
黑腹果蝇头部中RNA结合蛋白RBP9的互作蛋白筛选
2022-02-08 02:04:08时佳园顾佳蕙汪生鹏张国政李木旺
昆虫学报 2022年12期
时佳园,马 达,顾佳蕙,秦 笙,2,汪生鹏,2,张国政,2,李木旺,2,孙 霞,2,*
(1.江苏科技大学生物技术学院,江苏镇江 212018;2.中国农业科学院蚕业研究所,江苏镇江 212018)
在真核生物中,RNA结合蛋白(RNA-binding protein,RBP)是基因表达过程中重要调控因子,主要参与调控转录、转录后和翻译过程。ELAV/Hu(embryonic lethality abnormal vision system/human)家族是一类高度保守的RNA结合蛋白,在多细胞生物的神经系统中表达。该家族编码基因elav首次在黑腹果蝇Drosophilamelanogaster中被发现(Camposetal.,1985),随后在多种生物中被发现(Sunetal.,2021;Hilgers,2022)。
双翅目黑腹果蝇ELAV/Hu家族包括ELAV,RBP9和FNE,三者均含有3个RNA识别基序(RNA recognition motifs,RRMs)。ELAV和FNE表达于神经系统中,前者分布于细胞核中,后者分布于细胞质中,二者的表达贯穿黑腹果蝇的整个发育周期(Samson and Chalvet,2003)。RBP9从3龄幼虫开始表达,存在于成虫神经系统的细胞核和卵子发生时包囊细胞的细胞质中(Kim-Haetal.,1999)。ELAV在黑腹果蝇神经系统的发育与分化、选择性剪接中具有重要调控作用(Lisbinetal.,2001;Simionatoetal.,2007)。ELAV能够结合在pre-mRNA的3′内含子和自身3′UTR,参与靶标基因和自我表达调控(Lisbinetal.,2001;Borgeson and Samson,2005)。ELAV功能完全缺失会导致胚胎致死。FNE在蘑菇体(mushroom body)发育、神经突触形成、交配行为中具有重要调控作用(Zaninietal.,2012;Olesnickyetal.,2014)。研究发现FNE蛋白表达水平受性别决定因子SXL(sex lethal)的调控,而不受TRA(transformer)和TRA2(transformer 2)的调控(Sunetal.,2015)。RBP9在卵巢发育分化、血脑屏障形成和RNA稳定性中具有重要调控作用(Kimetal.,2010;Tobaetal.,2010)。RBP9功能缺失导致黑腹果蝇寿命缩短。ELAV,RBP9和FNE通过选择性多聚腺苷酸化(alternative polyadenylation,APA)和最后一个外显子可变剪接(alternative last exon,ALE)对mRNA 3′UTR长度进行转录后调控,且三者在黑腹果蝇的生长发育过程中存在重要的相互作用(Zaharievaetal.,2015;Weietal.,2020;Leeetal.,2021)。
为了进一步了解RBP9蛋白在神经系统发育中的功能,我们首先利用CRISPR/Cas9技术构建了3×Flag标签与RBP9融合表达的转基因黑腹果蝇,其次,以此转基因黑腹果蝇为材料,使用质谱法分析了在神经系统中与RBP9相互作用的蛋白质。本研究结果为进一步探索RBP9在黑腹果蝇神经系统发育中的生物学功能提供了重要信息。
1 材料与方法
1.1 试虫
