
三峡库区马尾松人工林土壤酶活性和微生物生物量对氮添加的季节性响应
2022-01-16 05:35:32王丽君程瑞梅肖文发沈雅飞曾立雄孙鹏飞
生态学报 2021年24期
王丽君,程瑞梅,2,*,肖文发,2,沈雅飞,2,曾立雄,2,杨 邵 ,孙鹏飞,陈 天
1 中国林业科学研究院森林生态环境与保护研究所,国家林业和草原局森林生态环境重点实验室,北京 100091 2 南京林业大学南方现代林业协同创新中心,南京 210037
在20世纪里,粮食和能源生产以及其他人类活动使大气中的氮沉降增加了3到5倍,氮沉降增加已成为全球变化的重要现象之一。目前我国已成为第三大氮沉降区,年总沉降均值约为21.1 kg hm-2a-1,且仍将继续增长[1]。氮沉降的增加改变了土壤生态系统物质输入,对植物净初级生产力的提高和土壤固碳具有一定的贡献,但同时也导致植物多样性的丧失,加速了土壤酸化[2-3],并对参与碳氮磷循环关键过程的酶活性产生显著影响[4-5]。
目前,关于氮添加对酶活性的影响研究多集中在北方和温带森林,且结果存在差异[6- 11]。研究表明,氮添加显著提高了温带森林生态系统和农田生态系统氮循环水解酶N-乙酰氨基葡萄糖苷酶、纤维二糖水解酶和酸性磷酸酶活性,但是对于热带和沙漠生态系统则影响不显著[6]。吕来新等[7]研究发现氮添加提高了温带红松林土壤N-乙酰氨基葡萄糖苷酶活性,而对酸性磷酸酶、β- 1- 4葡萄糖苷酶活性无显著影响。多数研究表明,氮沉降通过抑制与氧化酶合成和分泌有关的真菌(如白腐真菌、丛枝菌根真菌)的活性,从而使其活性也下降[8- 10]。但张艺等[4]、周嘉聪等[11]发现氮添加显著促进了土壤氧化酶活性,且低氮水平(50 kg hm-2a-1)添加比高氮水平(150 kg hm-2a-1)添加的促进作用更为明显。研究表明土壤养分有效性可能是各地研究结果差异的主要原因[12],整合分析也表明不同生态系统类型,林分类型、氮肥类型、施肥速率和时间都对土壤酶活性具有影响[6,13]。相比于温带森林,亚热带森林土壤的有效氮相对富集[14],养分循环较快,对氮沉降的响应可能更加剧烈[11]。因此,研究氮沉降增加对亚热带森林土壤酶活性和微生物生物量的影响十分必要。
三峡库区作为我国重点敏感生态区和长江中下游的生态屏障,大气氮沉降通量已达到约30 kg hm-2a-1,超过了我国氮沉降平均值,属于高氮沉降地区[15],库区有大面积的马尾松(PinusmassonianaLamb.)人工林[16]。氮沉降的持续增加对土壤微生物的扰动势必会影响土壤的养分循环,而目前有关大气氮沉降增加对三峡库区马尾松人工林土壤酶活性和微生物生物量的影响研究鲜有报道,影响机制也尚未明晰。基于此,本研究以亚热带三峡库区马尾松人工林为对象,研究土壤酶活性、微生物生物量和养分含量对氮添加的初期响应规律,为预测该地区在大气氮沉降持续增加背景下森林土壤养分循环的响应提供依据。
