
甘肃省扎尕梁北坡头花杜鹃枝叶性状特征
2022-02-15 02:48杨克彤陈国鹏鲜骏仁俞筱押张金武王立
草业学报 2022年2期
杨克彤,陈国鹏*,鲜骏仁,俞筱押,张金武,王立
(1.甘肃农业大学林学院,甘肃 兰州 730070;2.四川农业大学环境学院,四川 成都 611130;3.黔南民族师范学院旅游与资源环境学院,贵州 都匀 558000)
植物与生境之间的相互关系是当今植物生态学研究的热点问题[1-3]。植物功能性状是植物长期进化形成的能够反映物种对外界环境适应策略的核心属性,不仅可以揭示植物对环境的响应和适应机制,还可以反映植物对资源的分配和利用规律[3-5]。阐明不同器官功能性状对异质环境的响应及其相互关系是理解植物进化与适应的基础[6-10]。加强对环境变化响应敏感器官的研究,可以及时反映植物的生长和适应动态,在揭示物种性状的进化表达、资源获取与物种分布等方面具有重要意义。
叶片和小枝作为植物与外界环境接触最密切的部分,是对环境变化感知最敏锐的构件之一[8-11],其性状可塑性能够较为全面地展示植物对异质环境的适应与生存策略。叶片是光合产出的最重要器官,其功能性状不仅直接影响植物碳收益,还可以平衡、转换、弥补植物与外界环境的差异从而决定植物的生存状态。小枝作为叶片生长的平台,主要为叶片输送养分并调整叶片受光面的夹角。此外,小枝的更新发育进一步决定了植物的外部结构和空间特征[11-13],不仅维持了植物生存空间的稳定,其向外扩展的特性还剧烈影响着物种间的竞争格局[14]。叶片和小枝的自我更新能力明显优于其他构件,能够迅速对环境变化做出响应并保持相对稳定[15-16],为深入探究植物对环境的适应提供了基础,因其性状特征具有普适性与易测性,也因此成了研究植物在异质生境中性状表达的首选指标。
海拔梯度包含了温度、气压、水分和辐射等一系列具有连续分布特征的环境因子,是研究植物适应异质生境变化的最佳场所[1,3,17-21]。不同植物在适应海拔异质生境时的生存策略各不相同,多数植物主要通过功能性状的权衡与适应来寻求最佳的生存状态[21-24],具体表现在:随海拔上升个体大小差异、器官间生物量分配、异速生长与权衡、枝叶形态异化或内含物含量的波动以及超微结构变化等多个方面[22-25]。由海拔变化引起的环境差异会导致植物产生生理及形态上的多重响应,并逐步经过“瀑布式上升效应”可能会引起整个生态系统的转变[1,26-27],因此海拔梯度细微的变化有时也会带来巨大的结果差异。虽然近年来部分研究在群落功能性状随海拔梯度变化方面已有所成果,但部分结论存在差异甚至相反,并且群落水平的性状变化掩盖了物种的个体特征,导致其对生态系统的预测精度不高[28-29],而在单物种尺度上开展枝叶性状与环境关系的研究可以深入了解关键植物适应机制并凸显环境对其功能性状的影响。
山地生态系统是对全球气候变化最敏感的区域,是研究植被与气候关系的天然实验室[1,30]。头花杜鹃(Rhododendron capitatum)作为典型的高山植物,是维持青藏高原东北缘山地生态系统稳定的重要类群。本研究以甘肃扎尕梁北坡的优势种头花杜鹃为研究对象,测定其23组枝叶功能性状,探究海拔梯度下头花杜鹃枝叶性状的适应与响应模式,以进一步解析高山植物对海拔梯度的应对策略。基于此,本研究试图回答以下问题:1)随海拔梯度上升,头花杜鹃枝、叶性状的变化有何特征与规律?2)性状间的相互关系如何?3)不同性状的表型分化有何特征?