黑腹果蝇品系在25℃标准培养基中进行培养和杂交。利用CRISPR/Cas9技术构建转基因黑腹果蝇品系3×Flag-RBP9/3×Flag-RBP9 (3FRBP9)和V5-FNE/V5-FNE(VFNE),分别在RBP9和FNE序列N端编码序列插入3×Flag和V5标签编码序列(Renetal.,2013)。在RBP9和FNE起始密码子ATG附近分别设计1个sgRNA靶点,其序列分别为:5′-CAGCGTTCGCAAGATGGTCGAGG-3′;5′-AA TCACAATGACCAACGCCATGG-3′(粗体代表起始密码子)。将sgRNA序列利用酶切方法构建到表达质粒pU6a中。在RBP9和FNE起始密码子ATG后分别插入3×Flag和V5标签编码序列,在该位置的上下游分别扩增约1 kb作为供体质粒的同源臂,再利用overlap PCR将两个同源臂序列连接并克隆至pMD-19T载体。pU6a和供体质粒的黑腹果蝇胚胎的显微注射由珠海联合华益科技有限公司(UniHuaii)完成,其中胚胎注射的黑腹果蝇为转基因株系nos-Cas9(attP2),每个基因注射300粒胚胎,注射后胚胎置于正常培养基上于27℃至孵化,之后在正常条件下饲养。单头G0黑腹果蝇分别与野生型黑腹果蝇进行杂交,杂交后留取G0黑腹果蝇基因组,分别对其利用同源臂、起始密码子ATG后的标签编码序列进行克隆测序(引物序列见表1),筛选保留序列正确的黑腹果蝇后代,并将其与黑腹果蝇野生型品系(Bloomington no.5905)回交6代,消除潜在的脱靶可能同时获得遗传背景均一的转基因黑腹果蝇。将杂合转基因黑腹果蝇自交和杂交,分别构建纯合个体3FRBP9/3FRBP9(3FRBP9),VFNE/VFNE(VFNE)和V5-FNE/V5-FNE;3×Flag-RBP9/3×Flag-RBP9(VFNE;3FRBP9)。

表1 引物信息Table 1 Primer information
1.2 免疫沉淀
分别收集黑腹果蝇转基因品系3FRBP9,VFNE和VFNE;3FRBP9以及野生型品系成虫头部各600个,用1 mL裂解缓冲液[50 mmol/L Tris-HCl(pH 8.0),50 mmol/L NaCl,1 mmol/L EDTA,1% Triton X-100,0.5% NP40,1 mmol/L PMSF和1×Protease Inhibitor(Roche,瑞士)]冰上研磨。超声破碎。低温离心后取上清。将上清分别与anti-V5和anti-FLAG M2磁珠(Sigma,美国)孵育。用裂解缓冲液洗涤磁珠4次,去上清后加入50 μL的1×蛋白上样缓冲液,重悬后煮沸,提取RBP9蛋白。上清样品使用anti-FLAG(Sigma,美国)单克隆抗体进行Western blot或银染分析。
1.3 蛋白质谱实验
将1.2节免疫沉淀得到的3FRBP9和野生型品系成虫头部中RBP9蛋白样品进行免疫沉淀后,洗脱样品送公司进行蛋白质谱分析。使用TripleTOF 5600+Eksigent nanoLC质谱联用系统(SCIEC,美国)进行质谱检测,分析头部中RBP9的互作蛋白。整个过程由武汉金开瑞生物工程有限公司(Wuhan GeneCreate Biological Engineering Co.,Ltd.)完成。利用ProteinPilot 4.5软件和数据库uniprot_Drosophila_melanogaster蛋白测序数据(包括43 023条蛋白序列)对质谱数据进行蛋白质鉴定(2012年7月;AB Sciex,美国)。对鉴定的蛋白结果进行过滤,肽段Unused Protscore>1.3(可信度≥95%)认为是可信肽段,且保留包含unique peptide≥1肽段的蛋白质。
1.4 GO和KEGG富集分析
鉴定的190个与RBP9相互作用蛋白编码基因的GO和KEGG信息由KOBAS 3.0软件进行注释(Xieetal.,2011),然后使用R包ClusterProfile基于超几何检验进行GO和KEGG富集分析,多重校正方法为BH法,由R包ggplot2进行可视化,富集注释条目均含3个以上基因且校正后P≤0.05的用作后续分析(Wuetal.,2021)。富集通路利用hypergeometric/Fisher检验进行统计学分析。