1 研究区域与研究方法
1.1 研究区概况
研究区域位于湖北省秭归县茅坪镇泗溪村,地处于30°46′N,110°55′E,平均海拔825 m,平均坡度20°,属于亚热带大陆性季风气候,四季分明,年平均气温14—22℃,年平均降水量1400 mm,降水集中于每年的4—9月。研究区内马尾松林为20世纪80年代飞播造林,平均树龄40 a,平均树高16.96 m,平均胸径18.3 cm,平均密度675株/hm2,样地内林木分布均匀。土壤类型主要有黄棕壤、紫色土等,土壤深度约40 cm。植被以天然植被为主,林下伴生灌木植物主要有:茶(Camelliasinensis(L.) O.Kuntze)、细齿叶柃(EuryanitidaKorthals)、紫金牛(Ardisiajaponica(Thunb) Blume)、宜昌荚蒾(ViburnumerosumThunb.)等;草本植物有:黑足鳞毛蕨(DryopterisfuscipesC.Chr.)、鱼腥草(HouttuyniacordataThunb.)和千里光(SenecioscandensBuch.Ham.ex D.Don)等[17]。
1.2 试验设计
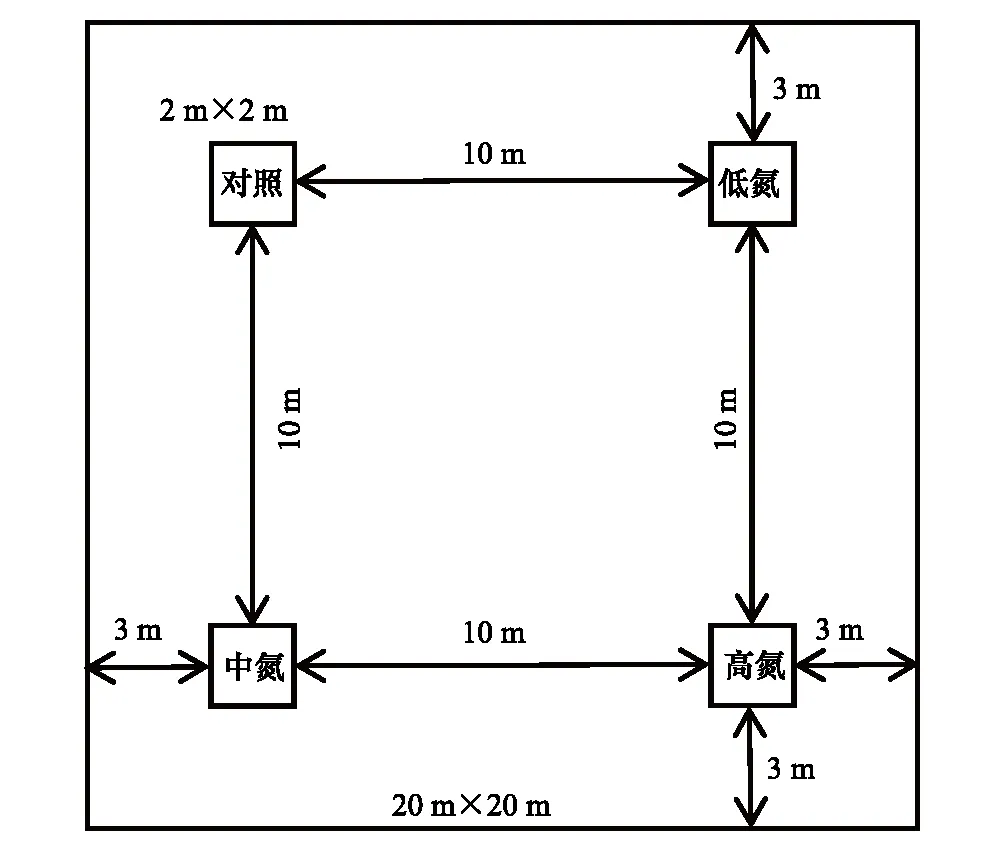
图1 氮添加实验样方示意图Fig.1 Schematic diagram of nitrogen addition experiment
以马尾松人工林为研究对象,根据当地大气氮沉降背景值(30 kg hm-2a-1)[16],通过定位氮添加的方式,设置4个不同浓度梯度的氮处理,对照处理N对照(0 kg hm-2a-1),低氮处理N低( 30 kg hm-2a-1),中氮处理N中(60 kg hm-2a-1),高氮处理N高(90 kg hm-2a-1)。在马尾松人工林内设置3块20 m×20 m的样地,在每块样地内设置4个2 m×2 m的样方,样方之间设10 m缓冲带(具体样地设置见图1),样方边缘用PVC板进行隔离,PVC板插入土壤深度为20 cm,共计12个样方。将NH4NO3的年施用量平均分成12 等份,从2019 年2月起,每月初进行定量氮添加处理:将NH4NO3溶解于2 L水中,用喷雾器在样方内均匀喷洒,对照组则喷洒相同量的清水[18-19]。
1.3 土壤样品采集分析