1 材料与方法
1.1 研究区概况
研究区位于甘肃省舟曲县插岗梁自然保护区(103°57′05″-104°42′05″E,33°14′32″-33°53′52″N,海拔1173~4536 m),保护区处在青藏高原东北缘与秦岭西翼及岷山山脉交汇地带,属西秦岭地质构造带南部陇南山地[7]。该区域处于我国温带季风、亚热带季风和高原山地气候带的汇合区域及半湿润和半干旱区的分界线上。近30年来,林区年平均降水量为951.0 mm,年平均蒸发量918.8 mm,相对湿度为82.0%,干燥度0.5,年平均气温4.3℃,最冷月(1月)平均气温为-13.3℃,最热月(7月)平均气温为20.8℃,全年无霜期96.7 d(最多143 d,最少53 d);日平均气温≥5℃的日数为178 d,年积温1994.6℃;日平均气温≥10℃的日数为103 d,年积温1387.9℃。年日照时数为1398.4 h,日照百分率为32.0%。植被分布的垂直带谱明显,从低到高依次为干旱河谷灌丛、亚高山次生林、亚高山暗针叶林和高山杜鹃灌丛[25]。头花杜鹃植株主要分布在该区域海拔3100~3700 m范围内,在3400 m以下主要为单株零星分布或可形成小规模种群,3400 m以上与陇蜀杜鹃(Rhododendron przew alskii)混生形成灌丛群落。
1.2 数据采集与测定
于2019年8月在研究区进行全面勘查后,在插岗梁保护区的扎尕梁北坡的林线到山顶处自下而上沿等高线按海拔高度3400、3500、3600和3700 m设置调查样线。在样线中随机选取株高50 cm左右的标准木,以保证植株年龄基本一致,并进行小枝与叶片的采样。每个海拔梯度共采集30余个小枝,并放于自封袋内,叶片夹放在湿润滤纸中,并置于便携式低温箱中带回。本研究所述小枝定义为植株顶端无分支的1年生茎。
小枝与叶片带回后即刻测定各项指标,每个海拔随机测量15个样本。对于小枝,首先剪下小枝上的叶片,然后测量小枝基径(stem diameter,SD)、小枝长(stem length,SL),再用扫描仪(EPSON V 700)对所有叶片进行扫描并求出总叶面积(total leaf area,TLA)和叶片数(leaf number,LNO.),用电子天平测量小枝鲜重(stem fresh weight,SFW)、小枝干重(stem dry weight,SDW)、总叶鲜重(total