
大豆GmRPK2基因的克隆及功能初步鉴定
2022-02-15 02:55顾哑慧张常青张雪冰张鑫月张鑫杰葛荣朝
华北农学报 2022年6期
顾哑慧,张常青,张雪冰,张鑫月,张鑫杰,葛荣朝
(河北师范大学 生命科学学院,河北 石家庄 050024)
植物生长发育过程中会遇到各种环境的胁迫,其中外环境因素主要有水、温度、光照等,植物激素则是主要的内环境因素,如ABA、生长素、细胞分裂素、乙烯、GA、茉莉酸和油菜素内酯等。植物对于这些环境因素变化做出适当的反应,是其正常生长发育的必要条件[1]。
植物在受到非生物胁迫后,往往会激发自身细胞中的精密信号传导网络,调控相应基因的表达,以应对胁迫带来的细胞生理变化。类受体激酶(Receptor-like protein kinases,RLKs)就是这个精密网络中的一类能够感知环境信号、放大信号传递,使细胞对逆境做出应答的蛋白激酶[2]。类受体蛋白激酶是结构类似于动物体内受体蛋白激酶的一类从植物中发现的激酶[3]。植物类受体蛋白激酶基因种类繁多,其中拟南芥中有大约610个,约占拟南芥所有蛋白质编码基因的2.5%[2]。类受体激酶一般包括胞外受体结构域(Extracellular ligand binding domain,ECLB)、跨膜结构域(Transmembrane domain,TM)和胞内激酶域(Protein kinase catalytic domain,PKC)3个结构域[4]。
根据其胞外域的结构不同,植物类受体蛋白激酶可以分为6个亚家族,本试验选择的大豆类受体激酶GmRPK2属于富含亮氨酸重复序列的类受体蛋白激酶(Leucine-rich repeats,LRRs)[5-7]。LRRs在植物激酶中几乎占到50%,在植物代谢、生长和发育中发挥重要作用[8-9]。水稻凝集素类受体蛋白激酶OsLPK1的突变体表现为白化,这表明该激酶可能与植物叶片的发育有关[10]。拟南芥LRR类受体激酶GSO1和GSO2对于拟南芥胚胎发育及气孔分布起着重要作用[11]。拟南芥受体激酶CLV1则参与调控茎的生长和花的数量[12]。另外,LRR类受体激酶还参与水稻的产量和分枝调节、拟南芥莲座叶和花的发育以及植物的器官程序性死亡调控[13-15]。
LRRs类受体激酶在植物抵御非生物胁迫应答方面也起着重要的作用[16-21]。Osakabe等[22-23]研究表明,在拟南芥生长发育的过程中,AtRPK1基因的表达量降低会导致植株对ABA的敏感性显著降低。AtRPK1、OsRPK1利用RNA干涉降低表达量后,可明显提高拟南芥、水稻对盐胁迫的耐受性[24-25]。
大豆GmRPK2基因与AtRPK1有着高度的同源性,本研究拟对其抗逆功能进行初步研究,以探究GmRPK2过量表达后对植物高盐、干旱等非生物胁迫耐受性的影响,为之后研究大豆抗逆新品系及培育奠定基础。
1 材料和方法
1.1 试验材料
拟南芥(Arabidopsisthaliana)为Columbia型,大豆(Glycinemax)品种为冀豆22,大肠杆菌DH5α、根癌农杆菌GV3101、表达载体pCAMBIA1300和p2300-GFP由河北师范大学遗传学实验室保存。
1.2 载体构建与拟南芥转化及筛选
利用大豆叶片提取总RNA,反转录获得cDNA。以cDNA为模板,采用高保真PrimeSTAR酶,利用基因特异引物FP: 5′-GTCTAGAGAACAAGGGTTTTGG
GTCTCAC-3′(酶切位点XbaⅠ)和RP: 5-′CGGTACC
CACACAAGGGGCTAGCATGATG-3′(酶切位点KpnⅠ)进行PCR扩增。扩增产物进行琼脂糖电泳后,将GmRPK2电泳条带进行切胶、回收纯化并连接到pMD18-T载体,转入大肠杆菌。测序准确后的质粒利用XbaⅠ、KpnⅠ进行酶切、回收,将GmRPK2连接到表达载体pCAMBIA1300。酶切鉴定后将p1300-35S-GmRPK2重组载体转化到农杆菌GV3101中。
另外,利用pMD18-T-GmRPK2质粒作为PCR模板,用引物GFP-RP:5′-CGGTACCGAACAAGGGTTT