分别在氮添加 3(2019年5月)、6(2019年8月)、9(2019年11月)个月后,在下次氮添加之前,在每个样方内随机选择5点,用土钻(内径为5 cm)采集0—20 cm的土壤,混合均匀后放入保温箱内带回实验室。一部分过2 mm筛后置4℃冰箱里用来测定土壤酶活性、微生物生物量碳、微生物生物量氮和微生物生物量磷,一部分室内风干后过2 mm和0.149 mm筛后用于土壤pH、有机碳、全氮、全磷的测定。
本研究共测定了3种水解酶活性,分别是β- 1- 4葡萄糖苷酶(BG)、N-乙酰氨基葡萄糖苷酶(NAG)、酸性磷酸酶(AP),三者分别在土壤多糖类物质的水解过程和氨基酸物质的降解过程及有机磷的分解过程中起到了重要促进作用,是土壤碳、氮、磷转化的重要参与者[7],2种氧化酶活性,多酚氧化酶(PPO)和过氧化物酶(POD),二者是土壤氧化还原反应的主要参与者,直接制约土壤有机质的分解[9]。土壤酶活性测定参照Saiya-Cork等[20]的方法,用伞形酮(MUB)作为底物标示水解酶BG、AP、NAG活性,用L-二羟苯丙氨酸(DOPA)为底物标示氧化酶PPO、POD活性。用多功能酶标仪测定荧光度(水解酶)或吸光度(氧化酶)。采用氯仿熏蒸-K2SO4浸提法测定土壤微生物生物量碳(MBC)、微生物生物量氮(MBN),氯仿熏蒸-NaHCO3浸提法测定微生物生物量磷(MBP),电极法测定土壤pH值 (水土比2.5∶1.0,pH211酸度计)。高温外热重铬酸钾氧化-容重法测定土壤有机碳(SOC),凯氏定氮法测定全氮(TN),利用钼锑抗比色法测定全磷(TP),烘干法测定土壤含水量(SMC)。
1.4 数据分析
采用Excel 2016统计处理数据,用SPSS 24.0软件中的单因素方差分析(one-way ANONA)的最小显著差法(LSD)检验各项指标之间的差异显著性(P<0.05);用双因素方差分析(two-way ANOVA)分析氮添加和季节是否存在交互作用,用Pearson相关系数和逐步回归分析探索土壤酶活性与理化性质、微生物生物量之间的关系;用Canoco 5.0 软件,以5种酶活性为响应变量,以土壤理化性质及微生物生物量为解释变量做冗余分析(RDA)。绘图由Origin 9.1软件完成 。
2 结果分析
2.1 不同水平氮添加对土壤理化性质的季节性影响

图2 不同水平氮添加对土壤理化性质的影响Fig.2 Effects of nitrogen addition at different levels on soil physical and chemical properties不同大写字母表示同一氮水平不同月份间差异显著,不同小写字母表示同一月份不同氮水平间差异显著(P<0.05);图中数据为平均值±标准差(n=3);**表示P<0.01,ns表示P>0.05
