
松油烯-4-醇对荧光假单胞菌抑菌能力及作用机理
2022-02-15 05:06耿一鸣李婷婷励建荣申照华郭晓华劳敏军周小敏于建洋
食品科学 2022年1期
耿一鸣,李婷婷*,励建荣,*,谢 晶,林 洪,王 轰,申照华,郭晓华,劳敏军,周小敏,于建洋
(1.渤海大学食品科学与工程学院,生鲜农产品贮藏加工及安全控制技术国家地方联合工程研究中心,辽宁 锦州 121013;2.大连民族大学生命科学学院,辽宁 大连 116600;3.上海海洋大学食品学院,上海 201306;4.中国海洋大学食品科学与工程学院,山东 青岛 266100;5.蓬莱京鲁渔业有限公司,山东 烟台 265600;6.山东美佳集团有限公司,山东 日照 276800;7.浙江兴业集团有限公司,浙江 舟山 316120;8.泰祥集团荣城泰祥食品股份有限公司,农业农村部冷冻调理海洋食品加工重点实验室,山东 威海 264309)
微生物是导致食品腐败的主要因素。微生物在生长的过程中,会分解破坏食品中的碳水化合物、蛋白质、脂质等营养成分,并产生对人体有害的物质[1]。嗜冷菌是一类菌的总称,其具有独特的嗜冷机制,能在极寒的条件下生存繁殖。荧光假单胞菌为假单胞菌属嗜冷革兰氏阴性菌,是最常见的嗜冷菌之一[2-3]。在冷藏及有氧的条件下,荧光假单胞菌会产生蛋白酶、脂酶等,破坏食品中的营养物质,导致食品腐败[4]。荧光假单胞菌是水产品、蔬菜、肉制品、乳制品的优势腐败菌[5-6]。
目前,化学防腐剂已广泛应用于食品行业,其可以高效地防止食品被污染,进而延长其保质期。然而,长期使用化学防腐剂会增加细菌的抗药性,导致一些潜在的副作用[7]。天然防腐剂具有无毒无害、抗菌能力强、抗菌谱广、能够提升风味等特点,因此,寻找替代化学防腐剂的天然防腐剂现已成为近年来研究热点[8]。
精油是一类从具有芳香味植物原料中提取的芳香油性液体,由于其具有广谱抗菌性,被认为是天然安全的防腐剂和抗菌剂[9]。茶树精油是从互叶白千层的枝叶中提取出来的一种具有薄荷醇气味的无色至淡黄色液体,具有良好的杀菌、杀虫、防腐等特性,在香料、食品工业、化妆品中广为应用[10-14]。松油烯-4-醇又名4-松油烯醇、4-松油醇、萜品烯,是茶树精油的主要有效成分之一,也是多数植物精油所含的单萜类成分,具有抗炎、抗肿瘤等作用,含有胡椒香、陈腐木材的香气,是用于配制香辛类香精的食品用香料[15-16]。近年来,消费者愈来愈重视食品的品质,在运输及贮藏过程中,使用天然防腐剂取代人工合成的化学防腐剂已成重点研究方向。前人对松油烯-4-醇的研究多集中于抗肿瘤抗疾病,对抑菌活性及机理报道尚少[17]。基于此,本实验通过测定最小抑菌浓度(minimum inhibitory concentration,MIC)并绘制抑菌生长曲线,评价松油烯-4-醇对食品中常见的致腐菌-荧光假单胞菌的抑菌能力;并通过测定电导率评价了其对细胞膜通透性的影响;同时,利用扫描电子显微镜(scanning electron microscope,SEM)、二乙酸荧光素(di-O-acetylfluorescein,FDA)染色实验评价了其对细胞膜完整性的影响;此外,测定胞外总蛋白含量、DNA含量、胞内ATP酶活性、碱性磷酸酶活性,以进一步阐明其抑菌机理。旨在为松油烯-4-醇在食品保鲜领域的开发应用提供理论依据。
1 材料与方法
1.1 菌种与试剂
荧光假单胞菌(Pseudomonas fluorescens)由辽宁省渤海大学食品安全重点实验室保藏,于28 ℃、160 r/min摇床中培养。
