
芝麻芽期耐铝胁迫相关性状的QTL鉴定
2022-02-15 02:48梁俊超赵云燕王郅琪颜廷献颜小文饶月亮周红英乐美旺
华北农学报 2022年6期
梁俊超,赵云燕,王郅琪,孙 建,颜廷献,颜小文,饶月亮,周红英,乐美旺
(1.江西省农业科学院 作物研究所,江西 南昌 330200;2.国家油料改良中心南昌分中心,江西 南昌 330200;3.江西省油料作物生物学重点实验室,江西 南昌 330200;4.长江大学 农学院,湖北 荆州 330033)
铝毒害是酸性土壤中导致作物减产的最主要因素之一。全世界范围内大约30%的土地和50%的耕地为酸性土,每年造成约6亿美元经济损失[1],我国约有22.7%的国土面积为酸性土壤。近年来,由于酸沉降和铵态氮肥过量施用等原因,我国农田土壤呈现加速酸化的趋势,使得土壤中活性铝离子浓度增大,造成铝毒害增强,严重影响作物生长和产量形成。铝毒害最主要症状表现为抑制根系的伸长,还会改变细胞壁和质膜的结构,影响离子的转运和根系对水分的吸收,导致营养元素失衡,植株生长受到抑制,产量降低[2]。
芝麻(SesamumindicumL.)是世界上最重要的油料作物之一。在我国长江以南的热带和亚热带地区,土壤铝毒害是限制芝麻产量提升的最主要因素之一。虽然通过灌溉措施或直接施用生石灰可减轻酸性土壤中表土的Al3+毒害作用,但这种方法并不能从根本上改善亚表层和底土酸度,成本也较高,还有可能导致农业生态环境破坏,不利于农业可持续发展[3]。培育和种植耐铝胁迫的芝麻品种是解决酸性土壤中铝毒害最为经济有效的措施。了解芝麻耐铝胁迫的遗传基础和定位相关QTL(Quantitative trait loci)对理解芝麻耐铝毒分子机理以及培育耐铝胁迫的芝麻品种具有重要意义。植物对铝胁迫耐受性的遗传较为复杂,既有单位点控制的质量性状遗传模式,又有多基因控制的数量性状遗传模式。禾本科作物的耐铝胁迫性状研究较为深入。比如高粱AltSB位点和大麦Alp控制的耐铝性状符合质量遗传模式[4-5],而Navakode等[6]利用一个大麦双单倍体群体,在染色体2H、3H和4H中鉴定到了多个耐铝胁迫QTL;水稻被认为是禾本科作物中最耐铝毒的作物[7],目前已利用连锁作图和关联分析等方法鉴定得到超过150个耐铝胁迫QTL[7-9],分布于所有(12条)染色体上。
相比于耐旱、耐湿和耐盐等非生物胁迫性状,芝麻耐铝胁迫性状的研究仍处于起步阶段。胡萃等[10]最先报道了铝胁迫对芝麻根系生长的影响,发现一定浓度的铝处理显著抑制芝麻主根伸长,但对侧根发生的影响程度较低。贺根和等[11]研究了铝胁迫对不同芝麻品种根系分泌物的影响,发现在铝处理下,芝麻根系电解质外渗率、氨基酸和糖类的泌出量随铝离子浓度的升高而增加。孙建等[12]对96份芝麻种质资源进行芽期铝胁迫处理,筛选得到一批耐铝胁迫种质资源。然而,目前还没有芝麻耐铝毒相关QTL被报道。
本研究利用前期通过全基因重测序构建的一个包含1 354个标记的高密度遗传连锁图谱[13],对铝胁迫下芝麻芽期的根长、芽长和苗鲜质量进行了调查,利用其相对性状进行了QTL定位,并根据芝麻基因组注释信息筛选和鉴定耐铝相关候选基因,为后续基因克隆和耐铝芝麻品种选育提供理论参考。