TGGGTCTCAC-3′(酶切位点XbaⅠ)和GFP-LP:5′-GTCTAGAGCATGATGGGGGTTGAAGTTGCTT-3′(酶切位点KpnⅠ)扩增得到不含终止密码子的GmRPK2基因序列,构建p2300-GmRPK2-GFP载体,转入农杆菌GV3101。
摇培含有p1300-35S-GmRPK2、p2300-GmRPK2-GFP载体的农杆菌,利用浸花法转化拟南芥,潮霉素(25 mg/L)筛选获得阳性苗,单株收获种子继续筛选,选取阳性苗占比为3/4的株系继续培养,单株收种后筛选得到GmRPK2过表达拟南芥纯合体。
1.3 GmRPK2过表达拟南芥对ABA、干旱和盐胁迫的抗逆性检测
选取野生型和3个GmRPK2过表达拟南芥纯合体株系种子,利用70%酒精和次氯酸钠进行消毒,分别点播到含有10% PEG、1.5 μmol/L ABA、125 mmol/L NaCl的MS固体培养基和对照MS培养基,4 ℃春化3 d,转至22 ℃光照培养箱中培养,统计萌发率,试验进行3次生物学重复。
同样选择3个GmRPK2过表达株系和野生型拟南芥种子,消毒后点播到MS固体培养基,4 ℃春化3 d,转到22 ℃光照培养箱中竖直培养3 d,选择长势、根长一致的幼苗分别转移至含有10% PEG、1.5 μmol/L ABA、125 mmol/L NaCl的MS固体培养基和对照MS培养基上,继续竖直培养5 d,对根相对生长量进行测量与统计,试验进行3次生物学重复。
另外,在MS固体培养基上铺布3个过表达株系和野生型拟南芥种子,4 ℃春化3 d,恒温光照培养箱培养10 d,幼苗转移至营养土中,光照培养14 d。选取生长状况一致的过表达株系和野生型植株,用175 mmol/L NaCl胁迫处理,每隔3 d浇一次盐水,胁迫处理18 d,观察其生长状况。另外选取一批长势一致的过表达株系和野生型植株,停止浇水,干旱处理14 d,观察其生长状况。
1.4 转基因拟南芥的生理检测
3个过表达株系和野生型拟南芥正常光照培养14 d后,取长势一致的植株,停止浇水14 d作为干旱处理组,175 mmol/L NaCl溶液浇灌18 d作为盐胁迫处理组,清水浇灌作为对照组。取叶片进行细胞质膜透性、丙二醛含量、叶绿素含量、脯氨酸含量、可溶性糖含量和离体叶片失水率等生理指标检测。植物叶片中过氧化氢积累量观察采用DAB组织染色法进行测定。
细胞质膜透性采用电导率法进行测定,每次取样称质量0.2 g拟南芥叶片,剪成适当小块,向离心管加入10 mL蒸馏水将叶片尽可能压入水中,真空抽气1 h后,室温搅动浸提1 h后,测定样品电导率。然后100 ℃水浴15 min,冷却至室温,测定各种样品的煮沸电导率。
相对电导率=(处理电导率-空白电导率)/(煮沸电导率-空白电导率)×100%。
1.5 GmRPK2的表达对拟南芥抗逆信号通路相关基因的表达量影响
选取定位于植物抗逆相关信号通路下游基因SAD1、FRY1、SOS3、ESK1、RD29B、ADH1和P5CS1。设计相应引物,以β-Actin基因作为内参,对野生型拟南芥和GmRPK2过表达拟南芥植株进行荧光定量PCR检测(表1)。
2 结果与分析
2.1 非生物胁迫对GmRPK2基因表达的影响
取正常培养14 d的大豆幼苗进行的120 mmol/LNaCl、1.5 μmol/L ABA、5% PEG胁迫0,1,6,12 h,取叶片进行qRT-PCR检测。结果表明,GmRPK2在盐胁迫1 hGmRPK2基因表达量明显上升,而在6,12 h出现下降趋势,PEG、ABA处理后GmRPK2基因表达量同样也在1 h有所上升,随后出现回落(图1)。

表1 qRT-PCR引物碱基序列Tab.1 qRT-PCR primer sequences

不同小写字母表示差异显著(P<0.05)。图4—5,7—9同。Different lowercase letters indicate significant differences(P<0.05).The same as Fig.4—5,7—9.