松油烯-4-醇(纯度98%) 上海迈瑞尔化学技术有限公司;LB肉汤培养基、LB营养琼脂 青岛高科园海博生物技术有限公司;卡那霉素 国药集团化学试剂有限公司;氯化钠 天津市优谱化学试剂有限公司;锌片上海迈砷化工有限公司;Ezup柱式细菌基因组DNA抽提试剂盒DNA Maker、琼脂糖 上海生工生物工程股份有限公司;50×TAE缓冲液、磷酸盐缓冲液(phosphate buffered saline,PBS) 北京索莱宝生物科技有限公司;超微量ATP酶(Na+、K+)测试盒、碱性磷酸酶(alkaline phosphatase,AKP)测试盒、总蛋白定量测定试剂盒 南京建成生物工程研究所。
1.2 仪器与设备
SW-CJ-2FD超净工作台 苏景集团苏州安泰技术有限公司;立式压力蒸汽灭菌锅 上海申安医疗器械厂;LRH系列生化培养箱 上海一恒科技有限公司;MS105UD电子分析天平、FE30电导率仪 瑞士梅特勒-托利多仪器有限公司;PE Victor X3多功能酶标仪美国PerkinElmer公司;Biofuge Stratos冷冻高速离心机、Sorvall Legend Micro21R台式微量离心机 美国Thermo Fisher公司;SS-4800场发射SEM、F7000荧光分光光度计、E-1045镀金仪 日本日立公司;GelDoc XR+全自动凝胶成像系统 美国Bio-Rad公司;UV-2550紫外-可见分光光度计 日本岛津公司;SCIENTZ-IID超声波细胞粉碎机 宁波新芝生物科技股份有限公司。
1.3 方法
1.3.1 松油烯-4-醇对荧光假单胞菌MIC的确定及抑菌曲线绘制
参考Yang等[18]的方法略作修改,测定松油烯-4-醇的最小抑菌剂量(minimum inhibitory concentration,MIC)。吸取75 μL培养至对数生长期(107CFU/mL)的荧光假单胞菌菌液于96 孔细胞培养板中。再将溶于无菌去离子水的松油烯-4-醇加入到上述孔中,使松油烯-4-醇的终剂量分别为4、2、1、0.5、0.25 μL/mL。以75 μL质量浓度为50 μg/mL的卡那霉素为阳性对照,以等体积的无菌去离子水为阴性对照。将96 孔细胞培养板放置于28 ℃生化培养箱中培养12 h,使用酶标仪测定OD595nm。将测定的结果进行比较,选择与阳性对照无明显差异的浓度作为松油烯-4-醇对荧光假单胞菌的MIC,每个样品进行3 次平行实验。
吸取200 μL培养至对数生长期(107CFU/mL)的荧光假单胞菌菌液于10 mL LB肉汤培养基中,加入松油烯-4-醇使其终剂量分别为1、2、4 MIC,以无菌去离子水作为空白对照。在28 ℃生化培养箱中培养,分别于0、2、4、6、8、10、12、24 h用酶标仪测定OD595nm。
1.3.2 碱性磷酸酶活力的测定
参考舒慧珍等[19]的方法并略作修改。将培养至对数生长期(107CFU/mL)的荧光假单胞菌菌液于4 ℃下8 000 r/min离心10 min,弃上清,菌体重悬于无菌PBS(pH 7.0)中,重复离心洗涤3 次,最终重悬于等体积PBS中。加入松油烯-4-醇使其终剂量分别为1、2、4 MIC,并以无菌去离子水作空白对照,于摇床中培养4 h。离心弃上清液收集沉淀菌体,加入PBS重悬,用超声破碎仪在300 W冰水浴中破碎菌体,每次超声时间为5 s,间隔10 s,重复4 次。采用碱性磷酸酶(alkaline phosphatase,AKP)试剂盒测定细胞内的碱性磷酸酶活力。