1 材料和方法
1.1 植物材料
供试材料包括金黄麻(Jinhuangma,JHM)和竹山白(Zhushanbai,ZSB),以及利用这2个材料,通过单粒传法构建得到的一个包含180个家系的F9重组自交系群体,植物材料繁殖均在初花期采用网帐隔离,以防止昆虫引起的串粉。金黄麻为江西地区广泛种植的耐铝优异地方品种,由江西省农业科学院作物研究所保存,竹山白为湖北省地方品种,对铝胁迫敏感,引自中国农业科学院油料作物研究所。
1.2 耐铝胁迫处理与表型鉴定
首先利用Al2(SO4)3·18H2O配制铝离子质量浓度分别为0,5,10,25,50 mg/L的溶液对金黄麻和竹山白进行芽期铝胁迫处理试验。选取50粒饱满的芝麻种子,利用75%乙醇溶液进行消毒处理,然后用蒸馏水冲洗3遍。将种子置于铺有2层滤纸的带盖发芽盒中(10 cm×10 cm×5 cm),加入40 mL不同浓度的铝溶液,同时设置不含铝离子的水处理作为对照,于28 ℃恒温培养箱中暗培养。处理5 d后,每个材料选择20株长势相近的植株,进行根长性状的测量以及相对根长的计算,以筛选铝胁迫处理适宜浓度。
利用筛选到的适宜浓度铝溶液对重组自交系群体进行萌发胁迫处理试验,以蒸馏水处理作为对照,每个家系选取50粒饱满种子,试验重复3次。处理5 d后从发芽盒中随机选取20株长势一致的植株调查根长、芽长和苗鲜质量等性状,并计算各性状相对值,即胁迫处理下性状测定值与水对照处理下性状测定值的比值,分别为相对根长(Relative root length,RRL)、相对芽长(Relative shoot length,RSL)和相对苗鲜质量(Relative fresh seedling weight,RSW)。3次重复的平均值用于后续遗传分析和QTL定位。采用SPSS软件(SPSS Inc.,Chicago,IL)进行数据统计学分析。
1.3 QTL定位分析
用于QTL定位的高密度遗传图谱由Liang等[13]前期通过全基因组重测序构建完成。利用Window QTL Cartographer v2.5[14](http://statgen.ncsu.edu/qtlcart/WQTLCart.htm)软件中的复合区间作图法(CIM,Composite interval mapping)进行目标性状QTL定位,采用标准模型6,窗口大小为10 cM,LOD阈值设为2.5。
1.4 候选基因分析
根据中芝13参考基因组Sinbase 2.0[15](http://www.sesame-bioinfo.org/Sinbase2.0/)的注释信息提取所有QTL定位区间内预测基因的蛋白序列,将这些预测基因蛋白序列BlastP比对拟南芥基因组数据库和NCBI蛋白库,结合文献检索,对基因功能进行预测,筛选潜在候选基因。
2 结果与分析
2.1 亲本金黄麻和竹山白对铝胁迫的耐受性