土壤含水量受季节的显著影响(图2),秋季样地土壤含水量总体上显著低于春季和夏季(P<0.05),这与当地降水主要集中于每年的4—9月有关。土壤pH值和全磷含量随着氮添加水平的提高先增加后下降,整体呈现N低>N对照>N中>N高,夏季>秋季>春季的特征。土壤有机碳在春季随着氮添加水平的提高持续增加,在夏季和秋季则先下降后增加,全氮含量在各个季节对氮添加的响应均表现为先下降后上升,与对照相比,N高处理使得土壤有机碳、全氮含量均增加,秋季含量略高于春夏两季,但差异不显著(P>0.05)。整体来看,在短期氮添加背景下,季节、施氮及其交互作用对土壤pH值、有机碳、全氮、全磷含量影响不显著(P>0.05),但低氮(30 kg hm-2a-1)水平添加整体上增加了土壤pH值和全磷含量,高氮(90 kg hm-2a-1)水平添加增加了土壤有机质、全氮含量,降低了土壤pH值和全磷含量。
2.2 不同水平氮添加对土壤酶活性的季节性影响
2.2.1土壤水解酶活性变化
BG活性受季节(P<0.01)和施氮(P<0.05)的显著影响(图3),呈现秋季>夏季>春季的特征,春季N低和N中处理均提高了BG活性, N高处理降低了BG活性。夏季和秋季,N中处理均使得BG活性增加,而N低和N高处理使得BG活性下降。NAG的活性受季节的显著影响(P<0.01),呈现秋季>春季>夏季的特征,且各季节均表现出N中处理使得NAG活性增加,N低和N高处理使得NAG活性下降。AP活性受季节(P<0.01)和施氮(P<0.05)的显著影响,呈现秋季>春季>夏季的特征,各季节土壤AP活性在N低处理下均有所下降,在N中和N高处理下均增加。
总体来看,土壤水解酶活性存在显著的季节差异,即秋季显著高于春季和夏季(P<0.05)。各季节土壤酶活性对不同水平氮添加处理的响应不一致,低氮水平(30 kg hm-2a-1)的添加使得土壤NAG、AP活性均下降,高氮水平(90 kg hm-2a-1)的添加提高了各季节AP活性,抑制了BG和NAG的活性,而中氮水平(60 kg hm-2a-1)的氮添加处理使得各季节土壤BG、NAG、AP活性均增加。
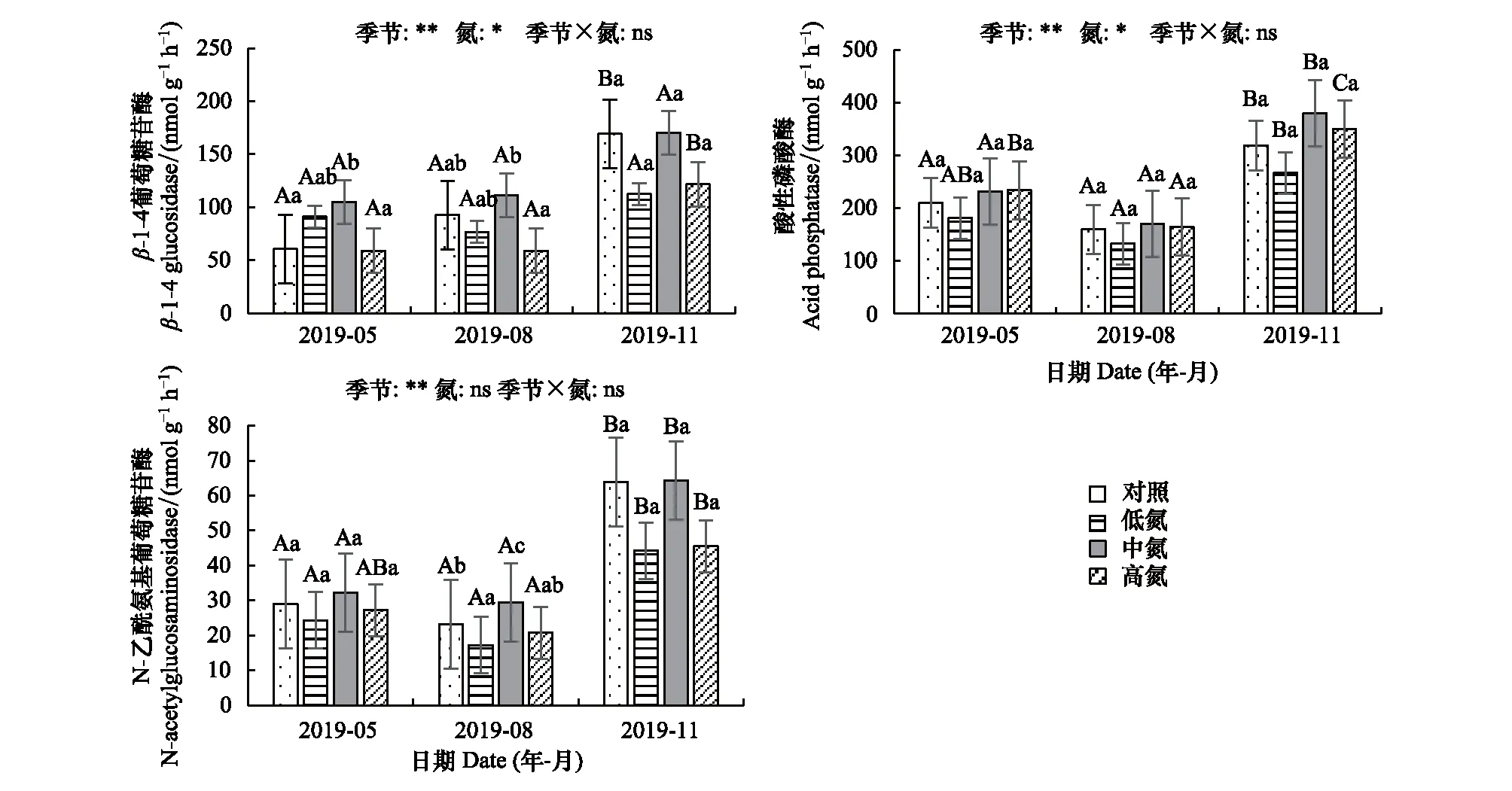
图3 不同水平氮添加对土壤水解酶活性的影响Fig.3 Effects of nitrogen addition at different levels on soil hydrolytic enzyme activities 不同大写字母表示同一氮水平不同月份间差异显著,不同小写字母表示同一月份不同氮水平间差异显著(P<0.05);图中数据为平均值±标准差(n=3);*,**分别表示P<0.05和P<0.01,ns表示P>0.05
