
乳酸菌拮抗食源性致病菌的研究及应用进展
2022-02-15 05:07康慎敏武瑞赟穆文强张世奇李平兰
食品科学 2022年1期
康慎敏,武瑞赟,穆文强,张世奇,2,李平兰,*
(1.中国农业大学食品科学与营养工程学院,北京 100083;2.北京科丽科技有限公司,北京 101300)
食源性致病菌是一类可以引起食物中毒或以食品为传播媒介的致病性细菌,常见的食源性致病菌有副溶血性弧菌(Vibrio parahaemolyticus)、沙门氏菌(Salmonella)、金黄色葡萄球菌(Staphylococcus aureus)和单核细胞增生李斯特菌(Listeria monocytogenes)(以下简称单增李斯特菌)等,由于其耐受性强、传播途径广、危害性大而在食品卫生方面存在较大的安全隐患[1-2]。致病菌感染人体后可能导致肠道炎症、慢性肾脏疾病、精神残疾、反应性关节炎和失明等疾病,甚至导致死亡。尽管卫生标准和生产规范不断提高,医疗和食品科学技术领域不断发展,但食源性致病菌引起的食源性疾病暴发仍然是影响机体健康的公共安全问题[3]。据世界卫生组织(World Health Organization,WHO)统计[4],全球每年感染食源性疾病人数高达10亿,并导致42万 人死亡,其中5 岁以下儿童处于特高风险,每年有超过12万 名儿童死于食源性疾病,因此,探寻一种有效的方法控制食源性致病菌具有重要的现实意义。
目前控制食源性致病菌的方法(巴氏杀菌、微波、辐照、电解水和等离子体等)虽然可以有效延长食品保质期,降低食品被致病菌污染的风险,但在实际应用中存在影响食品品质、安全争议大、自动化生产受限、能源消耗大等缺点,在食品的生产加工过程中已被部分限制使用[5]。生物抑菌法是指利用拮抗性微生物或其代谢产物,抑制或破坏食品中的病原体,相比其他抑菌方法,具有更安全、更温和、大众接受度更高的优势[6-7]。最近研究发现,乳酸菌对致病菌有一定的抑制能力,如罗伊氏乳杆菌可通过调节一氧化氮,有效地保护小鼠免于鼠伤寒沙门氏菌侵袭及肝脾扩散[8]。Frick等[9]研究表明双歧杆菌能够保护小鼠抵抗高致病性小肠结肠炎耶尔森氏菌的侵染。此外,乳酸菌可以与食源性致病菌竞争营养素(维生素、矿物质、微量元素和多肽)和能量[10],并可以产生一系列代谢产物(乳酸、丙酸、过氧化氢、抗菌肽和细菌素)抑制某些食源性致病菌的生长繁殖[11-13]。Schuenzel等[14]发现乳酸菌培养物可以延长蔬菜产品的保质期和降低产品被多种致病菌污染的风险,同时能减少化学防腐剂的添加量。因此越来越多的学者将乳酸菌应用于不同食品中以控制食源性致病菌。本文将介绍乳酸菌在食品中的应用基础,并主要从竞争作用、加强肠上皮屏障、干扰细菌毒力信号系统和产生抗菌代谢产物等方面说明乳酸菌拮抗致病菌的机理,同时综述乳酸菌在不同食品(乳、肉和果蔬)中抑制食源性致病菌的进展,最后对未来的研究方向进行展望。
1 乳酸菌拮抗致病菌在食品中的应用基础
乳酸菌是一类可在碳水化合物发酵过程中产生乳酸的革兰氏阳性细菌,其能产生有机酸、过氧化氢和细菌素等代谢产物,大量研究证明它是抑制食源性致病菌的天然有效菌株,并且乳酸菌在食品中已被广泛用作生产辅助剂、添加剂和生物防腐剂,具有良好的应用基础和前景[15-18]。
1.1 控制食源性致病菌的方法