leaf fresh weight,TLFW)和总叶干重(total leaf dry weight,TLDW);对于单叶,先用天平称量叶片鲜重(leaf fresh weight,LFW),再用扫描仪进行扫描并求出单叶面积(leaf area,LA),随后测量叶厚(leaf thickness,LT)与叶干重(leaf dry weight,LDW),测量叶厚时靠近叶片中心但避开主脉,测量3次求取平均值后作为叶片厚度。测量小枝与叶片干重时,将其置于玻璃皿放入烘箱,120℃杀青30 min后转为75℃烘至恒重后称取干质量。叶长(leaf length,LL)、叶宽(leaf width,LW)、单叶面积(LA)与总叶面积(TLA)均使用Image J面积测量软件进行。鲜、干质量的测定均使用电子天平(精确至0.0001 g),长、宽、厚、径均使用数显游标卡尺(精确至0.01 mm)测量。由于头花杜鹃叶柄极短,难以测量,所以用解剖刀将叶柄除去而不纳入测定指标,因此本研究所述叶片性状均不包括叶柄。
相关枝叶性状的推算如下[7,10]:叶形指数(leaf shape index,LSI)=叶长/叶宽,比叶面积(specific leaf area,SLA)=叶面积/叶干质量,叶干物质含量(leaf dry matter content,LDMC)=叶干重/叶鲜重,茎干物质含量(stem dry matter content,SDMC)=茎干重/茎鲜重,叶体积(leaf volume,LV)=叶面积×叶厚,茎体积(stem volume,SV)=π×茎长×(茎径/2)2×1/3,叶密度(leaf density,LDE)=叶干重/叶体积,茎密度(stem density,SDE)=茎干重/茎体积,纤细率(stem slender ratio,SSR)=茎长/茎径,总比叶面积(total specific leaf area,TSLA)=总叶面积/总叶干质量,总叶干物质含量(total leaf dry matter content,TLDMC)=总叶干重/总叶鲜重,总叶茎重比(total leaf and stem weight ratio,TLSWR)=总叶干重/总茎干重。
1.3 数据分析