2.2.2土壤氧化酶活性变化
各季节PPO活性均呈现随氮添加水平的提高先上升后下降的趋势(图4)。其中,春季N低水平处理使得PPO活性增加到最大,夏季和秋季N中水平处理使得土壤PPO活性增加到最大,与N对照相比,PPO活性分别增加了36.0%和43.0%。春季和夏季N中处理使得POD活性增加,N低和N高处理则使得土壤POD活性下降,秋季各水平氮添加处理均使得POD活性上升,且POD活性随氮添加水平的提高而增加。整体来看,在短期氮添加背景下,施氮对PPO和POD活性的影响没有达到显著水平(P>0.05),但不同水平氮添加处理均提高了各季节土壤PPO活性,表现为随氮添加水平的提高先上升后下降,中氮(60 kg hm-2a-1)水平的氮添加处理使得各季节土壤POD活性均增加。PPO和POD活性均呈春季>夏季>秋季的特征,其中PPO活性受季节的显著影响差异较大(P<0.05),而POD活性季节间差异不显著(P>0.05)。

图4 不同水平氮添加对土壤氧化酶活性的影响Fig.4 Effects of nitrogen addition at different levels on soil oxidase enzyme activities **表示P<0.01
2.3 不同水平氮添加对土壤微生物生物量的季节性影响
土壤MBC、MBN含量对氮添加的季节响应表现一致(图5)。春季,各水平氮添加处理均使得MBC、MBN含量增加,且随氮添加水平的提高呈现先增加后下降的趋势,其中N中处理增加最为显著,分别达到30.5%、37.7%。夏季,N低和N中处理使得MBC、MBN含量分别下降39.3%、34.7%和20.9%、27.3%,而N高处理使得MBC、MBN含量增加。秋季,N低处理使得MBC、MBN含量下降,而N中和N高处理使得其含量略有增加,变化均不显著(P>0.05)。各季节土壤MBP含量对氮添加的响应一致,均表现为N低和N中处理使得MBP含量显著下降(P<0.05),N高处理使得MBP含量上升。

图5 不同水平氮添加对土壤微生物生物量的影响Fig.5 Effects of nitrogen addition at different levels on soil microbial biomass**表示P<0.01
整体来看,土壤MBC、MBN含量受季节的显著影响(P<0.01),分别呈现出秋季>春季>夏季,夏季>春季>秋季的特征,MBP含量呈现秋季>夏季>春季的特征,氮添加整体上没有改变这种季节差异,但高氮(90 kg hm-2a-1)水平的添加处理使得各季节土壤微生物生物量碳、氮、磷的含量都上升。