随着现代食品工业的发展,人们对食品的安全、卫生和营养性的要求不断提高,如何在最大限度保持食品感官品质的前提下开发出安全且高效的控制食品中食源性致病菌的方法是现代食品科学研究的一个热点。目前,热灭菌技术(低温长时间、高温短时间、超高温、微波加热和欧姆加热)因低成本、高效益的优点,成为食品工业控制致病菌的主导技术。低温长时间灭菌技术是一种广泛用于乳业的杀菌工艺,这种杀菌方式能够杀灭牛乳中生长型致病菌,灭菌效果达到99.9%,但研究报道,耐热性致病菌如单增李斯特菌会由于在热环境压力下的提前暴露,而导致细菌存活率和热损伤恢复能力提高[19-20]。高温短时间灭菌技术和超高温灭菌技术可以杀灭致病菌,并灭活细菌繁殖体和孢子。然而较高的温度可能会导致产品中热敏营养成分损失,以及引起食品外观、颜色、风味和质地等感官特性发生变化[21]。微波加热对食品中鼠伤寒沙门氏菌、单增李斯特菌、大肠杆菌和金黄色葡萄球菌等致病菌具有良好的抑杀效果[22],例如,在肉鸡胴体表面接种单增李斯特菌(1.6×106CFU/mL),在2 450 MHz和900 W下微波60 s可实现表面完全灭菌[23]。然而研究表明,微波加热会对食品中营养成分(碳水化合物、蛋白质、维生素)造成一定影响[24]。欧姆加热能通过电流迅速且均匀地提高食品的温度,对果汁产品、牛奶和肉类中食源性致病菌的灭活有良好的作用效果,Sagong等[25]研究发现20 V/cm欧姆加热120 s能使橙汁和番茄汁中大肠杆菌O157:H7、鼠伤寒沙门氏菌和单增李斯特菌的数量降低5(lg(CFU/mL))以上。但欧姆加热不适合应用于非导电性物质(脂肪、油、空气、酒精和骨头)含量高的食物,具有一定的局限性[26]。
非热灭菌技术包括电解水、辐照和等离子体灭菌等。电解水作为一种生态友好、环保高效的新型消毒剂,对多种微生物(病毒、细菌和真菌)表现出广谱抗菌活性,并能在相对较短的时间内(通常5~20 s)杀灭食品、食品加工表面和非食品表面中的食源性致病菌,例如,研究表明酸性电解水(20 mg/mL有效氯质量浓度、pH 3.1、氧化还原电位1 150 mV)在20 ℃下处理2 min能有效灭活副溶血性弧菌、空肠弯曲杆菌、单增李斯特菌和嗜水气单胞菌[27],然而较低pH值的电解水具有腐蚀性,会影响某些食品的感官特性,并且电解水中的有效氯质量浓度会随着时间的推移而降低,从而导致灭菌效果下降[28]。此外,辐照灭菌法例如紫外辐照被证明能通过破坏核酸有效灭活微生物二聚体,并诱导微生物细胞膜损伤,对多种食源性致病菌具有显著杀菌效果[29],但同时较高剂量的辐照会影响食品的感官和功能特性[30]。等离子体可以通过损伤致病菌DNA来达到抑菌效果,Ulbin-Figlewicz等[31]研究发现氦等离子体分别处理2、5 min和10 min后小肠结肠炎耶尔森氏菌、大肠杆菌和荧光假单胞菌均显著减少了约2 个数量级,该技术虽环境友好但设备成本高昂,难以实现大范围的推广和应用。为了提高食品的安全性和功效性,生物抑菌正在逐渐替代物理和化学抑菌,基于生物源的抗菌剂主要包括益生菌及其代谢产物、噬菌体等[6]。
1.2 乳酸菌食品应用基础
乳酸菌的菌体和代谢物均能防止食源性致病菌的生长,提高食品的安全性和质量,已在乳制品、肉制品和果蔬等食品的生产加工中被广泛应用,食品中乳酸菌主要发挥3 种作用:一是作为生产辅助剂,在食品生产过程中起到发酵作用或者促进食品质地的形成,例如在搅拌型酸奶中添加干酪乳杆菌后,酸奶表现出更高的弹性指数和更低的固液平衡性,形成了更固态的凝胶结构,产品的内聚力和黏度显著增加,具有更好的细菌活力和贮藏稳定性[32];二是作为添加剂,能够增添食品风味、增强食品营养成分,Abadía-García等[33]发现添加干酪乳杆菌和鼠李糖乳杆菌(Lactobacillusrhamnosus)制成的奶酪中生物活性肽和游离氨基酸含量显著增加,从而增强了奶酪的抗菌、抗高血压、免疫调节和抗氧化等潜在的益生作用;三是作为生物防腐剂,例如在植物乳杆菌发酵制成的低盐香肠中,微生物群落多样性减少,香肠的pH值、酪胺、腐胺和尸胺的含量降低,乳杆菌的增殖在保证食品品质的同时延长了食品的保质期,显著抑制多种腐败菌(发光杆菌属、不动杆菌属、环丝菌属、嗜冷杆菌属和假单胞菌属)的生长[34]。