1.3.3 荧光假单胞菌胞内电导率的测定
取培养至对数生长期(107CFU/mL)的荧光假单胞菌液,于8 000 r/min离心10 min,吸出上清液,留菌体,用PBS重悬菌体后,以8 000 r/min离心10 min,重复两次,最终重悬于PBS。加入溶于无菌去离子水的松油烯-4-醇,使其终剂量分别为1、2 MIC,以无菌去离子水作空白对照。于摇床中培养12 h,每2 h取样测其电导率。
1.3.4 荧光假单胞菌细胞膜通透性的测定
根据舒慧珍等[8]的FDA染色方法并略作修改。取培养至对数生长期的荧光假单胞菌菌液(107CFU/mL),离心去上清液并用PBS重悬菌体,加入松油烯-4-醇使其终剂量为1、2、4 MIC,以无菌去离子水作空白对照,于摇床培养。分别取培养2、5、8 h的菌悬液6 mL于8 000 r/min离心10 min,吸出上清液,所得到的菌体用PBS重悬3 次,离心后吸出上清液,加入250 μL溶于丙酮的FDA(2 mg/mL),在常温下静置20 min,PBS重悬洗涤3 次,离心弃上清液重悬于6 mL的PBS。使用荧光分光光度计测定平均荧光强度。荧光分光光度计激发波长297 nm,发射波长527 nm,激发狭缝与发射狭缝5 nm。
1.3.5 扫描电子显微镜观察松油烯-4-醇处理后荧光假单胞菌形态
取培养至对数生长期(107CFU/mL)的荧光假单胞菌菌液,加入终剂量为1/2、1、2、4 MIC的松油烯-4-醇,以无菌去离子水做空白对照,于摇床中培养6 h。将培养好的菌液放置于离心管中以8 000 r/min离心10 min,吸出上清液,所得到的菌体用PBS重悬洗涤3 次后,每次再以8 000 r/min离心10 min。加入体积分数2.5%的戊二醛,将5 mm×5 mm×1 mm无菌锌片放入固定3 h,最后依次用30%、50%、70%、100%的无水乙醇溶液对硅片梯度洗脱,每次30 min。锌片表面喷金后用扫描电镜观察表面荧光假单胞菌细胞的形态。
1.3.6 荧光假单胞菌细胞内DNA含量的测定
将荧光假单胞菌培养至对数生长期(107CFU/mL),离心吸出上清液,重悬于PBS中,重复离心重悬洗涤3 次。加入松油烯-4-醇,使其终剂量为0.5、1、2、3、4 MIC,并以无菌去离子水作空白对照,于28 ℃摇床培养3 h。采用Ezup柱式细菌基因组DNA抽提试剂盒提取荧光假单胞菌的基因组DNA,进行DNA琼脂糖凝胶电泳。电压为90 V,电流为110 mA,电泳时长为30 min,电泳完成后,将凝胶放置于全自动凝胶成像装置,拍照并分析结果。
1.3.7 荧光假单胞菌细胞内总蛋白质量浓度的测定
将荧光假单胞菌培养至对数生长期(107CFU/mL),弃上清液留下层菌体,加入0.85%无菌生理盐水重复离心重悬洗涤两次,再加入松油烯-4-醇使其终剂量为1、2 MIC,以无菌去离子水作空白对照,于摇床中培养。分别取0、3、6、9、12 h的菌悬液离心去上清液留菌体,加入生理盐水,用超声破碎仪在300 W冰水浴中破碎菌体,每次超声时间为5 s,间隔10 s,重复4 次并于4 ℃保存。采用总蛋白定量测定试剂盒得到各组样品的蛋白质量浓度,根据结果分析松油烯-4-醇对荧光假单胞菌细胞内总蛋白质量浓度的影响。
1.3.8 荧光假单胞菌细胞内超微量Na+、K+-ATP酶活力的测定