2.4 土壤酶活性与微生物生物量及环境因子的关系
相关性分析可知(表1),BG、NAG活性均与全氮(P<0.05)、MBC(P<0.01)和MBP(P<0.05,P<0.01)存在显著正相关,与MBN(P<0.05)和土壤含水量(P<0.01)存在显著负相关。AP活性与MBC、MBP存在极显著正相关(P<0.01),与MBN和土壤含水量存在极显著负相关(P<0.01)。PPO活性与MBN存在显著正相关(P<0.05),POD活性与MBN(P<0.05)和全磷(P<0.01)存在显著正相关。除了POD外,其余4种酶活性均与季节存在极显著相关性(P<0.01),说明季节变化可能是酶活性变化的主要因素之一。此外,土壤全磷含量与氮添加水平存在显著负相关(P<0.05)。整体来看,土壤水解酶活性与土壤含水量,全氮、微生物生物量碳氮磷含量相关性较高,氧化酶活性和微生物生物量氮和全磷含量相关性较高。
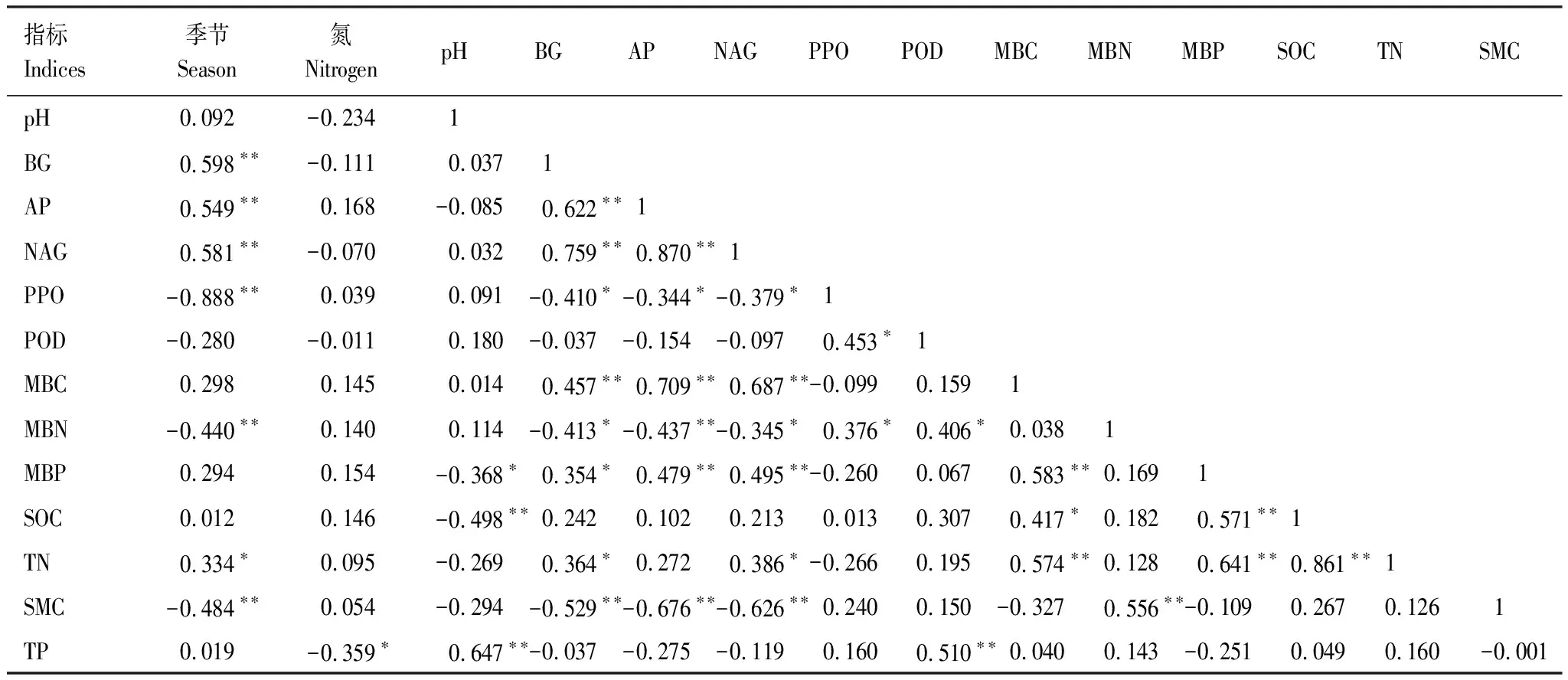
表1 土壤酶活性、微生物生物量和土壤环境因子的相关性分析
以5种酶活性为响应变量,以土壤理化性质及微生物生物量为解释变量进行逐步回归分析和冗余分析(表2,图6),可以看出,土壤含水量和全氮共同解释了BG活性46.8%的变异,MBC和土壤含水量共同解释了NAG活性65.3%的变异,以MBC、MBN、土壤含水量和全磷为主的因子共同解释了AP活性变化的84.5%,MBN和MBP共同解释了PPO活性变化的24.9%,MBN和全磷则共同解释了POD活性37.3%的变异。两轴共同解释了土壤酶活性变异的59.08%,且土壤含水量、MBC、MBN、MBP和全氮是解释度较高的5个环境因子,其中,土壤含水量与MBC分别解释了土壤酶活性变异的28.1%和26%(P<0.05)。可见氮沉降背景下,土壤酶活性的变异是多因子综合作用的结果,不同酶活性变异的主导因子不同。
3 讨论
3.1 不同水平氮添加对土壤理化性质的季节性影响


表2 土壤酶活性与微生物生物量及土壤环境因子的回归分析模型
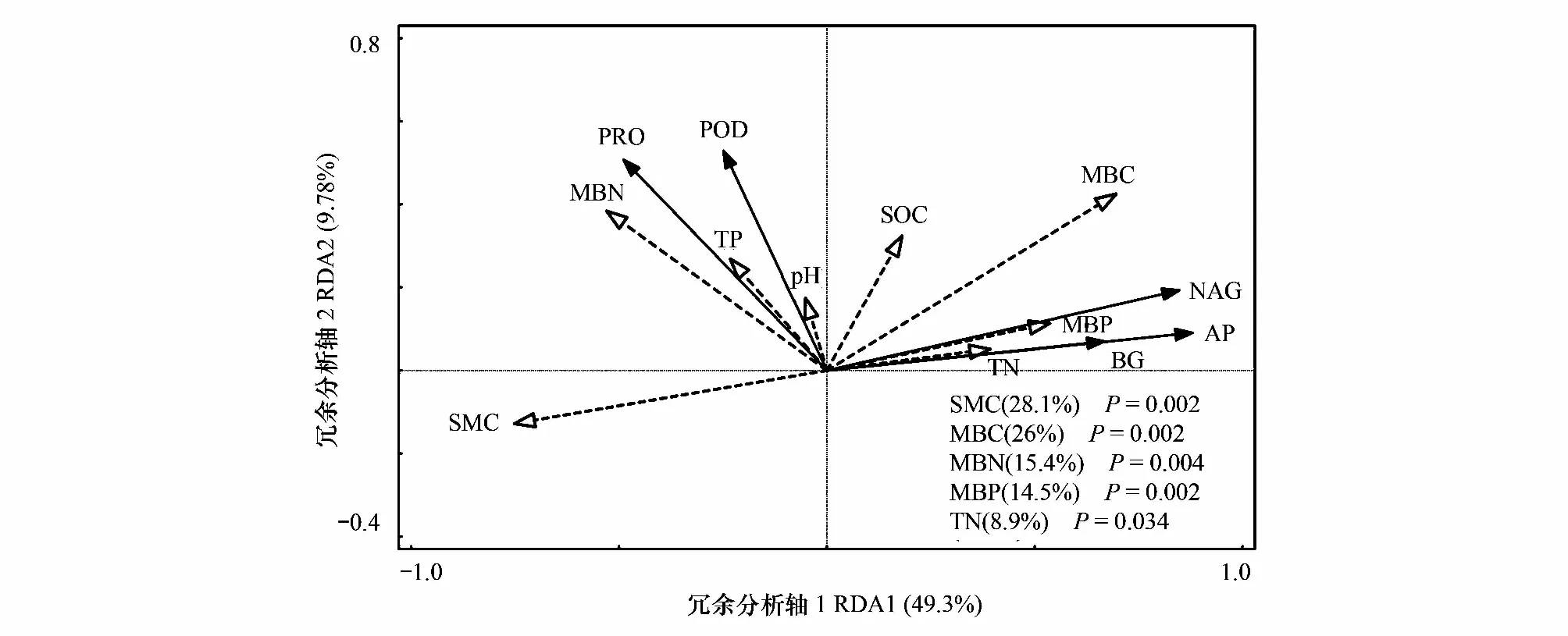
图6 土壤酶活性与微生物生物量及环境因子的冗余分析Fig.6 Redundancy analysis of soil enzyme activities and environmental factorsBG:β- 1- 4葡萄糖苷酶 β- 1- 4 glucosidase;NAG:N-乙酰氨基葡萄糖苷酶 N-acetylglucosaminosidase;AP:酸性磷酸酶 Acid phosphatase;PPO:多酚氧化酶 Polyphenol