1.5 免疫共沉淀
在3FRBP9中鉴定到的与RBP9互作的蛋白质中包含ELAV/Hu家族的另外两个成员FNE和ELAV。利用免疫共沉淀方法检测RBP9和FNE之间的相互作用。分别收集黑腹果蝇转基因品系3FRBP9,VFNE和VFNE;3FRBP9以及野生型品系成虫头部各600个,头部裂解处理同1.2节,离心后上清分别与anti-FLAG M2磁珠(Sigma,美国)或anti-V5 agarose(Sigma,美国)孵育。样品洗涤及处理同1.2节。洗脱样品使用anti-FLAG(Sigma,美国)和anti-V5(Sigma,美国)单克隆抗体进行Western blott检测分析。
2 结果
2.1 黑腹果蝇转基因品系3FRBP9和VFNE的获得
利用CRISPR/Cas9技术,分别将3×Flag和V5序列插入到RBP9和FME基因的起始密码子ATG序列下游(图1:A)。注射的供体质粒即修复质粒包含插入标签序列位置的上游和下游各约1 kb片段(图1:A)。通过筛选和遗传杂交,最终我们获得转基因黑腹果蝇纯合个体3FRBP9,VFNE和VFNE;3FRBP9。使用anti-V5和anti-FLAG抗体,经Western blot检测融合蛋白V5-FNE(约39 kD)和3×FLAG-RBP9(约60 kD)分别在3个转基因黑腹果蝇品系中能够正确表达(图1:B)。

图1 黑腹果蝇转基因品系3FRBP9(3Fr),VFNE(Vf)和VFNE;3FRBP9(Vf-3Fr)构建示意图(A)及Western blot检测融合蛋白(V5-FNE和FLAG-RBP9)的表达(B)Fig.1 Schematic diagram of construction of transgenic Drosophila melanogaster strains 3FRBP9 (3Fr),VFNE (Vf) and VFNE;3FRBP9 (Vf-3Fr) (A) and the expression of fusion proteins (V5-FNE and FLAG-RBP9) detected by Western blot (B)WT:野生型品系Wild-type strain (Bloomington no.5905).数字①-⑧为扩增片段引物编号(见表1),箭头标示引物方向。Numbers ①-⑧ are the primer numbers (see Table 1) of the amplified fragments.The arrows indicate the primer direction.
2.2 转基因品系3FRBP9成虫头部RBP9免疫沉淀产物质谱
利用免疫沉淀和Western blot方法,在转基因品系3FRBP9成虫头部洗脱的两个样品E1和E2均能检测到RBP9蛋白(图2:A);同时银染结果显示,与野生型品系相比,转基因品系3FRBP9两个洗脱样品中均有明显差异条带(图2:B,红色箭头标示)。
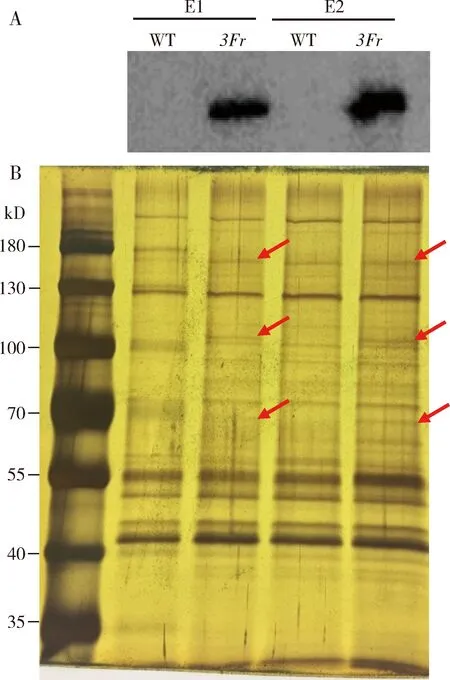
图2 黑腹果蝇野生型品系(WT)和转基因品系3FRBP9(3Fr)成虫头部中RBP9表达的Western blot(A)和银染(B)检测Fig.2 Expression of RBP9 protein in the adult heads of the wild-type strain (WT) and transgenic strain 3FRBP9 (3Fr) of Drosophila melanogaster by Western blot (A) and silver staining (B)E1,E2:洗脱样品Eluted samples.红色箭头标示的是差异蛋白。The red arrows represent the differential proteins.