取培养至对数生长期(107CFU/mL)的荧光假单胞菌菌液,弃上清液并于菌体中加入生理盐水,重悬两次后置于生理盐水中,加入终剂量为1、2 MIC的松油烯-4-醇,以无菌去离子水作空白对照,与摇床中培养。取0、3、6、9、12 h菌悬液离心加入生理盐水重悬3 次,用超声破碎仪破碎细胞。采用超微量Na+、K+-ATP酶试剂盒测定荧光假单胞菌细胞内Na+、K+-ATP酶活力。
1.4 数据处理与分析
所有实验重复3 次,结果以平均值±标准差表示。使用Excel软件进行数据分析,运用Origin 8.5软件作图。
2 结果与分析
2.1 松油烯-4-醇的MIC及其对荧光假单胞菌时间抑菌曲线
如图1所知,不同终剂量的松油烯-4-醇对荧光假单胞菌均有抑制作用,终剂量为0.25、0.5、1 μL/mL的松油烯-4-醇对荧光假单胞菌具有轻微的抑制生长作用。在培养至对数生长期(107CFU/mL)的荧光假单胞菌中加入终剂量为2 μL/mL的松油烯-4-醇培养12 h后,其菌液密度低于等体积终质量浓度为50 μg/mL卡那霉素的阳性对照,表明松油烯-4-醇对荧光假单胞菌的MIC为2 μL/mL。与Kang Shimo等[20]研究的抑菌剂乳糖醛酸相比,松油烯-4-醇对荧光假单胞菌具有较好的抑菌效果。

图1 不同剂量的松油烯-4-醇对荧光假单胞菌抑菌效果Fig.1 Antibacterial effect of terpinene-4-ol at different concentrations on Pseudomonas fluorescens
由图2可知,加入1、2、4 MIC的松油烯-4-醇在培养2 h后对荧光假单胞菌的生长产生了明显的抑制作用,与空白对照相比差异较大。随着培养时间的延长,松油烯-4-醇对荧光假单胞菌的抑菌能力没有变化,表明松油烯-4-醇有较强的抑菌能力,并具有良好的抑菌稳定性。
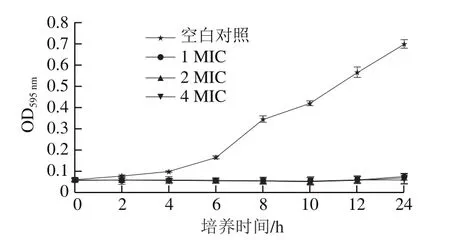
图2 不同剂量的松油烯-4-醇对荧光假单胞菌的时间抑菌曲线Fig.2 Time-inhibition curves of terpinene-4-ol at different concentrations on Pseudomonas fluorescens
2.2 松油烯-4-醇对荧光假单胞菌细胞壁的影响
AKP是一种大量存在于微生物中的磷酸水解酶,其在磷酸盐的代谢中起着无可替代的作用[21]。AKP存在于细胞壁与细胞膜之间,当细胞壁或细胞膜的完整性遭到损伤,AKP便会泄露到菌液中,使菌体内AKP活力降低。本实验通过测定松油烯-4-醇对AKP活力的影响,以反映其对荧光假单胞菌细胞壁的损伤情况。如图3所示,经4 h松油烯-4-醇培养后,空白对照组AKP活力明显增加;1、2、4 MIC处理组AKP活力下降明显。结果表明,0~4 h培养后的空白对照组菌体生长,使其胞内AKP活力增加;处理组AKP活力随松油烯-4-醇剂量的增加及培养时间的延长而降低,表明松油烯-4-醇能有效地破坏荧光假单胞菌的细胞壁并且可能改变细胞膜的通透性,使胞内AKP活性降低。这与柏梅[22]研究发现山苍子精油能够破坏金黄色葡萄球菌菌体的细胞壁及细胞膜,使胞内AKP活力被抑制的实验结果相同。
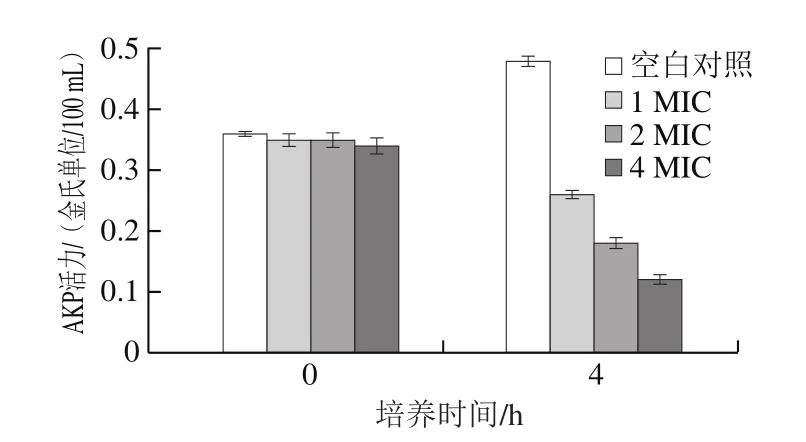
图3 松油烯-4-醇对AKP活力的影响Fig.3 Effect of terpinene-4-ol on AKP activity
2.3 松油烯-4-醇对荧光假单胞菌内电导率的影响