采用单因素方差分析(one-way ANOVA)检验海拔高度对头花杜鹃枝叶功能性状的影响,并采用Tukey’s HSD多重比较检验性状在不同海拔间的差异显著性;利用方差分析提取海拔间与海拔内及随机误差的方差分量并计算各性状的表型分化系数(Vst),表型分化系数Vst=δ2t/s/(δ2t/s+δ2s),其中δ2t/s为海拔间方差值,δ2s为海拔内方差值[31-33];采用Pearson相关系数、双尾检验进行海拔与性状间的相关分析,最后用单因素方差分析检验海拔内与海拔间方差分量的显著性。利用Excel 2010和SPSS 20.0进行数据统计、分析,用Origin 9.1绘图。
2 结果与分析
2.1 头花杜鹃枝叶性状随海拔的变化
头花杜鹃叶性状随海拔梯度变化差异显著,除LSI无显著变化外,其他性状均有显著变化(图1),LL、LW、LA、LV和LDW在3600 m时显著小于其他海拔;LDMC和LDE随海拔上升逐渐增大,并在海拔3700 m时显著高于其他海拔;而SLA逐渐减小,海拔3700 m时显著小于其他海拔;LT整体呈增大趋势。小枝性状除SD、SDMC、LNO.和SDE随海拔梯度无显著变化外,其他性状均变化显著(图2),其中SL、SDW、TLDW、TLA、SSR和SV在3600 m显著小于其他海拔,TSLA呈下降趋势,TLDMC呈上升趋势,TLSWR则先增大后减小。
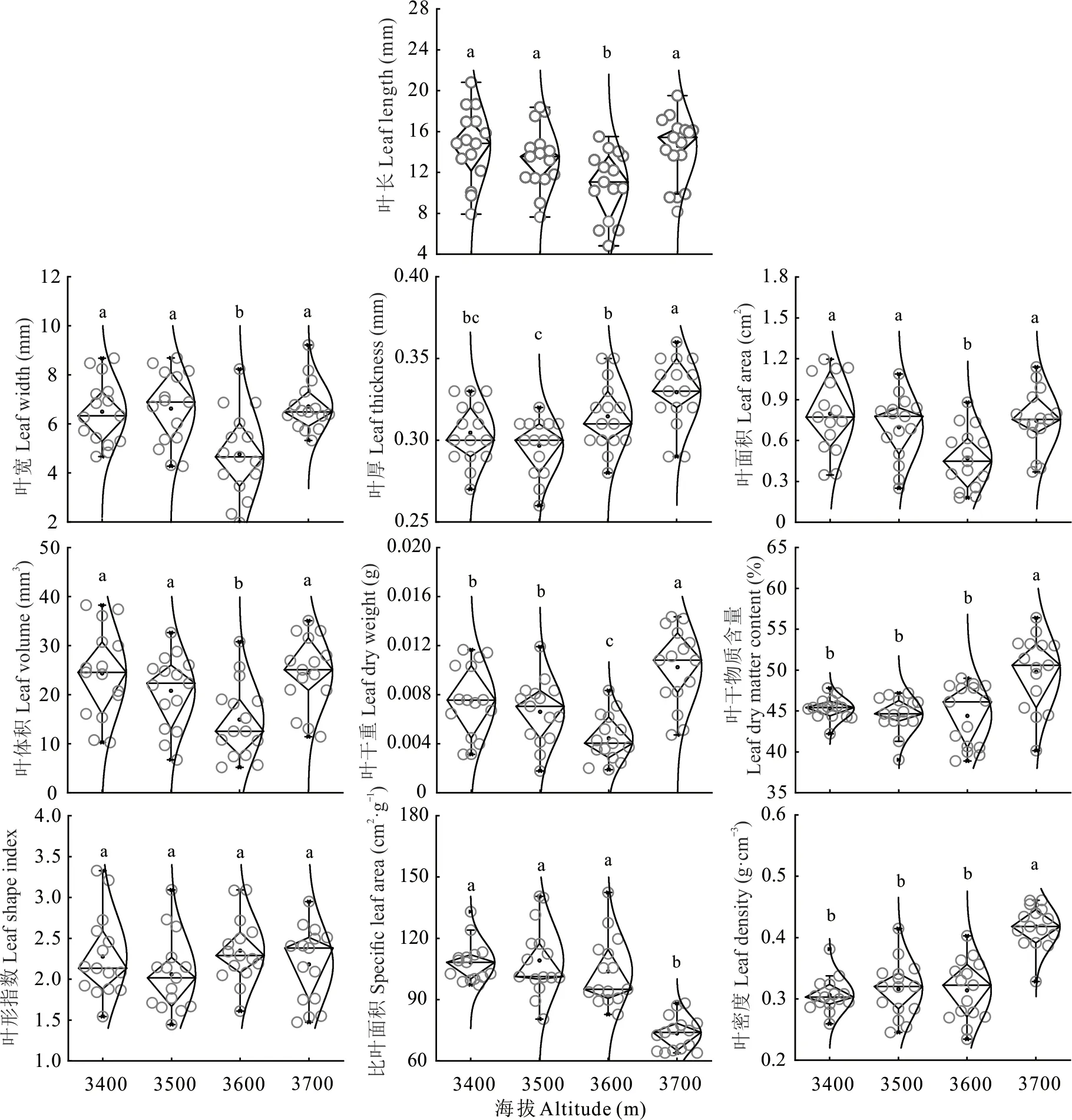
图1 头花杜鹃叶性状随海拔的变化Fig.1 Variation of R.capitatum leaf traits with altitude
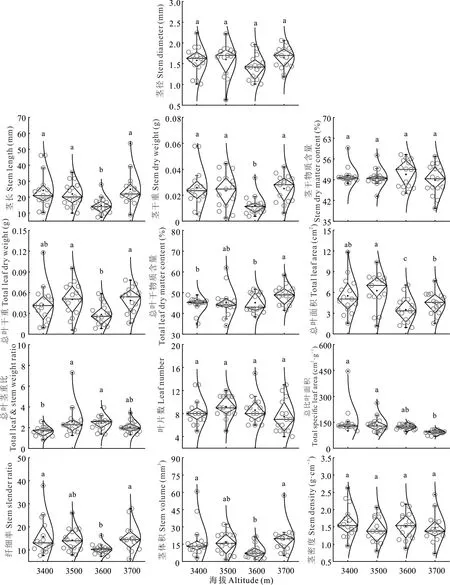
图2 头花杜鹃小枝性状随海拔的变化Fig.2 Variation of R.capitatum stem tr aits with altitude
2.2 枝叶性状间的相互关系
头花杜鹃大部分枝叶性状间均为显著正相关(图3),LL、LW、LA、LV和LDW均与SD、SL、SDW、TLDW、TLSWR和SV显著正相关,LT、LSI和SLA与多数性状显著负相关,小枝水平的SDMC、TLSWR、LNO.、SSR和SDE与部分性状的相关性较弱,总体而言小枝水平不及叶片水平的相关性强。
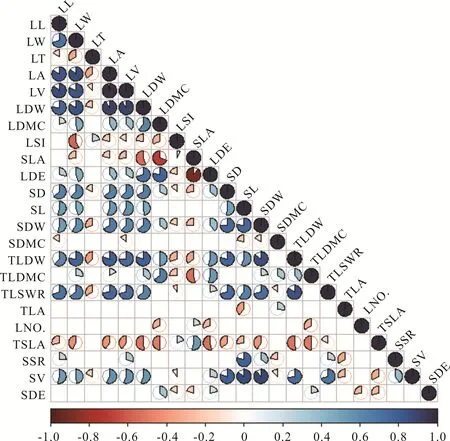