3FRBP9和野生型品系头部中RBP9蛋白质谱分析结果显示,样品质谱产生的二级谱图数分别为7 684和3 449个,解析的二级谱图数分别为1 743和391个,通过ProteinPilot软件检索分析,在3FRBP9和野生型品系中分别鉴定到可能与RBP9相互作用的蛋白质分别为220和43个。其中在野生型品系和3FRBP9中鉴定到特异与RBP9相互作用的蛋白分别有13和190个,共有的蛋白质有30个。
2.3 与RBP9互作的190个蛋白质的功能注释
GO分析结果表明(图3),190个蛋白质编码基因被注释为三大类:生物学过程(biological process)、细胞组分(cellular component)和分子功能(molecular function),主要包括生物学过程类别中的染色质组装或拆卸(chromatin assembly or disassembly)、蛋白质-DNA复合物亚基组装(protein-DNA complex subunit organization)和核小体组装(nucleosome organization);细胞组分中的染色质(chromatin)、核小体(nucleosome)和DNA包装复合物(DNA packaging complex);分子功能中的蛋白质二聚化活性(protein dimerization activity)、核糖体结构成分(structural constituent of ribosome)和含蛋白质复合物结合(protein-containing complex binding)等。同时,KEGG富集分析发现,这190个编码基因主要富集在核糖体(ribosome)、碳代谢(carbon metabolism)、柠檬酸循环(三羧酸循环)[citrate cycle(TCA cycle)]和氨基酸生物合成(biosynthesis of amino acids)等生物通路(图4)。

图4 黑腹果蝇转基因品系3FRBP9成虫头部中鉴定到的与RBP9互作的190个蛋白编码基因的KEGG通路分析Fig.4 KEGG pathway analyses of genes encoding 190 proteins interacting with RBP9 in the adult head of transgenic Drosophila melanogaster strain 3FRBP9
2.4 RBP9和FNE的相互作用
免疫共沉淀实验结果显示,利用anti-V5 agarose,RBP9能够从转基因黑腹果蝇VFNE;3FRBP9成虫头部中被拉下,且能被anti-V5抗体正确检测(图5)。同时,利用anti-FLAG磁珠,FNE能够从VFNE;3FRBP9成虫头部中被拉下,且能被anti-FNE抗体正确检测(图5)。以上结果表明RBP9在黑腹果蝇神经系统中与FNE存在直接或间接的相互作用。
3 讨论
在本研究中,我们利用免疫印迹和蛋白质谱方法在黑腹果蝇转基因品系3FRBP9成虫头部中鉴定了190个与RBP9相互作用的蛋白质。经GO注释和KEGG富集分析,这些蛋白质编码基因主要涉及染色质组装或拆卸、蛋白质-DNA复合物组装、蛋白质二聚体化活性、核糖体、碳代谢、三羧酸循环和氨基酸生物合成等生物学过程或通路(图3和4),该结果表明作为RNA结合蛋白,RBP9可能在头部神经系统中的基因表达、能量代谢等过程中具有重要的调控作用。
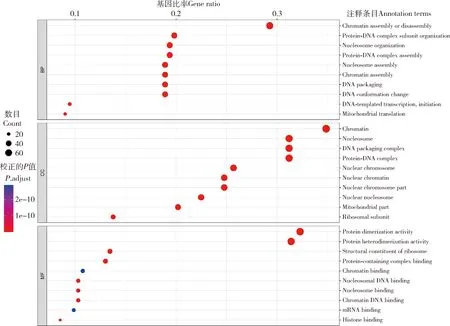
图3 黑腹果蝇转基因品系3FRBP9成虫头部中鉴定到的与RBP9互作的190个蛋白编码基因的GO功能注释Fig.3 GO functional annotation of genes encoding 190 proteins interacting with RBP9 in the adult head of transgenic Drosophila melanogaster strain 3FRBP9BP:生物学过程Biological process;CC:细胞组分Cellular component;MF:分子功能Molecular function.
在黑腹果蝇中,ELAV家族包括ELAV,FNE和RBP9。Toba和White (2008)利用酵母双杂交验证了三者在体外存在相互作用。该研究组进一步利用黑腹果蝇杂交实验发现elav和rbp9间的遗传相互作用在黑腹果蝇寿命中具有重要作用(Tobaetal.,2010)。在本研究鉴定的与RBP9互作蛋白质中发现同家族的ELAV和FNE,进而利用免疫共沉淀实验验证了RBP9和FNE之间在体内存在相互作用(图5),该结果与已报道结果相一致。由此我们推测该家族3个成员间的相互作用可能在黑腹果蝇生长发育中具有重要作用,其具体调控机制有待进一步研究。

图5 免疫共沉淀法检测黑腹果蝇转基因品系3FRBP9成虫头部中RBP9和FNE的相互作用Fig.5 Interaction between RBP9 and FNE in the adult head of transgenic Drosophila melanogaster strain 3FRBP9 detected by immune coprecipitation method1:野生型品系Wild-type strain (Bloomington no.5905);2:VFNE;3:VFNE;3FRBP9;4:3FRBP9.4个品系头部蛋白样品分别与anti-V5 agarose和anti-FLAG M2 磁珠孵育;Input和immune coprecipitation样品分别使用anti-V5和anti-FLAG抗体检测。Protein samples from the head of four strains were incubated with anti-V5 agarose and anti-FLAG M2 beads,respectively.Input and immune coprecipitation samples were detected using anti-V5 and anti-FLAG antibody,respectively.