此外,乳酸菌因其排斥竞争作用、微生物群体感应和抗菌代谢产物能在食品中充当生物源保护剂,防止食品被致病菌污染,抑制食品中致病菌生长[35-36]。
1.3 乳酸菌的抗菌机制
乳酸菌不仅具有增加食物的营养价值、延长食品保质期和促进健康等益处,还可以通过分泌黏蛋白增强自身黏附定植能力,从而竞争性地排除致病菌,有效地抑制致病菌生物膜的形成及对细胞表面的黏附作用(图1A、D、E)。此外,还可以通过产生抗菌物质、与致病菌争夺环境中有限的营养和生长因子等方面来抑制致病菌(图1B、C)[37]。

图1 乳酸菌抑制致病菌的作用机制[37]Fig.1 Mechanisms of action of probiotics[37]
1.3.1 竞争作用
乳酸菌能通过与致病菌竞争营养物质及能量来源,抑制致病菌的生长和增殖,Nadelman等[38]的研究表明干酪乳杆菌01(Lactobacillus paracasei01)能附着在牙齿表面形成生物膜,与变形链球菌和其他口腔致龋菌竞争营养素,从而降低口腔病原菌的致龋性,改善口腔的环境条件。蒙氏肠球菌(Enterococcus mundtii)与大肠杆菌NCTC 12900共培养30 h后,和单独培养相比,蒙氏肠球菌与碳水化合物代谢、细胞分裂、氨基酸代谢和能量产生及转化有关的蛋白质过表达,以此提高自身代谢速率和竞争营养底物的能力,从而抑制大肠杆菌的生长[39]。并且,乳酸菌产生的一些有机酸能够螯合微生物生长所需的金属离子,形成螯合物,从而抑制致病菌的生长[40]。此外,乳酸菌能阻断致病菌的定植位点,通过排斥竞争作用在该位置对致病菌产生物理屏障。Sribuathong等[41]研究发现,乳酸菌能通过分泌代谢产物,在肠道中与致病菌释放的分子竞争相同的受体,从而抑制致病菌对肠上皮细胞的黏附定植。Woo等[42]研究发现嗜酸乳杆菌KACC 12419、副干酪乳杆菌KACC 12427、干酪乳杆菌KACC 12413和鼠李糖乳杆菌KACC 11953(LactobacillusacidophilusKACC 12419、LactobacillusparacaseiKACC 12427、LactobacilluscaseiKACC 12413、LactobacillusrhamnosusKACC 11953)能通过竞争、排斥和置换作用,有效减少肠炎沙门氏菌亚种的生物膜形成。
1.3.2 加强肠屏障功能
致病菌可能会在食品加工的不同阶段污染食品,一旦致病菌通过食物进入宿主体内,它们会通过胃肠道分泌能够破坏肠道上皮细胞紧密连接的酶或毒素,从而破坏宿主组织的稳定。乳酸菌通过分泌能与肠上皮细胞表面糖基构成黏液层下层的黏蛋白,增加黏液层的厚度和空间位阻,从而抑制致病菌的黏附定植及毒素释放[43],例如Kim等[44]发现嗜酸乳杆菌A4(Lactobacillus acidophilusA4)能使细胞中黏蛋白-2(mucin-2,MUC-2)的表达增加,显著抑制了大肠杆菌O157:H7对HT-29肠上皮细胞的黏附。此外,乳酸菌能通过提高与肠上皮细胞紧密连接相关的复合体蛋白(紧密连接蛋白、闭合蛋白、胞质衔接蛋白和连接黏连分子)表达量来增强上皮屏障的完整性[45]。