图3 头花杜鹃枝叶性状间的相关关系Fig.3 Relationship between stem and leaf traits of R.capitatum
2.3 枝叶性状的表型分化
头花杜鹃枝叶性状在海拔间和海拔内的方差分量百分比均值分别为17.30%和70.76%,随机误差为11.94%(表1),表型分化系数整体介于3.19%~57.73%,均值为20.65%,仅8组性状表型分化系数高于均值,占所有性状(23组)比例的34.78%。头花杜鹃在同一海拔内的方差分量显著高于海拔间,其中LSI、SD、SDMC、LNO.和SDE的表型分化系数较低(小于11.11%,下四分位数),LT、LDW、LDMC、SLA和LDE的表型分化系数较高(大于25.41%,上四分位数),即在应对海拔梯度引起的异质生境时,这10组性状分别表现出较低的变异性与较高的可塑性,表明头花杜鹃枝叶性状变化范围大、表型变异丰富。
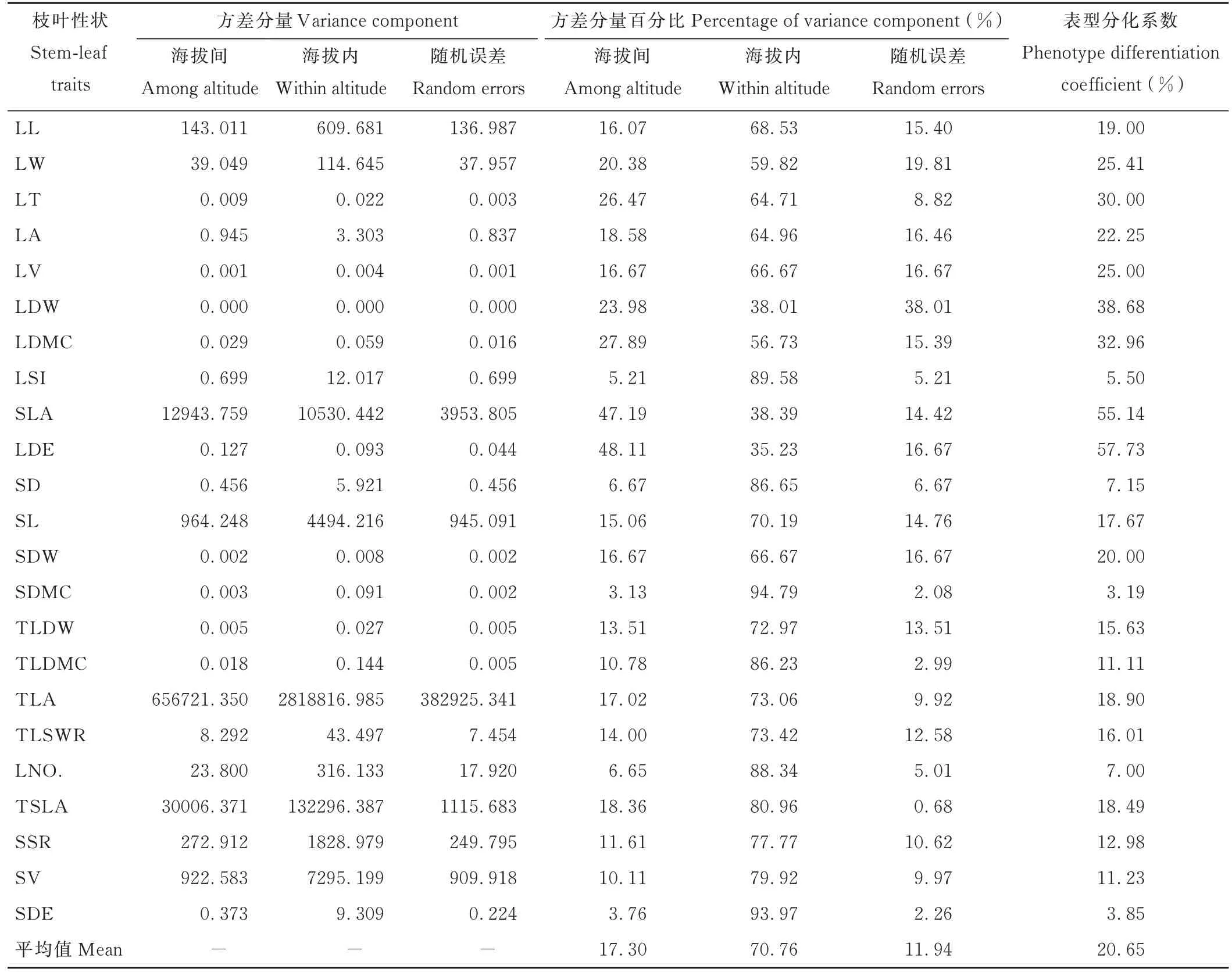
表1 头花杜鹃枝叶性状的方差分量与表型分化系数Table 1 Var iance component and phenotype differentiation coefficient of stem-leaf tr aits of R.capitatum
3 讨论
海拔梯度引起的生境变化会显著影响植物功能性状,并且部分性状也会随之表现出梯度变化,这取决于性状对环境的敏感程度与自身功能特性[32-34]。本研究中,头花杜鹃叶长、叶宽、叶面积、叶体积、叶干重、茎长、茎干重、总叶干重、总叶面积、纤细率和茎体积均在3600 m处显著减小,整体呈“V”形变化(图1和图2)。可能是由于山地植被逐步从林线过渡到树线、再过渡到高寒杜鹃灌丛后,杜鹃灌丛抗环境干扰能力强,土壤理化性质相对稳定,头花杜鹃产生适应性策略,因此3600 m可能是生境适应连续变化梯度的转折点[25]。叶厚、叶干物质含量、叶片密度和总叶干物质含量均随着海拔梯度上升而增大,与以往研究结果一致[34-35]。随着海拔升高,温度降低、光照辐射增强,叶片将资源集中于构建防御结构[36],一方面可以降低辐射伤害,另一方面增加叶厚有助于抵抗低温冻害。茎干物质含量降低则可能与单叶面积减小有关,Corner[28]法则认为:大叶片需要较大的枝条支撑并输送养分,随着海拔升高,单叶面积减小,由此导致植株对茎的投资减少,这种策略实现了有限资源的最大利用效率。通常情况下,分布在资源有限生境及生长缓慢的物种比叶面积都较低,不论草本、灌木与乔木,还是极地、温带与热带,植物的比叶面积变化都随着海拔上升而降低[2-3,35],导致这种情况的原因则主要与温度下降有关,低温环境下,叶肉细胞增大、表皮细胞壁增厚,导致叶片厚度增加,因此比叶面积下降[14]。