RBP9分布于细胞质和细胞核中,能够在不同水平上调控基因的表达(Kim and Baker,1993;Kim-Haetal.,1999)。在我们鉴定与RBP9相互作用的蛋白中,包含了与基因表达相关的蛋白,如线粒体BSF(bicoid stability factor),Pep(protein on ecdysone puffs),PS(pasilla)和qkr58E-2(quaking related 58E-2)等。BSF能够结合bicoid基因的3′UTR以维持其稳定性(Braticetal.,2011)。PS,Pep和qkr58E-2均具有RNA结合活性,并且通过剪接体参与mRNA剪接调控(Allende and Allende,1995;Martinetal.,2014;Liuetal.,2018)。研究发现RBP9与poly U序列结合,调节目的RNA的剪接和稳定性(Parketal.,1998;Kimetal.,2010)。最近研究报道通过抑制远端多聚腺苷酸化位点的使用,RBP9可以诱导3′UTR延长(Weietal.,2020)。本研究质谱鉴定的蛋白中包含了多聚腺苷酸结合蛋白[poly(A) binding protein,PABP]和DmelCG13928,前者能够与靶标mRNA 3′UTR结合,进而调控其亚细胞定位(Kamiyamaetal.,2020);后者可调控靶标mRNA poly(A)的长度(Khanetal.,2015)。由此我们推测RBP9在3′多聚腺苷化形成中具有重要调控作用。另外,我们研究鉴定的与RBP9相互作用的蛋白还包括与翻译相关的蛋白,如真核翻译起始因子4B(eIF4B)、真核翻译延伸因子1 α1(eEF1 α1)和真核翻译延伸因子1γ(eEF1γ)。这些发现表明RBP9可能与这些蛋白质相互作用,共同参与靶标基因的翻译过程。
编码RBP9蛋白基因缺失会导致黑腹果蝇运动活动减少,血脑屏障异常,寿命缩短,但导致这些缺陷的机制尚不清楚(Kimetal.,2010)。神经系统复杂而有序的活动需要消耗大量的能量(Camandola and Mattson,2017;Rittschof and Schirmeier,2018)。神经元中糖原的异常积累导致小鼠和黑腹果蝇神经元死亡、运动缺陷和寿命缩短(Duranetal.,2012;Yamadaetal.,2019)。我们的研究鉴定了与RBBP9相互作用的糖原代谢相关、TCA循环的线粒体蛋白,如糖原合成酶、脂肪酸合成酶1、CG9485、ATPsynbeta、琥珀酰辅酶A合成酶α亚基1、异柠檬酸脱氢酶3a等。这些蛋白质在黑腹果蝇的能量代谢中发挥重要作用(Garridoetal.,2015;Quanetal.,2017)。有研究发现RBP9与同家族的ELAV和FNE共同参与黑腹果蝇神经突触生长。我们鉴定了Eps-15(epidermal growth factor receptor pathway substrate clone 15)和Chc(clathrin heavy chain),这两个蛋白在突触囊泡循环中具有重要作用(Kohetal.,2007;Kasprowiczetal.,2008)。这些蛋白的鉴定表明,RBP9可能调控能量代谢途径和神经突触发生中重要基因的表达或者与其中重要蛋白质相互作用,进而影响神经系统的活动。
根据本研究结果,我们推测,在黑腹果蝇神经系统中RBP9能够与多种蛋白质以直接或间接的方式相互作用,在基因表达多个水平、能量代谢和神经突触发生的调控中起着重要作用,该研究为RBP9在黑腹果蝇神经系统发育中的生物学功能提供了重要的实验依据。
猜你喜欢
学苑创造·A版(2023年10期)2023-11-04 13:14:04
贵州畜牧兽医(2023年3期)2023-06-29 07:07:28
农技服务(2023年2期)2023-03-15 00:43:08
大自然探索(2023年11期)2023-03-01 09:04:36
学苑创造·A版(2022年3期)2022-03-29 23:32:16
基层中医药(2021年1期)2021-07-22 06:56:44
军事文摘(2020年22期)2021-01-04 02:16:38
园林科技(2020年2期)2020-01-18 03:28:18
学苑创造·A版(2019年6期)2019-07-11 01:07:39
天津医科大学学报(2015年2期)2015-12-22 09:24:34