Johnson-Henry等[46]研究发现鼠李糖乳杆菌对大肠杆菌O157:H7诱导的犬肾小管上皮细胞(madin-darby canine kidney-1,MDCK-1)和人结肠腺癌肺转移细胞(T84)组织培养物中闭锁小带蛋白-1(zonula occludens-1,ZO-1)和闭合蛋白-1(Claudin-1)的破坏具有保护作用。Bhat等[47]发现发酵乳杆菌(Lactobacillus fermentumMTCC-5898)能够通过维持ZO-1和Claudin-1蛋白的分布来限制大肠杆菌在Caco-2细胞中的黏附和侵袭。
1.3.3 干扰毒力信号系统
细菌在生长的过程中会分泌信号因子,在周围环境中通过与受体结合抑制或激活特异性基因的表达,从而调控相关的生物学功能,这种现象称为群体感应。致病菌借助群体感应机制,不仅能与周围环境相互交流,还能通过称为“自我诱导剂”的分子对化学物质进行信号传导,从而调节毒力的表达。鼠伤寒沙门氏菌和大肠杆菌O157:H7具有特定的毒力遗传成分,如致病岛和毒力质粒。这两种微生物都具有III型分泌系统,该系统负责将效应蛋白转移并注入宿主细胞,以帮助其黏附、连接和细胞侵袭。有报道表明乳酸菌产生的生物活性分子会抑制大肠杆菌O157:H7[48]和沙门氏菌[49]中毒力基因的表达,且乳酸菌如乳酸杆菌和双歧杆菌能通过产生酶、分泌自诱导拮抗剂等方式抑制致病菌的定植或毒性相关基因转录中的信号传导,从而降解自诱导分子[50]。
1.3.4 产生抗菌类代谢物质
1.3.4.1 细菌素
细菌素是核糖体合成的长度为20~60 个氨基酸的分泌型抗菌肽(antimicrobial peptide,AMP),它能够抑制革兰氏阴性和革兰氏阳性致病菌[51]。细菌素主要分为I类(羊毛硫抗生素)、II类(热稳定且无修饰的小分子肽)、III类(热不稳定的大分子肽)和IV类(蛋白质化合物)[52]。其中,乳酸菌产生的乳酸链球菌素(nisin)是美国食品药品管理局(Food and Drug Administration,FDA)认证为安全的物质,已被发现能够应用在牛奶、火腿和鲑鱼等食品中抑制革兰氏阳性致病菌[53]。弯曲乳杆菌32Y(Lactobacilluscurvatus32Y)产生的细菌素能够降低猪肉、牛排、碎牛肉和法兰克福香肠中的单增李斯特菌数量[54]。Liu Guorong等[55]研究发现中国传统发酵宣威火腿中分离出的戊糖乳杆菌31-1(Lactobacillus pentosus31-1)产生的细菌素显示出热稳定性,并具有良好的抗李斯特菌活性。鼠李糖乳杆菌1.0320(Lactobacillus rhamnosus1.0320)产生的新型细菌素显示出对致病菌的广谱抗菌活性[56]。细菌素的抗菌机制主要是由于其能引起细胞质膜的构象变化,在细胞膜表面形成孔洞,破坏细胞膜通透性,导致质子动力(protonmotive force,PMF)耗散和细胞内离子及分子泄漏(图2I和II),从而达到抑菌效果。Wang Yao等[57]研究表明植物乳杆菌LPL-1(LactobacillusplantarumLPL-1)产生新型IIa类细菌素能够通过静电相互作用积聚在单核细胞增生李斯特菌的细胞膜上,并通过疏水相互作用引起孔形成,从而导致胞内物质(ATP、无机磷酸盐、K+、蛋白质和核酸)泄漏以及质子动力的损失。

图2 细菌素抑菌机理[58]Fig.2 Antibacterial mechanism of bacteriocin[58]
1.3.4.2 有机酸类