为了明确用于重组自交系群体铝胁迫处理的铝离子溶液浓度,利用亲本金黄麻和竹山白进行了芽期铝离子浓度梯度胁迫处理试验。在5,10,25,50 mg/L铝离子质量浓度处理下,金黄麻幼苗的根长分别被抑制了3.05%,15.86%,42.37%和84.43%,而竹山白的根长分别被抑制了3.92%,12.08%,81.43%和91.79%,这表明在低质量浓度(5,10 mg/L)的铝离子胁迫下,金黄麻和竹山白根系生长受到的抑制程度相当。在铝离子质量浓度升高到25,50 mg/L时,相比于竹山白,金黄麻对铝胁迫更不敏感,其中铝离子质量浓度为25 mg/L时,二者根长差异最大(图1)。因此,本研究选择25 mg/L作为芝麻芽期铝胁迫处理的质量浓度。
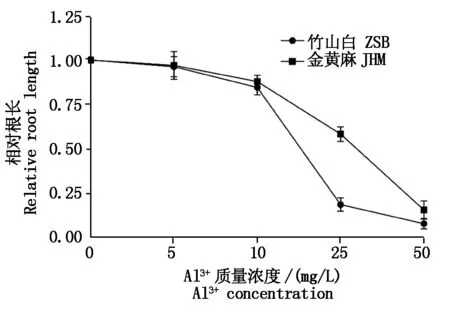
图1 不同铝离子浓度对金黄麻和竹山白相对根长性状的影响Fig.1 Effects of different Al3+ concentrations on relative root length of JHM and ZSB
2.2 铝胁迫下重组自交系群体的表型分析
利用铝离子质量浓度为25 mg/L的溶液对亲本以及包含180个家系的重组自交系群体进行芽期胁迫处理,同时设置水为对照,发芽后5 d调查相对根长、相对芽长和相对苗鲜质量3个性状。结果表明,在铝离子溶液处理下,3个指标中相对根长的数值最小,其次为相对芽长和相对苗鲜质量,说明双亲根和芽的生长均受到不同程度抑制,铝对根长的抑制作用最大,而芽长和苗鲜质量的抑制程度相对较轻。在RIL群体中,3个相对性状均呈现连续分布,且均有超金黄麻亲本的分离现象(表1、图2)。相关性分析结果表明,在RIL群体中3个相对性状之间均表现为极显著正相关(表2),相对根长和相对芽长之间的相关系数最高。
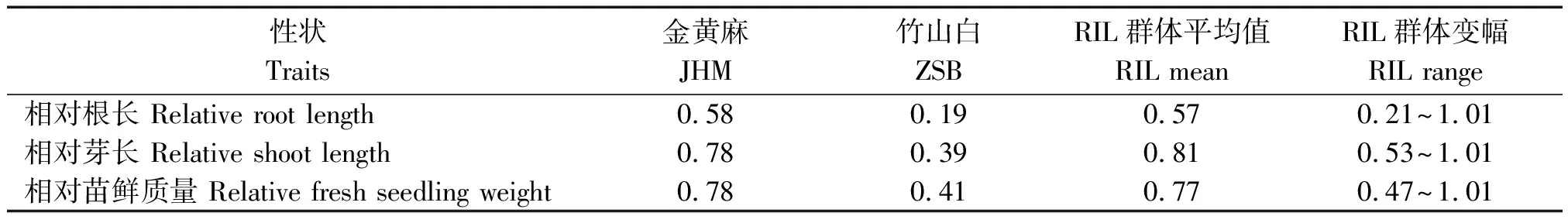
表1 25 mg/L铝离子浓度胁迫下亲本和RIL群体的各性状表现Tab.1 Phenotypic data of three traits in parents and RIL population under 25 mg/L Al3+ treatment

表2 RIL群体中3个耐铝相关性状的相关性分析Tab.2 Correlation coefficients for three Al-tolerant traits in the RIL population
2.3 芝麻芽期耐铝胁迫性状QTL定位
在前期研究中,对一个以金黄麻和竹山白为亲本构建的包含180个家系的F9群体进行了高通量测序,并构建得到一个由13个连锁群组成的、包含1 354个bin标记的高密度遗传图谱[13]。本研究利用该图谱对耐铝胁迫相关性状进行QTL定位,共检测到7个与相对根长、相对芽长和相对苗鲜质量相关的QTL,分布于第2,4,6,10,11,13号染色体上。这些QTL的LOD值为2.55~3.55,解释5.46%~7.75%的表型变异。

黑色箭头和灰色箭头分别表示金黄麻和竹山白的平均值。Black and grey arrows represent mean of JHM and ZSB,respectively.