当抑菌剂作用于菌体后,会使菌体细胞膜的通透性增加,导致胞内的钾离子、钙离子、氢离子等小离子从膜内外泄,这些离子会平衡细胞的能量状态,使菌体进行正常运动,还具有代谢控制、溶质转运等作用[23];因此,测定电导率以表征松油烯-4-醇对荧光假单胞菌细胞膜的渗透性变化。如图4所示,在加入松油烯-4-醇后0~6 h,1 MIC与2 MIC处理的荧光假单胞菌电导率迅速增加,明显高于空白对照组,且2 MIC样品的电导率大于1 MIC。表明随着松油烯-4-醇的剂量增加,其电导率增大,松油烯-4-醇破坏了荧光假单胞菌的细胞通透性,进而导致细胞内离子外渗;6~12 h,两种剂量的松油烯-4-醇处理组增长都趋于平缓,表明松油烯-4-醇能有效改变荧光假单胞菌细胞膜的通透性,使膜内外电导率增加。此结果与测定AKP活力得到的结果都表明松油烯-4-醇能破坏荧光假单胞菌的细胞壁且改变细胞膜的通透性。Zhang Yunbin等[24]研究了肉桂精油对大肠杆菌的抗菌机制,也发现加入肉桂精油会使菌液电导率升高,对细胞膜通透性造成影响,与本实验结果相同。

图4 不同剂量的松油烯-4-醇对荧光假单胞菌电导率的影响Fig.4 Effect of terpinene-4-ol at different concentrations on the electrical conductivity of Pseudomonas fluorescens
2.4 松油烯-4-醇对荧光假单胞菌细胞膜通透性的影响
FDA是一种本身不发荧光且不带电荷的物质,因其是脂质性小分子,更易透过细胞膜进入细胞,在胞内被非特异性脂酶催化,FDA会产生黄绿色的荧光素。当细胞膜完整,FDA会在激发和发射波长分别为488 nm和530 nm处被检测出;当细胞膜遭到破坏,FDA很快流出细胞,不能被检测到。通过测定FDA荧光强度的大小,可反映出细胞膜的通透性与完整性[25]。如图5所示,经松油烯-4-醇对荧光假单胞菌处理2 h后,1、2、4 MIC处理组与空白对照组测定的荧光强度差异不大,表明经松油烯-4-醇处理的荧光假单胞菌的细胞膜完整性较好,仅细胞膜通透性有所降低;经松油烯-4-醇处理5 h和8 h,1、2、4 MIC处理组荧光强度随剂量增加及时间的延长而降低,表明从培养5 h起,松油烯-4-醇能大幅增强荧光假单胞菌细胞膜的通透性,并可能对细胞膜产生破坏作用,导致FDA外渗,荧光强度降低。随着松油烯-4-醇剂量的增加及培养时间的延长,对荧光假单胞菌的损伤程度也越大。

图5 松油烯-4-醇对荧光假单胞菌FDA荧光强度的影响Fig.5 Effect of terpinene-4-ol on fluorescence intensity of Pseudomonas fluorescens cells stained with FDA
2.5 松油烯-4-醇对荧光假单胞菌超微结构的影响
SEM可以更直观、更清晰地观察到松油烯-4-醇对荧光假单胞菌菌体损伤情况。图6A是正常的荧光假单胞菌(为空白对照组),其菌体饱满、完整、且表面光滑的短杆状形态;经1 MIC与2 MIC的松油烯-4-醇处理后(图6B、C),菌体表面变得粗糙,细胞膜均有破裂,2 MIC处理组菌体皱缩塌陷,与1 MIC处理组相比,其细胞膜破坏程度更为严重;图6D为4 MIC松油烯-4-醇处理组,其菌体破碎、部分菌体被溶解、细胞膜几乎完全破裂。