乳酸菌通过碳水化合物代谢发酵产生乳酸、乙酸和丙酸等有机酸,导致了酸性环境的形成,如图3所示,当细菌细胞质的pH值高于周围环境的pH值时,环境中未解离的有机酸会通过微生物膜扩散进入细胞内解离出酸性阴离子,使细胞质的pH值降低,从而影响细菌的酶活性、结构蛋白合成和DNA/RNA功能[59-60],而且离解出的酸性阴离子积累会阻碍质子原动力并抑制微生物重新碱化细胞质。Harris等[61]发现在牛肉条表面喷洒2%和4%的乳酸可分别导致大肠杆菌O157:H7和沙门氏菌数量减少约2.0(lg(CFU/mL))和1.5(lg(CFU/mL))。Al-Rousan等[62]发现用体积分数0.4%乙酸分别处理沙门氏菌和金黄色葡萄球菌5 d和7 d后,细菌数量能降低至检出限以下。乳酸菌产生的各种有机酸已被用作致病菌抑制剂,且有研究表明这些有机酸在抑制致病菌方面存在协同作用[62]。
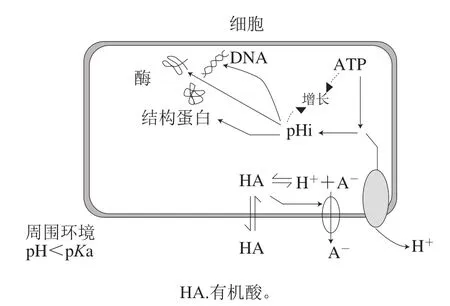
图3 有机酸抑菌机理[60]Fig.3 Antibacterial mechanism of organic acids[60]
1.3.4.3 胞外多糖
胞外多糖是乳酸菌通过非共价相互作用松散结合到细胞上或释放到周围环境中的一种代谢产物,具有抗肿瘤、抗氧化、免疫调节等功能特性。Nehal等[63]发现乳酸乳球菌F-mou菌株(Lactococcus lactisF-mou)产生的胞外多糖对9 种致病菌(金黄色葡萄球菌、大肠杆菌、铜绿假单胞菌、单增李斯特菌和蜡样芽孢杆菌等)均表现出抑制作用,其中对革兰氏阳性菌具有显著的抗菌活性。此外,乳酸菌产生的胞外多糖能够抑制致病菌的生物膜形成,这是由于胞外多糖起到了空间排阻的作用,通过屏蔽表面黏附位点,抑制了致病菌的黏附[64]。Merghni等[65]发现干酪乳杆菌产生的胞外多糖对金黄色葡萄球菌具有抗黏附和抗生物膜活性,Wang Ji等[66]研究发现植物乳杆菌YW32(LactobacillusplantarumYW32)产生的胞外多糖能够抑制大肠杆菌O157、志贺氏菌CMCC、金黄色葡萄球菌AC1和鼠伤寒沙门氏菌S50333的生物膜形成,且呈现浓度依赖性。嗜酸乳杆菌A4产生的1 mg/mL胞外多糖能够使大肠杆菌O157:H7生物膜形成量减少87%,这可能是胞外多糖通过部分影响细菌表面特性(减弱细胞表面修饰或减少细胞-细胞表面相互作用)来抑制病原菌的初始附着和自聚集所致[67]。
2 乳酸菌在食品中对致病菌的抗菌作用
2.1 奶及奶制品
奶制品(如奶酪)因低盐、高蛋白质和脂肪含量的理化特性,为食源性致病菌的生存提供了有利条件。而发酵乳和其他奶制品相比,因其低pH值、低贮藏温度及可能含有发酵微生物产生的抑菌化合物等因素具有更好的安全性,但某些致病菌如单增李斯特菌、沙门氏菌和大肠杆菌O157:H7仍可在发酵乳中生存或生长。近些年的研究发现多种乳酸菌不仅可以抑制奶制品中致病菌的生长,对奶制品的理化特性无负面影响,还能起到增强食品营养性、赋予奶制品特殊风味的作用。
2.1.1 奶酪
奶酪是生牛乳在发酵剂和凝乳酶的作用下,经过凝固、压制、后熟等过程得到的乳制品。鼠李糖乳杆菌EM1107(Lactobacillus rhamnosusEM1107)有助于乳制品理想风味和质地的形成,它产生的乳酸可以和乳制品中其他物质形成中间体,产生风味化合物[68]。Rolim等[69]在半硬质山羊奶酪上接种鼠李糖乳杆菌EM1107后,继续接种金黄色葡萄球菌,发现鼠李糖乳杆菌EM1107在奶酪模拟消化后仍能存活(6.75(lg(CFU/g))),且在奶酪贮存的7、14 d和21 d时分别抑制了1.55%、1.70%和21.66%的金黄色葡萄球菌,他们推测,这可能与鼠李糖乳酸菌在发酵代谢过程中产生的各种化合物(有机酸、抗菌肽和活性蛋白)有关。