3个QTL与相对根长性状相关,分别位于第2,10,11号染色体上,解释5.46%~7.01%的表型变异,其中位于2号染色体上的qRRL02效应最大(表3);相对芽长性状共检测到了2个QTL,分别位于6,13号染色体上,分别解释7.75%和6.68%的表型变异;相对苗鲜质量性状同样检测到2个位点,分别位于2,4号染色体上,分别解释5.85%和5.95%的表型变异。除与相对根长相关的qRRL10以及与相对苗鲜质量性状相关的qRSW04外,其余QTL的增效等位位点均来源于金黄麻。此外,位于2号染色体上的控制相对根长性状QTLqRRL02(c02b009~c02b013)与控制相对苗鲜质量的qRSW02(c02b008~c02b010)所在的基因组区域相近。
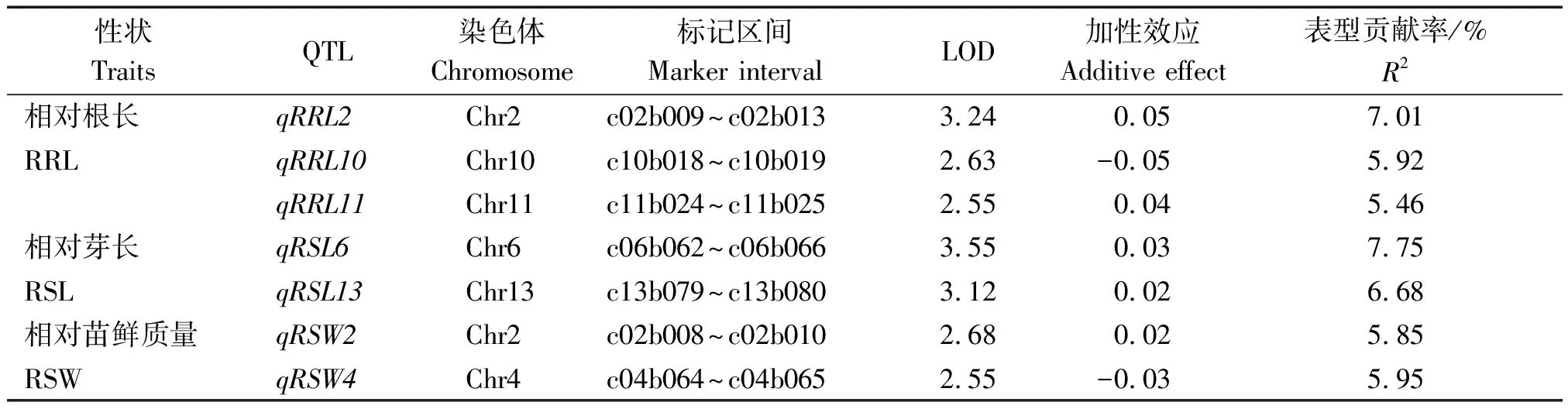
表3 3个耐铝胁迫性状相关QTL的鉴定Tab.3 The identification of QTL associated with three Al-tolerant traits
2.4 候选基因筛选
利用芝麻参考基因组序列,对QTL标记区间内的候选基因进行注释,并通过与拟南芥蛋白数据库和NCBI蛋白库比对,分析候选基因的功能。7个QTL基因组区域中共有364个预测基因,其中303个基因在拟南芥中存在同源基因,74个基因被注释为未知蛋白、假定蛋白或重复序列。功能注释和文献检索的结果表明,除qRRL11外,其余QTL区域共筛选到15个可能与耐铝毒相关的候选基因(表4),分布于5条染色体上。
相对苗鲜质量QTLqRSW02和qRSW04区域共筛选到6个候选基因,SIN_1007432、SIN_1007469、SIN_1007434、SIN_1012986、SIN_1013011和SIN_1013013,分别编码重金属转运与脱毒家族蛋白、过氧化物酶体过氧化氢酶、木质部半胱氨酸肽酶1、组蛋白赖氨酸N-甲基转移酶SETD1B类蛋白、紫色酸性磷酸酶22以及紫色酸性磷酸家族蛋白。研究表明,这些蛋白在拟南芥等其他植物中的同源蛋白参与介导了植物重金属转运、毒性离子积累、细胞程序性坏死、细胞壁生物合成以及金属离子结合和去磷酸化等各种非生物胁迫防御生理进程[16-21]。
控制相对根长的QTL区间内共鉴定到5个候选基因,除qRRL02区域中与相对苗鲜质量QTLqRSW02区域重叠的3个基因外,在qRRL10区间内还筛选到2个候选基因,SIN_1025967和SIN_1025968,分别编码膜结合内切-1,4-β-D-葡聚糖酶和谷胱甘肽转移酶。SIN_1025967在拟南芥中的同源基因AT5G49720是细胞伸长和分裂相关基因,在耐铝毒突变体grp3-1根系中优势表达[22]。SIN_1025968的拟南芥同源基因AT2G02380则被证明参与铝胁迫诱导产生的ROS的清除[23]。