植物在应对异质环境时,并非某一性状采取适变策略,而是多个性状间的组合(即功能性状组)产生共变,这种共变机制更有利于应对环境引起的激变[22,30]。本研究中,仅7组性状与海拔梯度为极显著(或显著)相关,其余16组性状均不显著,即这7组性状可能是枝叶功能性状的组合中心。由这7组性状产生较为剧烈的应对策略并组合其他性状产生微小变化便足以应对海拔梯度引起的生境异质,这种应对策略减少了能量损耗与成本投资,具有高效率、低能耗的特点[22]。此外,叶长、叶宽、叶面积、叶体积、叶干重均与茎径、茎长、茎干重、茎体积、总叶干重极显著正相关,说明头花杜鹃枝叶性状支持Corner[28]法则,即较大的叶片需要大的枝条来承担;另一方面可能随着海拔升高,温度降低,土壤水分难以利用,导致头花杜鹃产生干旱胁迫,加之较小枝条在低温环境运送水分与养分时容易产生栓塞,较大且健壮的枝条有助于缓解栓塞并抵御高山地区大风引起的损伤[29-30]。
枝叶性状的变异与表型分化共同决定了其变异程度与范围,较强的变异与较大的可塑性有助于应对高山地区极端气候。本研究发现,头花杜鹃枝叶性状表型分化系数均值为20.65%(表1),低于红椿(Toona ciliata,80.73%)、砂生槐(Sophora moorcroftiana,39.25%)、珙桐(Davidia involucrata,37.00%)、尾叶樱桃(Cerasus dielsiana,30.78%),而高于云南红豆杉(Taxusyunnanensis,14.66%)、红松(Pinuskoraiensis,12.39%)、马尾松(Pinus massoniana,6.44%),说明其表型变异程度低于一般的阔叶树种却高于针叶树种。有研究认为生存环境会极大地影响表型分化程度[31-35],头花杜鹃长期生活于低温与强光辐射等不利环境中,环境因子的多变与复杂可能是造成其表型分化相对保守的主要原因。