上述结果表明1、2、4 MIC的松油烯-4-醇处理组能对荧光假单胞菌细胞壁和细胞膜产生不可逆的损坏,且破坏程度随着松油烯-4-醇剂量增大而增强,细胞壁及细胞膜的破坏导致细胞内容物外泄,菌体干瘪并产生皱缩,从而抑制荧光假单胞菌的生长。植物精油类抗菌物质,因其疏水性更易作用于细胞膜,使细胞膜破损,并增强了细胞膜的通透性[26]。此结果与抑菌曲线结论一致,松油烯-4-醇能够抑制荧光假单胞菌的生长。Zhao Yi等[27]用SEM观察发现没食子酸能改变金黄色葡萄球菌的疏水性,对细胞膜造成破坏,导致细胞内成分泄露,与本研究结果相同。

图6 不同剂量松油烯-4-醇处理对荧光假单胞菌的SEM图Fig.6 SEM images of Pseudomonas fluorescens treated with different concentrations of terpinene-4-ol
2.6 松油烯-4-醇对荧光假单胞菌DNA含量的影响
DNA中含有合成RNA和蛋白质所必需的遗传信息,是生物发育和正常运转所不可或缺的大分子物质。DNA含量的减少和损伤导致基因无法正常表达,使胞内的酶与受体合成受阻,最终导致其衰亡[28]。琼脂糖凝胶阻滞电泳中的DNA条带亮度与DNA分子大小成正比,条带亮度能够反映DNA含量。如图7可知,经3 h的松油烯-4-醇培养后,空白对照组的DNA条带明亮清晰,0.5、1、2、3 MIC处理组的DNA条带亮度依次随着松油烯-4-醇的剂量增大而降低,4 MIC处理组检测不出荧光假单胞菌的DNA。以上结果表明松油烯-4-醇能破坏荧光假单胞菌的细胞膜,使胞内DNA泄露,且随着松油烯-4-醇的剂量增大,对荧光假单胞菌细胞膜的破坏也越强,胞内剩余的DNA含量越少。此结果与上文中SEM、电导率、FDA实验结果相印证,松油烯-4-醇能够有效破坏荧光假单胞菌细胞膜,不仅导致小分子外泄,大分子的DNA也会随之泄露出来。Shen Suxia等[28]研究发现肉桂醛对金黄色葡萄球菌产生抑菌作用时菌体细胞壁和细胞膜破裂,内部空化,核酸大分子物质外渗。
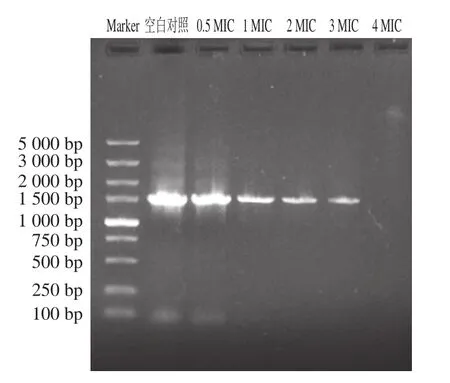
图7 松油烯-4-醇对荧光假单胞菌DNA的影响Fig.7 Effect of terpinen-4-ol on DNA of Pseudomonas fluorescens
2.7 松油烯-4-醇对荧光假单胞菌总蛋白质量浓度的影响
蛋白质是生命物质的基础,是构成细胞最重要的成分,具有维持细胞内渗透压、酸碱度、运输代谢物等重要作用,蛋白质的降低会直接影响细胞的活力[29]。由图8可知,空白对照组中随着培养时间的延长,荧光假单胞菌代谢繁殖,其胞内蛋白质量呈现上升趋势;经1 MIC和2 MIC松油烯-4-醇处理0~3 h的菌体,胞内蛋白质量浓度迅速降低,此时松油烯-4-醇对菌体细胞膜破坏严重,蛋白质大量外渗;3~12 h两处理组的蛋白质量浓度均缓慢下降,可能是0~3 h松油烯-4-醇对细胞膜破坏较严重,蛋白质已较多泄露所致。测定结果表明松油烯-4-醇能大幅度且迅速破坏荧光假单胞菌的细胞膜,使其通透性增强,导致大分子蛋白质泄露,并且随着松油烯-4-醇的剂量增加,对细胞内蛋白的含量影响越大,此结果与SEM、核酸电泳结果相印证。松油烯-4-醇也可能会破坏细胞内蛋白质的合成,从而影响细胞的生长与代谢。Yin Lizi等[29]研究了肉桂醛对嗜水气单胞菌的抑菌机制,表明肉桂醛可以降低细胞中的蛋白含量,干扰细胞代谢,影响了细胞膜的通透性。