类似地,Prezzi等[70]将鼠李糖乳杆菌和单增李斯特菌共同接种到米纳斯鲜奶酪中,发现鼠李糖乳杆菌对米纳斯鲜奶酪的pH值、水分、脂肪、蛋白质和质地未产生明显影响,并且鼠李糖乳杆菌在奶酪模拟胃肠道消化后仍使李斯特菌的存活率下降了20%,这说明鼠李糖乳杆菌能在宿主体内通过竞争细胞受体、分泌抗氧化剂或影响肠细菌病原体定植相关基因表达来降低病原体的侵袭性。同样为了抑制单增李斯特菌,Abadía-García等[33]将干酪乳杆菌和鼠李糖乳杆菌加入含单增李斯特菌的干酪中发现,在贮存20 d后李斯特菌数量减少了约1(lg(CFU/g)),且乳酸菌的加入使干酪中生物活性肽增多,增加了干酪的营养性。
2.1.2 发酵乳
发酵乳因其营养丰富、风味独特的特点备受大众青睐,乳酸菌通过发酵作用生成酯、醛等芳香化合物,同时还可生成乳酸、细菌素等抑菌成分,提高发酵乳的安全性,延长产品的保质期。研究表明在利用植物乳杆菌Tennozu-SU2(LactobacillusplantarumTennozu-SU2)和乳酸乳球菌BF1(LactobacilluslactisBF1)发酵豆乳的过程中接种6(lg(CFU/mL))的沙门氏菌,发酵2 d后沙门氏菌数量下降了1 个数量级,而对照组中的沙门氏菌数量却增加了2 个数量级[71],可见乳酸菌在豆奶发酵过程中不仅可以作为一种发酵剂,还能降低豆奶在发酵过程中被致病菌污染的风险。Haraguchi等[71]在植物乳杆菌Tennozu-SU2和乳酸乳球菌BF1发酵制得的发酵豆奶中接种致病菌,10 ℃下贮存7 d后单增李斯特菌和鼠伤寒沙门氏菌的数量均减少了约3(lg(CFU/mL)),研究分析这与益生菌发酵剂的抗菌化合物、酸化力和氧化还原活性有关。Kamal等[72]发现添加鼠李糖乳杆菌发酵制成的酸奶和普通酸奶相比,在接种高剂量致病菌(6(lg(CFU/mL)))8 h后,乳杆菌发酵组中的大肠杆菌O157:H7、金黄色葡萄球菌和小肠结肠炎耶尔森氏菌菌量分别下降了3、1、2(lg(CFU/mL)),有明显的抑制效果,由此可见乳酸菌制成的发酵乳产品在存储消费的过程中具有更长的货架期和更好的安全性。
2.2 肉及肉制品
肉类最初的微生物区系由多种微生物组成,但贮藏温度和包装是影响肉类和肉制品微生物生态和质量稳定性的重要因素,肉制品的贮藏方式一般有冷藏(4 ℃)、冰藏(0 ℃)和冰冻(-20 ℃)3 种。由于嗜冷致病菌如单增李斯特菌等的存在,如果受污染的食物在冷藏(4 ℃)条件下长时间保存,致病菌很容易生长到感染水平。近些年来的研究发现乳酸菌可有效抑制新鲜肉、熟肉和真空包装肉制品中的致病菌,并且还能赋予肉制品独特风味。
2.2.1 普通包装肉制品
普通包装无法达到控制微生物的目的,且微生物种类复杂。但是因为操作流程的简便,应用较为广泛,并适合家庭贮藏使用。近些年,人们在追求快节奏生活方式的同时,也开始注意食品的健康和安全问题,由此,控制普通包装食品贮藏中致病菌的研究应运而生。如发酵乳杆菌ACADC179(Lactobacillus fermentumACA-DC179)被证明能应用于生鸡肉,有效减少肠炎沙门氏菌和单增李斯特菌的生长,并对生鸡肉没有不利影响[73]。Koo等[74]将接种有混合乳酸菌(Lactobacillus animalisLa51、Lactobacillus amylovorusM35和Pediococcus acidilacticiD3)与单增李斯特菌的法兰克福香肠置于4 ℃冷藏8 周后发现,接种乳酸菌会降低李斯特菌的生长幅度,但乳酸菌的数量在发酵过程中未出现较大的变化。