另外,枝叶性状中仅8组(叶宽、叶厚、叶面积、叶体积、叶干重、叶干物质含量、比叶面积和叶密度)性状的表型分化系数高于平均值(20.65%),说明这8组性状具有相对丰富的表型变异,尤其比叶面积与叶密度表型分化系数高达55.14%和57.73%,可能是应对高山环境海拔异质的核心性状[32-36]。叶形指数、茎径、茎干物质含量、叶片数和茎密度表型分化系数较低(<11.11%),说明这些性状具有稳定特质,并不轻易随海拔梯度做出改变,其稳定特质可能对于维持植物枝叶生长发育有特殊意义,如:极端环境中维持生存的下限等[13-17]。另外,头花杜鹃枝叶性状海拔内变异普遍高于海拔间变异,即同一海拔内的居群是产生丰富表型变异的主要来源,可能是应对生境异质的主要贡献者,也可能由于在研究区范围内头花杜鹃分布集中,并未产生地理隔离,从而导致海拔间居群表型变异较弱。
头花杜鹃枝叶功能性状对环境的适应有助于其应对高寒生境,可在一定程度上改善其存活、生长和繁殖状态。头花杜鹃73.91%(17组/23组)的枝叶性状均在3700 m显著增大,这种策略可能是为了避免较低海拔(3400 m)的天敌或中海拔(3500~3600 m)的物种竞争;也可能因为头花杜鹃属于典型的高山植物,在长期的生物演化与自然选择过程中已经适应了高山特殊环境,其固有的生态特性可能更有助于头花杜鹃在更高海拔而非低海拔生存[7-12]。
4 结论
头花杜鹃主要以枝叶功能性状组合与协变共同应对高山生境的不利因素;部分性状在海拔3600 m显著减小,大部分性状在海拔3700 m达到最大;海拔内表型变异普遍大于海拔间,叶片的表型分化系数显著大于小枝。头花杜鹃枝叶功能性状对高山生境的积极响应与适应在提高头花杜鹃生境适合度、占据稳定生态位、维持高山物种竞争格局与生物多样性等方面具有重要意义,在未来的研究中应继续加强对高山物种的关注。
猜你喜欢
中华医学图书情报杂志(2022年1期)2022-11-18
农学学报(2022年8期)2022-11-16
农业与技术(2022年16期)2022-09-01
中国现代医生(2022年21期)2022-08-22
农村科学实验(2022年2期)2022-03-12
三农资讯半月报(2020年2期)2020-03-09
华东师范大学学报(自然科学版)(2019年3期)2019-06-24
福建基础教育研究(2019年3期)2019-05-28
西部资源(2018年1期)2018-11-01
江苏农业科学(2016年9期)2016-11-28
