正常组;b.模型组术后14d;c.模型组术后30d;d.模型组术后60d。与正常组比较:*P<0.05
讨 论
目前探究肩袖损伤的病理机制在肌腱病的组织病理学方面研究较多[8-9],但在肩袖肌腱止点处骨重塑研究较少。肩袖是由冈上肌、冈下肌、小圆肌和肩胛下肌组成的一组肌群,四组肌群呈套袖样包绕在肩关节周围,止于肱骨头大小结节处[10]。其中冈上肌损伤最为常见[1],冈上肌止点即肱骨大结节处,因此本研究采用离断冈上肌肌腱止点构建肩袖损伤模型。动物实验模型是研究疾病病理机制以及预防的必要条件,由于SD大鼠肩关节解剖结构被认为与人类最为相似[11],常用于肩袖损伤机制的研究。因此本研究采用SD大鼠离断冈上肌肌腱止点处,与肱骨大结节处分离开,构建肩袖损伤模型。
正常骨组织重塑是由成骨细胞和破骨细胞之间通过偶联作用完成的,成骨细胞的骨形成与破骨细胞的骨吸收之间的平衡维持了骨稳态[12-13]。在异常骨重塑中,破骨细胞与成骨细胞偶联机制失调,破骨细胞急剧增加,埋藏在骨基质中TGF-β1被释放出髓腔当中,髓腔里的间充质干细胞(MSC)感受众多高浓度TGF-β1后,通过趋化作用,将MSC募集于骨吸收处,导致非偶联重建最终形成骨岛[14]。近期有研究发现异常骨重塑中活化的破骨细胞引起过度的骨吸收,可能参与骨质疏松、类风湿关节炎等不同骨疾病的发病机制[15-16]。Logar等[17]分析了骨关节炎和伴有骨质疏松的髋关节骨折组织中破骨细胞标记物(TRAP)的表达,发现与骨折组相比较,TRAP的表达呈上升趋势。Auréal等[18]对类风湿关节炎在局部骨丢失病理生理学方面进行综述,在类风湿性关节炎患者骨侵蚀部位发现大量破骨细胞,表明破骨细胞在关节骨质退化起着关键性作用。值得注意的是,本研究切断冈上肌肌腱止点后,通过TRAP染色发现在软骨下骨骨吸收异常活跃,模型组术后14d破骨细胞大量活化表现骤升趋势,直到术后60d恢复至正常水平,同时异常骨吸收活跃使得骨基质中TGF-β1大量释放,在软骨下骨中TGF-β1和p-Smad2/3分子阳性细胞表达量随破骨细胞的数量变化而变化。Osterix作为成骨前体细胞的标记物,也随着破骨细胞变化而变化,Osterix阳性细胞数量在术后模型组保持着较高水平,表明软骨下骨中骨的吸收和形成异常活跃,这种异常增加的活性可能会使软骨下骨异常骨重塑加快,进而导致病理机制的发生。
此外,关节软骨和软骨下骨作为肩关节结构的重要组成部分,关节软骨又位于软骨下骨上方,其两者之间相互作用,同时维持着骨重塑内环境的稳态[19]。当软骨下骨发生异常骨重塑时关节软骨也发生着病理学改变。本研究通过HE染色结果显示,在模型组术后30、60d关节软骨内出现细胞紊乱且部分密度不均匀、透明软骨厚度减小,钙化软骨增厚及潮线显著上移病理学改变。此外,本研究通过SO-FG染色结果得出,离断肩袖冈上肌止点构建肩袖损伤模型组,随着时间术后14、30、60d增加,大鼠关节软骨内番红着色逐渐较浅,表明软骨基质丢失逐渐严重,异常骨质形成。另一项研究显示,在异常骨重塑过程中,会伴随着微血管的形成[20]。在本研究中观察术后模型组CD31表达量逐渐增加,这与Lotz[21]发现骨关节炎的病理改变中包括软骨下骨血管形成的增加结果一致,由此可见,在异常骨重塑中往往伴随着异常血管生成。
综上所述,笔者在离断冈上肌肌腱止点构建肩袖损伤模型中观察到肌腱止点处发生异常骨重塑,关节软骨和软骨下骨微观结构发生异常改变,这可能是术后康复训练中术区锚钉脱出发生再次损伤的原因。本研究证实,TGF-β1信号通路参与肩袖损伤肌腱止点处异常骨重塑的病理机制,但本研究中还存在不足之处,没有找到治疗靶点来验证通过Smad2/3介导的转化生长因子β1信号通路参与肩袖损伤肌腱止点处异常骨重塑的病理机制,未来将会继续深入探究。
作者贡献声明:孙孝月:研究实施、数据收集、论文撰写及修改;李亦丞:统计学分析、实验实施、数据分析(收集);岐飞:实验操作、动物饲养;刘阳、孙学斌:研究指导、论文指导及修改、经费支持
猜你喜欢
现代仪器与医疗(2022年3期)2022-08-12 09:04:50
影像科学与光化学(2022年4期)2022-07-19 09:34:46
浙江医学(2022年9期)2022-05-22 13:52:02
运动精品(2022年1期)2022-04-29 08:58:08
福建中医药(2021年1期)2021-12-04 01:12:14
中华养生保健(2020年9期)2021-01-18 03:11:58
中华骨与关节外科杂志(2019年10期)2019-01-03 19:53:28
中国医科大学学报(2018年3期)2018-01-16 20:45:53
腹腔镜外科杂志(2016年9期)2016-06-01 12:10:07
体育科技(2016年2期)2016-02-28 17:06:03