2.2 GmRPK2亚细胞定位观察
构建得到p2300-GmRPK2-GFP质粒,以含有空载p2300-GFP质粒的农杆菌为对照组,含有p2300-GmRPK2-GFP质粒的农杆菌为试验组,注射烟草叶片,36~48 h内观察GmRPK2表达蛋白的亚细胞分布情况。其中注射空载质粒烟草叶细胞的细胞核及细胞膜中均可激发绿色荧光,但试验组仅细胞膜具有绿色荧光蛋白,由此推测GmRPK2蛋白主要分布在细胞膜上(图2)。

图2 GmRPK2亚细胞定位Fig.2 Subcellular localization of GmRPK2 fusion protein
2.3 GmRPK2转基因株系的分子水平鉴定
提取野生型和转基因过表达拟南芥的基因组DNA,同时提取总RNA,反转录为cDNA。以DNA作为模板,利用FP和RP引物进行PCR扩增,筛选获得成功转入外源基因GmRPK2的3个转基因株系OX1、OX2、OX3(图3-A)。对其cDNA同样利用FP和RP引物进行RT-PCR鉴定,确认GmRPK2基因在这3个拟南芥株系中均实现表达(图3-B)。

A.基因组PCR检测;B.RT-PCR检测;M.DL2000 DNA Marker。A.PCR detection of genome;B.RT-PCR detection;M.DL2000 DNA Marker.
2.4 GmRPK2转基因株系的抗逆性分析
2.4.1 转基因种子萌发过程中的抗逆性检测 对GmRPK2过表达拟南芥的3个纯合体株系OX1、OX2、OX3和野生型拟南芥种子进行消毒处理,点播在含有ABA、PEG和NaCl的MS固体培养基及对照培养基上。萌发率检测结果表明,对照培养基的3个过表达株系种子及野生型种子的萌发情况基本一致。在1.5 μmol/L ABA培养基上4 d后,野生型萌发率为85.56%,3个转基因株系萌发率分别为12.22%,15.56%和14.44%;在125 mmol/L NaCl培养基上培养4 d后,野生型拟南芥萌发率为70.00%,而3个过表达株系萌发率分别仅为10.00%,11.11%和8.89%。在10% PEG培养基上4 d后,野生型萌发率为84.44%,3个转基因的株系萌发率分别为21.11%,22.22%和22.4%(图4)。
由此可见,GmRPK2过表达株系在ABA、NaCl和PEG胁迫下的萌发率均显著低于野生型(P<0.05),表现出对非生物胁迫更加敏感。

图4 GmRPK2过表达拟南芥逆境胁迫下的萌发率检测Fig.4 Germination rate of GmRPK2 overexpressed Arabidopsis under stress
2.4.2GmRPK2转基因拟南芥幼苗根相对生长量的抗逆性检测 对3个GmRPK2过表达株系OX1、OX2、OX3和野生型拟南芥种子消毒处理,在MS培养基上垂直培养3 d,挑选根长度一致的幼苗分别转移到含有1.5 μmol/L ABA 、10% PEG和125 mmol/L NaCl的MS培养基。竖直培养5 d后,可发现在3种胁迫条件下,GmRPK2过表达拟南芥和野生型拟南芥的根生长都明显受抑,过表达拟南芥根的生长受抑制更加明显(图5)。因此,对于各种非生物胁迫,GmRPK2过表达拟南芥较野生型在幼苗根的生长发育方面表现得更加敏感。