oxidase;POD:过氧化物酶 Peroxidase;MBC:微生物生物量 Microbial biomass carbon;MBN:微生物生物量氮 Microbial biomass nitrogen;MBP:微生物生物量磷 Microbial biomass phosphorus;SOC:土壤有机碳 Soil organic carbon;TN:全氮 Total nitrogen;TP:全磷 Total phosphorus;SMC:土壤含水量 Soil moisture content
3.2 不同水平氮添加对土壤酶活性的季节性影响
一般认为,土壤微生物生产酶遵循经济学原则,氮添加减少了微生物对氮的需求,转而增加对碳、磷的需求,因而使得氮水解酶NAG活性下降,碳、磷水解酶BG、AP活性升高[31]。但是,经济学原则并非适用于所有森林系统,如有研究表明,氮添加抑制了温带地区土壤中碳、氮、磷水解酶活性[32],但却显著提高了亚热带地区土壤水解酶活性[6,11,13],可见氮添加对土壤水解酶活性的影响结果因地域类型、林分的不同存在较大差异[33]。本文发现,低氮水平(30 kg hm-2a-1)和高氮水平(90 kg hm-2a-1)的添加使得BG和NAG的活性下降,而中氮水平(60 kg hm-2a-1)的氮添加处理却使得各季节土壤BG、NAG、AP活性均增加,这与陈倩妹等人的研究结果相似,可能是由于中氮水平是增加当地马尾松人工林土壤有效氮的最适浓度,在此水平下,土壤水解酶BG和NAG的活性达到最大,低于或者高于此浓度,酶活性均会下降,这有待进一步验证[34-35]。
多酚氧化酶和过氧化物酶是土壤氧化还原反应的主要参与者,直接制约土壤有机质的分解。PPO能够氧化酚类化合物中的苯环,POD能够氧化木质素大分子得到简单的酚类,并能反映土壤的腐殖化和有机化的程度[9]。多数研究中,氮添加抑制了森林土壤氧化酶的活性,主要是因为氮添加抑制了与其合成和分泌有关的真菌(如白腐真菌、丛枝菌根真菌)的活性,从而使其活性也下降[8- 10,36]。但本文发现,中氮(60 kg hm-2a-1)水平的氮添加处理使得各季节土壤POD活性均增加,不同水平的氮添加处理均增加了各季节土壤PPO的活性,可见氮添加对土壤氧化酶活性的影响并不都是负效应。事实上,白腐真菌并不是唯一一种和多酚氧化酶活性相关的生物,而且其他生物对氮沉降增加的响应可能与白腐真菌差异很大,如一些软腐真菌在氮沉降增加时,多酚氧化酶活性明显提高[37- 38]。还有研究发现,环境中氮有效性的高低使得氮添加对氧化酶活性的影响不同,在不同林型中甚至得到相反的结果,如研究发现,施氮1年后,在黑橡和白橡混交林中,多酚氧化酶活性降低,而在糖槭和红橡及糖槭和椴木混交林中,氮沉降增加则使多酚氧化酶活性增加[37]。由于土壤氧化酶在腐殖化过程中扮演重要作用,其活性的增加表明氮沉降可能会加速当地马尾松人工林土壤腐殖质的形成[34]。