除了乳酸菌和致病菌共培养产生的抑制作用,植物乳酸杆菌NRRL B-4496(Lactobacillus plantarumNRRL B-4496)的无细胞上清液也可以抑制新鲜牛肉中的大肠杆菌、金黄色葡萄球菌、鼠伤寒沙门氏菌和单增李斯特菌,提高牛肉的安全性和质量,且熟肉的感官肉色不会受到影响,通过pH值中和实验及上清液组成分析实验,证明上清液抗菌活性可以归因于有机酸(主要是乙酸和乳酸)[13]。
此外,尽管发酵肉制品因多种因素(低pH值、低水分活度、亚硝酸盐、化学防腐剂和天然微生物区系)导致产品的卫生风险普遍较低,但某些病原微生物(例如单增李斯特菌)可以适应发酵的环境并破坏食品的安全性[75]。乳酸菌作为发酵剂不仅有助于提高发酵肉制品的质量,还能抑制肉制品中致病菌的生长。在接种有4.22(lg(CFU/g))单增李斯特菌的香肠中添加弯曲乳酸杆菌(Lactobacillus curvatus54M16)作为发酵剂,3 d内单增李斯特菌数量减少至3.05(lg(CFU/g)),而未添加乳酸菌发酵的香肠中单增李斯特菌菌量增加至5.72(lg(CFU/g)),同时,接种乳酸菌发酵的香肠中与腐败相关的细菌(环丝菌、嗜冷杆菌、假单胞菌等)数量显著降低[76]。
2.2.2 真空包装肉制品
当生肉和肉制品在真空或富含CO2的环境中冷藏包装时,占优势的微生物区系是乳酸菌,将其用作保护剂可提高食品的安全性[77]。清酒乳杆菌TH1(Lactobacillus sakeiTH1)不会影响熟火腿和法式香肠的感官特性,且在4 ℃和8 ℃下对真空包装的香肠中的单增李斯特菌具有抑制作用,这与清酒乳杆菌在微营养环境下的强竞争力、pH值的降低和未解离乳酸的抑菌作用有关[78]。类似地,清酒乳杆菌被证明能抑制真空包装的切片熟火腿中的单增李斯特菌和大肠杆菌O157:H7的生长,且不会影响火腿的感官品质[79]。肠系膜明串珠菌L124(Leuconostoc mesenteroidesL124)和弯曲乳杆菌L442(Lactobacillus curvatusL442)是一类可以产生细菌素的菌株,产生的细菌素能与低pH值环境和气体组成环境等因素协同抑制致病菌[80-81],目前已有研究将其用在真空包装的4 ℃的熟腌猪肩肉中,30 d内可以使李斯特菌种群减少约1.5(lg(CFU/mL))[82]。此外,多物种实验的种间竞争模型可以更真实地反映食品中微生物群落的动态,Ye Keping等[83]使用詹姆森效应模型拟合单增李斯特菌和乳酸杆菌在真空包装冷藏猪肉中混合培养的生长曲线,发现混合培养中估计的单增李斯特菌的最大种群密度(3.88(lg(CFU/mL)))远低于单增李斯特菌纯培养物中的估计密度(6.01(lg(CFU/mL))),且当乳酸杆菌生长到静止期时,单增李斯特菌也停止生长。
2.3 果蔬
2.3.1 新鲜果蔬
新鲜果蔬因其所含的营养是人类饮食中的重要物质而受到广大消费者的喜爱,但是由于缺乏食用前的烹饪过程,当诸如沙门氏菌、单增李斯特菌等致病菌污染或定植于果蔬后,可能导致人群中食源性疾病暴发。而常用的添加化学防腐剂或植物源天然防腐剂的杀菌方法会破坏果蔬的感官和营养特性。相比之下,将乳酸菌喷涂在果蔬表面的方法,既能对致病菌起到一定的抑制作用,又能不破坏果蔬品质和营养,深受研究者青睐。Alegre等[84]以6(lg(CFU/g))的鼠李糖乳杆菌添加到楔形苹果上,在5 ℃和10 ℃下14 d的贮藏期间使单增李斯特菌数量减少了1(lg(CFU/g)),而对苹果品质没有任何影响。