图5 GmRPK2过表达拟南芥逆境胁迫下根的生长发育检测Fig.5 Root growth and development of GmRPK2 overexpressed Arabidopsis under stress
2.4.3GmRPK2过表达拟南芥盐胁迫及旱胁迫的抗逆性检测 选取培养14~21 d、长势状况一致的GmRPK2过表达株系和野生型拟南芥植株,进行盐、旱胁迫处理。盐胁迫采用175 mmol/L NaCl溶液处理,每隔3 d浇灌一次。经过18 d处理,结果表明,在NaCl胁迫下,GmRPK2过表达拟南芥叶片枯萎的更加严重,茎秆萎蔫的更为厉害,幼嫩的茎尖部位严重萎蔫,抽薹高度也明显缩短(图6)。干旱处理14 d后,过表达拟南芥茎秆叶片明显枯萎,侧枝、果荚数量明显减少,整体长势较弱,而野生型植株整体长势相对较好。由此可见,GmRPK2过表达使拟南芥成株对盐、旱等非生物胁迫也表现出更加敏感的表型。
2.4.4GmRPK2过表达拟南芥盐胁迫后的生理指标检测GmRPK2过表达株系和野生型拟南芥正常培养14 d后,利用175 mmol/L NaCl处理18 d,检测其叶片的叶绿素含量、丙二醛含量、过氧化氢积累和相对电导率等生理指标。试验结果表明,在盐胁迫后GmRPK2过表达植株的丙二醛含量显著高于野生型,说明过表达植株所受细胞膜质过氧化的损伤程度更为严重(图7-A)。过表达拟南芥的相对电导率相比野生型显著增高(图7-B),表明过表达植株的脂膜透性显著大于野生型,这可能导致更多有害离子渗入胞内,进而造成细胞受损更加严重。盐胁

图6 GmRPK2过表达拟南芥植株对高盐和干旱的抗逆性Fig.6 Resistance of GmRPK2 overexpressed Arabidopsis adult plants to high-salt and drought
迫后过表达植株的叶绿素含量下降更为明显,提示其光合系统受到的损害更为严重(图7-C)。用DAB组织染色法对盐胁迫前后叶片中过氧化氢积累情况进行观察,结果显示,过表达植株在盐胁迫后其叶片出现更多的红褐色斑点积累,提示其过氧化氢积累更多,叶片遭受的氧化胁迫更为严重(图7-D)。
2.4.5GmRPK2过表达拟南芥旱胁迫后的生理指标检测 野生型和GmRPK2过表达拟南芥株系OX1、OX2、OX3正常培养14 d后,取生长状况一致的野生型和转基因植株进行干旱处理,14 d后测定其叶片的脯氨酸和可溶性糖含量。结果表明,干旱胁迫后的过表达植株叶片的脯氨酸和可溶性糖含量均显著低于野生型(图8-A、B)。