2个相对芽长QTL区间内则筛选到7个候选基因,6个位于qRSL06区间,1个位于qRSL13区间(表4)。SIN_1018446、SIN_1018450、SIN_1018441、SIN_1018445、SIN_1018453、SIN_1018481和SIN_1003975这7个候选基因分别编码乳酸/苹果酸脱氢酶家族蛋白、GDSL基序酯酶、假定脂质转运蛋白、非特异性脂类转运蛋白(nsLTP)、细胞色素P450、丝氨酸蛋白酶以及WRKY DNA结合蛋白75。其中6号染色体上的SIN_1018446编码乳酸/苹果酸脱氢酶家族蛋白,在拟南芥中的同源基因AT4G17260,又名CMDH4,参与了对重金属铅胁迫的耐受性[24];SIN_1018450编码GDSL基序酯酶,其拟南芥同源物AT1G29670参与了非生物胁迫下ROS信号介导的系统获得性适应机制[25];SIN_1018441与AT4G12480同源,编码一种假定脂质转运蛋白,与铝毒、低温、盐和ABA等多种非生物胁迫抗性有关[26];SIN_1018445与拟南芥AT2G45180同源,编码一类非特异性脂类转运蛋白(nsLTP),该基因受铝胁迫诱导下调表达,也可通过影响对ABA的敏感性来调控盐胁迫抗性[27]。SIN_1018453为细胞色素P450家族成员,其拟南芥同源基因AT5G07990属于金属结合跨膜P450细胞色素类型[18];SIN_1018481编码一个丝氨酸蛋白酶类蛋白,与拟南芥At1g04110同源,该蛋白通过输出到质外体与质膜结合,参与胞外信号的接收和传导[28]。位于13号染色体qRSL13区域内的SIN_1003975编码一个WRKY转录因子,与拟南芥WRKY75同源,该基因可通过水杨酸途径调控植物对盐、干旱等胁迫的应答[29]。
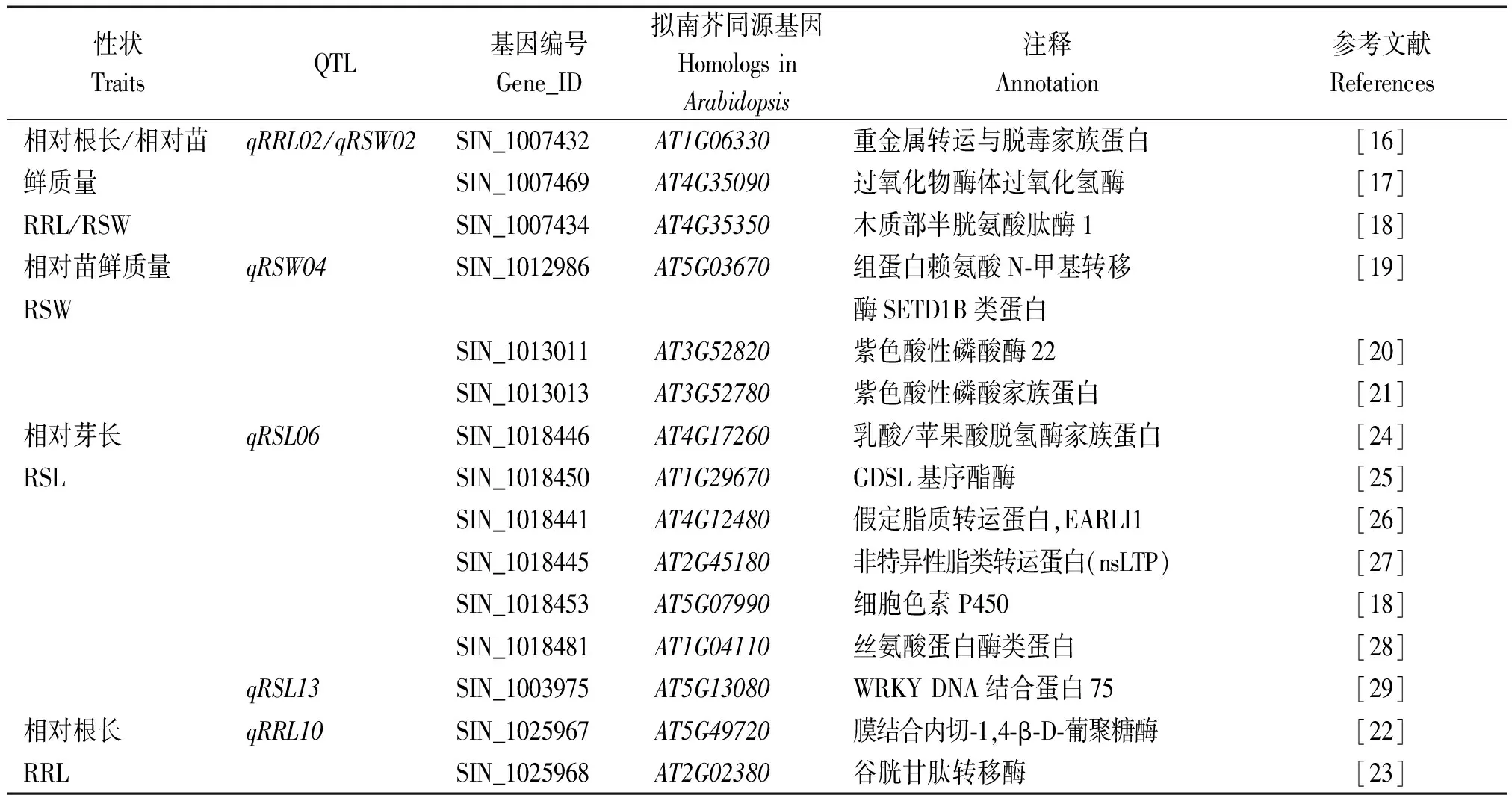
表4 耐铝胁迫QTL区间内的预测候选基因Tab.4 Putative candidate genes in the interval of major QTLs for Al-tolerance
3 结论与讨论
土壤铝毒害是限制我国南方地区芝麻产量的重要因素之一。不同基因型芝麻品种耐铝性存在显著差异。在前期的研究中,已有一些关于铝毒对芝麻根系影响以及芝麻耐铝种质筛选的研究报道[10-12]。然而,作物的耐铝性是一个复杂的数量性状,目前还未有关于芝麻耐铝胁迫QTL定位和遗传机制研究的报道。在本研究中,通过铝浓度梯度胁迫处理试验,发现江西地方品种金黄麻表现出较好的芽期铝胁迫耐受性(相比于竹山白),并确定了25 mg/L的Al3+溶液作为芝麻芽期铝胁迫处理的浓度。利用相对根长、相对芽长和相对苗鲜质量3个指标进行了亲本和重组自交系群体芽期耐铝性的表型评价,通过前期构建的高密度遗传图谱对芝麻芽期耐铝性进行了QTL定位。共鉴定到7个与耐铝胁迫性状相关的QTL,分布于6条染色体上。