图8 松油烯-4-醇对荧光假单胞菌总蛋白质量浓度的影响Fig.8 Effect of terpinen-4-ol on total protein concentration of Pseudomonas fluorescens
2.8 松油烯-4-醇对荧光假单胞菌细胞内Na+、K+-ATP酶活力的影响
Na+、K+-ATP酶是一种处于细胞质膜磷脂双分子层中的蛋白酶,能催化ATP水解为细胞直接提供能量,在物质运输、能量转化、调节膜电位及传递信息方面具有重要作用,细胞膜的损伤和重要离子的泄露会对细胞膜相关的能量转导系统产生影响[30]。如图9所示,空白对照组0~6 h因荧光假单胞菌的增殖,胞内Na+、K+-ATP酶活力增强;1、2 MIC处理组在0~6 h培养后,Na+、K+-ATP酶活力与空白对照组相比明显降低,且2 MIC松油烯-4-醇对荧光假单胞菌Na+、K+-ATP酶抑制能力更为出色。对测定结果分析可知,松油烯-4-醇可能对荧光假单胞菌细胞内的蛋白质有明显抑制作用,从而阻碍了胞内酶的合成,Na+、K+-ATP酶也遭到破坏导致胞内ATP不能正常供给能量,最终细胞凋亡。此结果与松油烯-4-醇对胞内总蛋白、电导率、核酸电泳的影响结果相印证,表明松油烯-4-醇能有效破坏荧光假单胞菌细胞膜的完整性,增加离子的通透性及细胞内Na+、K+-ATP酶的外渗。Sharma等[31]研究表明大蒜精油会显著降低蜡状芽孢杆菌胞内ATP的释放。
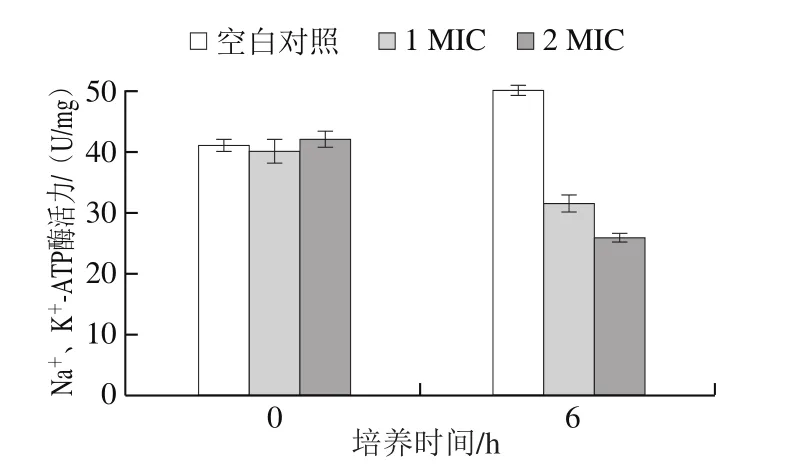
图9 松油烯-4-醇对荧光假单胞菌Na+ 、K+-ATP酶活力的影响Fig.9 Effect of terpinen-4-ol on Na+, K+-ATPase activity in Pseudomonas fluorescens
3 结 论
天然植物精油类物质具有安全无毒、抗菌光谱性强、原料来源广泛等特点,作为食品防腐剂日益受到众多学者们的关注[32]。松油烯-4-醇是茶树精油的主要有效成分之一,也是多种天然植物精油所含的单萜类成分,具有抗肿瘤、抗炎等作用,但对其抑菌能力报道尚少。
本研究从细胞机制方面探讨了松油烯-4-醇对荧光假单胞菌的抑制作用。本实验首先确定了松油烯-4-醇对荧光假单胞菌具有良好的抑菌效果,且明确其MIC为2 μL/mL;由时间抑菌曲线进一步看出松油烯-4-醇可较为迅速地抑制荧光假单胞菌的生长;AKP活性实验表明加入松油烯-4-醇能有效破坏菌体细胞壁,使细胞膜暴露出来;电导率测定结果证明松油烯-4-醇可以改变细胞膜的通透性,使胞内离子外泄;胞内荧光强度测定结果证实松油烯-4-醇能大幅增强细胞膜的通透性,并可能对细胞膜产生破坏作用;SEM观察到松油烯-4-醇对细胞膜造成了不可逆的损伤,大分子的DNA与蛋白质会从细胞内泄露,胞内的Na+、K+-ATP酶含量也会降低,导致ATP无法正常提供细胞能量,直接造成细胞凋亡。
猜你喜欢
中国调味品(2022年9期)2022-08-30
当代水产(2022年1期)2022-04-26
心肺血管病杂志(2020年5期)2021-01-14
读与写(2019年31期)2019-11-26
食品界(2019年2期)2019-03-10
中国实用医药(2016年16期)2016-07-26
天津农业科学(2016年3期)2016-03-12
科学中国人(2015年13期)2015-02-28
中国民族民间医药·下半月(2014年10期)2014-10-27
创新科技(2014年12期)2014-07-27