分别剪取在苗室正常培养28 d的野生型和GmRPK2过表达拟南芥的叶片进行离体失水率测定,结果表明,GmRPK2转基因植株叶片的失水率要大于野生型植株叶片的失水率,表明转基因植物离体叶片的水分散失要大于野生型,这可能也是其对干旱胁迫更加敏感的原因之一(图8-C)。
2.5 GmRPK2过量表达对下游基因的影响
为探究GmRPK2基因在拟南芥中过表达后造成其对盐胁迫的耐受性下降原因,选取了位于CDPK、SOS、MAPK等3个信号通路下游的5个基因,以野生型拟南芥和GmRPK2过表达拟南芥的cDNA为模板进行RT-PCR检测。结果表明,GmRPK2在拟南芥中的过表达显著抑制了CDPK信号转导通路下游的基因SAD1和脯氨酸合成相关基因P5CS1的表达。另外,GmRPK2的过表达也抑制了ABA信号转导途径ADH1基因的表达(图9)。

图7 盐胁迫后GmRPK2过表达拟南芥的生理指标检测Fig.7 Physiological index of GmRPK2 transgenic Arabidopsis under salt stress
3 结论与讨论
植物类受体蛋白激酶(Receptor-like protein kinases,RLKs)在植物生长发育、应对生物胁迫及非生物胁迫中起到重要的作用。迄今,水稻中已经发现大量和抗逆相关的类受体激酶,例如水稻的Ca2+依赖蛋白激酶基因OsCDPK7过表达后,转基因水稻对盐胁迫和干旱胁迫表现出更强的耐受性,但耐冷性并未提高[26]。此外,水稻的类受体胞质激酶基因GUDK的表达量与干旱胁迫、盐胁迫(200 mmol/L NaCl)、热激(45 ℃)和冷激(4 ℃)等都表现出了不同程度的相关性[27]。GmRPK2基因和已知的抗逆相关基因AtRPK1、OsRPK1高度同源,本研究初步探究了GmRPK2基因对拟南芥耐旱性、耐盐性的影响及其作用机理。

图8 GmRPK2过表达拟南芥旱胁迫后的生理指标检测Fig.8 Physiological indicators of GmRPK2 transgenic Arabidopsis under drought stress
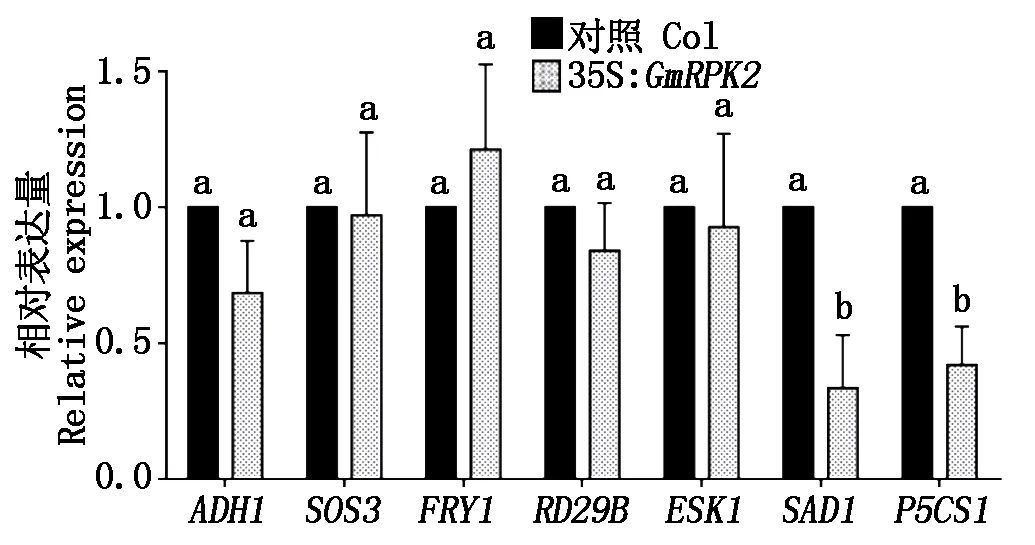
图9 GmRPK2过表达拟南芥中抗逆相关基因表达量的qRT-PCR检测Fig.9 qRT-PCR of the expression of stress-related genes in GmRPK2 overexpressed Arabidopsis
基因表达模式研究表明,GmRPK2在盐、干旱胁迫1 h时基因表达量明显上升而后则出现下降趋势。抗逆性检测结果表明,在盐胁迫、干旱胁迫和ABA处理下,GmRPK2过表达拟南芥在种子萌发期、幼苗生长阶段和成株时期均表现出抗逆性下降。