对于作物耐铝育种来说,虽然直接在酸性土壤中进行田间试验是最理想的表型鉴定方法,但地块间不一致的土壤性质和不可控的环境条件会显著降低表型数据的准确性[30-31]。因此,利用铝溶液进行胁迫处理试验被广泛用于植物耐铝性的鉴定和QTL定位,特别是对于植株的苗期和芽期。例如王瑞莉等[32]以80 μg/mL的铝胁迫浓度对一个甘蓝型油菜重组自交系群体进行种子萌发试验,鉴定到23个与相对根长、芽长和干质量以及相对发芽势和发芽率相关的QTL;Kobayashi等[33]以相对根长为表型指标,通过水培的方法在2个拟南芥重组自交系群体中分别定位到2个QTL;在大豆中,Sharma等[34]利用苗期水培铝胁迫处理试验,在一个重组自交系群体中鉴定到2个与相对根长有关的主效QTL。本研究采用萌芽期铝溶液胁迫处理的方法,对一个包含180个家系的芝麻重组自交系群体进行了相对根长、相对芽长和相对苗鲜质量的性状调查,3个性状在群体中均表现出广泛的表型变异,适合QTL定位。利用前期构建的高密度遗传图谱,定位到7个与耐铝胁迫相关的QTL。本研究系首次在芝麻中定位耐铝胁迫相关QTL。此外,在2号染色体的2个相近重叠区域中分别鉴定到一个相对根长QTLqRRL02(c02b009~c02b013)和一个相对苗鲜质量QTLqRSW02(c02b008~c02b010)。控制2个或2个以上耐铝相关性状QTL成簇分布的现象在大豆[34]和油菜[32]等其他作物中也有报道,这可能是由于遗传连锁或一因多效引起。
目前,已发掘的植物耐铝等金属离子胁迫相关基因主要参与2种生理进程:一种是通过介导分泌有机酸(比如苹果酸和柠檬酸)来螯合Al3+等金属离子,阻止其进入根尖细胞;另一种则是通过调控Al3+等金属离子的转运,将其隔离在胞内液泡中,使其远离对金属离子敏感的组织,减轻铝的毒害[35]。此外,一些响应逆境胁迫的信号分子途径相关基因也被证明参与了植物对铝胁迫的耐受性[36-37]。利用芝麻参考基因组信息,本研究对耐铝胁迫QTL区域内的候选基因进行了分析,在7个QTL区域内共鉴定到15个可能与耐铝胁迫有关的候选基因。这些基因中有的参与了金属转运、脱毒(比如SIN_1007432),有的基因的拟南芥同源物被报道参与金属离子的跨膜结合(比如SIN_1018453),也有的参与植物响应铝毒、干旱和盐等非生物胁迫的信号接收或传导(比如SIN_1018450、SIN_1018481和SIN_1003975)。