Dong Qingli等[85]研究发现植物乳杆菌可以通过抑制贮藏过程中李斯特菌毒力基因的表达,从而抑制卷心菜中李斯特菌的生长。此外,明串珠菌(LeuconostocCM160、LeuconostocCM135、LeuconostocPM249)也被应用于金冠苹果和卷心莴苣切叶中控制食源性致病菌,结果发现明串珠菌不会促进苹果伤口或菜叶中的变质反应(褐变、改变果胶活性或产生异味),并能使6(lg(CFU/g))的单增李斯特菌菌量降低至检出限以下,鼠伤寒沙门氏菌和大肠杆菌的菌量减少1~2(lg(CFU/g))[86]。
2.3.2 泡菜
泡菜是传统的由调味蔬菜发酵而来的食品,具有助消化、预防癌症等有益特性,乳酸菌是泡菜中的主要微生物,对泡菜风味的形成具有重要作用。Kim等[87]研究发现泡菜中分离出的短乳杆菌DF01(Lactobacillus brevisDF01)和乳酸片球菌K10(Pediococcus acidilacticiK10)能够降低鼠伤寒沙门氏菌和大肠杆菌O157:H7对肠上皮细胞的黏附能力。类似地,从泡菜中分离出营养改良的植物乳杆菌DGK-17(Lactobacillus plantarumDGK-17)显示出对铜绿假单胞菌和肺炎克雷伯菌的抗微生物活性[88]。
3 结 语
近年来,虽然食品原料、生产加工和贮存物流等环节的质量管理技术不断革新且成效良好,但食源性致病菌的控制仍是全球关注的问题,与其他控制方法相比,合理使用微生物控制的方法是符合食品安全、具有开发前景的一种技术手段,其中由于乳酸菌具有种类丰富和生理功效强的优势,并且能够改善食品风味、延长食品保质期,在食品领域具有良好的应用前景,受到了研究者的青睐,但是乳酸菌抑制食源性致病菌的机制研究多集中在乳酸菌的代谢产物抑菌方面,关于抗菌作用的分子模式解析较少。此外,现阶段大量文献报道了乳酸菌在乳、肉和果蔬等制品中对食源性致病菌的抑制效果,但对具有抗菌活性的乳酸菌添加到各类食品中后对食品的感官品质的影响及添加到各类食品中合适的剂量与组合仍缺乏系统研究,而这是具有抗菌活性的乳酸菌作为益生商用生物抑菌剂实际应用于食品加工生产中必须考虑的问题。
猜你喜欢
动物医学进展(2022年9期)2022-09-06
中老年保健(2022年1期)2022-08-17
中国食品学报(2022年6期)2022-07-19
中国饲料(2021年17期)2021-11-02
食品安全导刊(2021年21期)2021-08-30
食品安全导刊(2021年20期)2021-08-30
食品安全导刊(2021年20期)2021-08-30
中老年保健(2021年6期)2021-08-24
小小说月刊(2013年6期)2013-05-14
琴童(2009年2期)2009-02-26

- 食品科学的其它文章
- 花中多糖化学组成与生物活性研究进展
- 番茄碱的制备及其对乙酰胆碱酯酶的抑制作用
- 色氨酸抑制体外模型中晚期糖基化终末产物形成机理
- 松油烯-4-醇对荧光假单胞菌抑菌能力及作用机理
- Widely Targeted Metabolomic and Transcriptomic Analyses Reveal the Regulation of Trypsin on Antioxidant Enzymes and Saturated Fatty Acids in Hylocereus undatus Fruit during Storage
- 乙烯利处理‘赤霞珠’葡萄果实对其葡萄酒中酚类物质组分的影响