植物受到非生物胁迫处理后,往往会产生大量活性氧(ROS),进而引起氧化胁迫。植物体内过量的ROS会导致醛类物质的增多,从而破坏DNA、碳水化合物、脂膜及白质的结构。同时,过量的ROS也会造成质膜通透性增大,导致K+等电解质外渗[28]。植物对非生物胁迫的耐受性和低水平的ROS之间具有明显的正相关[29]。本研究用DAB组织染色法对盐胁迫前后拟南芥叶片中过氧化氢积累情况作了观察,结果显示,过表达植株叶片有显著增多的红褐色斑点积累,提示其过氧化氢的积累更多,叶片遭受的氧化胁迫更为严重。生理指标检测表明,盐处理后GmRPK2转基因拟南芥的MDA含量和电导率均高于野生型拟南芥,叶绿素含量下降更为明显,证明GmRPK2转基因拟南芥受到更加严重的氧化损伤,细胞质膜的损伤更为明显,这些可能都是GmRPK2转基因拟南芥耐盐性、耐旱性下降的内在生理原因。
另外,植物体的脯氨酸及可溶性糖是重要的渗透调节物质,植物在受到干旱等非生物胁迫时细胞内脯氨酸含量及可溶性糖含量会迅速增加,来维持细胞较高的渗透势,从而减少水分的散失[30]。干旱处理后,GmRPK2过表达植株的脯氨酸含量以及可溶性糖含量显著降低表明其在逆境下渗透调节能力要劣于野生型,这可能是其对干旱胁迫耐受性下降的原因之一。
已有研究表明,在拟南芥中存在一系列非生物胁迫信号的传导通路,如SOS、MAPK、CDPK、ABA信号通路等。这些信号通路相关的常见基因包括SOS3、ESK1、FRY1、SAD1、RD29B、ADH1和P5CS1等[31-34]。通过对拟南芥抗逆信号转导通路相关基因进行定量PCR,结果表明,SAD1、P5CS1、ADH13个基因的表达在GmRPK2过表达拟南芥中明显受抑。其中,SAD1位于CDPK信号通路,因此,GmRPK2可能通过抑制CDPK抗逆信号通路影响转基因植株的耐盐性,GmRPK2基因可能位于SAD1的上游。P5CS1则与植物的脯氨酸合成密切相关,GmRPK2的过表达造成了转基因拟南芥体内的脯氨酸合成受到抑制,从而导致转基因植株对干旱胁迫更为敏感[31]。ADH1是普遍存在于植物细胞中的一种正常条件下保持本底表达,逆境时表达活性提高的诱导酶。ADH1参与了多种植物激素如乙烯及ABA的传导途径,因此,其表达量与植物逆境也是密切相关。GmRPK2的过量表达明显抑制了ADH1的表达,这可能也是影响转基因拟南芥抗逆性的原因之一。
总之,经过初步研究,探明GmRPK2的过表达会造成拟南芥对非生物胁迫抗逆性的显著下降,初步揭示了这种抗逆性改变的内在生理机制和信号转导通路。对于GmRPK2影响植物抗逆性的其他内在机理,还需进一步研究证实。
猜你喜欢
亚热带植物科学(2022年1期)2022-05-17
天津医科大学学报(2021年3期)2021-07-21
世界科学技术-中医药现代化(2021年12期)2021-04-19
四川农业科技(2019年5期)2019-07-01
上海农业学报(2017年3期)2017-04-10
浙江柑橘(2016年1期)2016-03-11
中国医药生物技术(2015年4期)2015-12-26
红领巾·探索(2015年9期)2015-09-10
天然产物研究与开发(2014年6期)2014-04-27
植物营养与肥料学报(2014年1期)2014-03-11