此外,目标区域中还鉴定到一些与多种逆境胁迫耐受性有关的蛋白,比如位于6号染色体qRSL06区域内的2个脂质转运蛋白,这类基因通常在维持细胞膜和细胞壁的稳定性中起重要作用,其中SIN_1018441的拟南芥同源基因AT4G12480,编码一个位于细胞壁的富含脯氨酸结构域的分泌性脂质转运蛋白,又名EARLI1(early Arabidopsis aluminum induced 1),该基因由于迅速响应铝胁迫诱导而得名,同时还与低温、盐和ABA等非生物胁迫抗性有关[26]。10号染色体qRRL10区域中的SIN_1025968,编码一种谷胱甘肽转移酶,该蛋白酶参与多种生物体内有毒物质的代谢与解毒,SIN_1025968的同源基因AT2G02380在铝毒敏感突变体stop1中受铝胁迫诱导显著上调表达,而在野生型中其表达水平不受铝毒诱导[23]。研究表明,过表达谷胱甘肽转移酶基因可显著提高植株对铝毒的耐受性,该类基因参与了铝毒胁迫下细胞中诱导产生的ROS清除[38]。
在4号染色体qRSW04区域内发现2个物理距离相近的紫色磷酸酶基因SIN_1013011和SIN_1013013,与拟南芥AT3G52820(AtPAP22)和AT3G52780(AtPAP20)相似性最高,AtPAP22基因位点还与耐酸性胁迫性状关联[20-21]。候选基因的同源基因在其他作物耐铝胁迫QTL定位研究中同样被鉴定到的情况还有SIN_1018453和SIN_1007434,分别位于qRSL06区域和qRRL02/qRSW02区域内,SIN_1018453与拟南芥AT5G07990同源,编码与金属离子结合有关的跨膜细胞色素P450蛋白,而SIN_1007434编码木质部半胱氨酸肽酶1,该蛋白酶具有引导细胞凋亡的功能,参与调控了金属解毒过程中可能具有发育衰老特异性细胞的死亡[18,39]。SIN_1018453和SIN_1007434在臂形草中的同源基因也位于其耐铝QTL区域中,且均在抗、感材料中受到铝胁迫诱导差异表达[18]。本研究发掘的芝麻耐铝QTL以及相关候选区域中的基因和连锁标记为芝麻耐铝分子标记辅助选择和功能基因克隆提供了潜在的遗传资源。
猜你喜欢
汉字汉语研究(2021年2期)2021-08-30
现代园艺(2020年7期)2020-04-22
湖北农业科学(2019年20期)2019-12-11
汉字汉语研究(2019年2期)2019-08-27
农业与技术(2018年5期)2018-04-25
新高考·英语进阶(高二高三)(2018年8期)2018-01-15
农产品加工(2017年19期)2017-11-14
上海农业学报(2017年3期)2017-04-10
河北书画研究(2016年3期)2016-04-28
红领巾·探索(2015年9期)2015-09-10