范艳春等[39]研究发现,土壤酶活性呈现明显的季节动态。本研究中,土壤水解酶BG、AP、NAG和氧化酶PPO活性均存在显著的季节差异。秋季土壤水解酶活性显著高于春季和夏季(P<0.05),PPO和POD活性均呈现春季>夏季>秋季的特征,而氮添加对森林土壤酶活性的时间分异规律没有显著影响,这与前人研究结果一致[4]。
3.3 不同水平氮添加对土壤微生物生物量的季节性影响
土壤微生物生物量对推动土壤物质转换和能量流动起着重要作用,是衡量土壤微生物对养分循环影响的重要指标,也是土壤质量评价的有效指标[40]。研究表明,超过微生物耐受范围的氮沉降必然导致土壤微生物生物量下降[41],而本研究发现高氮(90 kg hm-2a-1)水平的添加处理使得各季节土壤微生物生物量碳、氮、磷的含量都上升,可能由于短期高氮水平的添加试验并没有使土壤中的氮超过微生物耐受范围,达到饱和状态,土壤中的微生物能够及时将输入的氮吸收利用[42]。众多研究表明,土壤微生物生物量含量存在明显的月动态变化特征[43-44],本研究发现,土壤微生物生物量碳、氮、磷含量存在显著的季节差异,微生物生物量氮含量在春夏季偏高,秋季较低,而微生物生物量碳、磷含量均在秋季最高,氮添加没有改变这种季节差异。这与前人的研究结果有一定差别,如宋蕾等[43]研究发现微生物生物量碳、氮含量的最大值在夏季,而周世兴等[45]研究得出微生物生物量碳、氮含量在秋季最高。这些结论差异与土壤温湿度、林分类型、地域气候类型等环境因子综合作用有关[46],有研究发现不同海拔梯度带土壤微生物生物量氮的季节变化与其对应的土壤湿度显著相关,土壤湿度是调控森林土壤微生物生物量氮季节变异的重要生态因子[47],本研究中秋季土壤含水量显著低于春夏两季(图2),因而造成土壤微生物生物量氮含量也偏低。也有研究发现地处高纬度的森林,春季和秋季温度较低,土壤微生物主要受到温度的限制,生物量低;夏季气温升高,适合微生物生长,因而土壤微生物生物量增加。而亚热带和热带地区土壤微生物生物量受温度影响较小,主要与土壤养分含量有关[44]。秋季植物生长缓慢,对土壤养分的吸收相对较少,土壤中积累的碳氮含量较高(图2),微生物有充足的碳源、氮源用于生长,因而秋季微生物生物量较多。
3.4 氮添加背景下土壤酶活性和微生物生物量及土壤环境因子的关系
周嘉聪等[11]研究发现,可溶性有机碳是驱动土壤酶活性的重要环境因子,本研究发现土壤酶活性和季节、土壤含水量、养分含量及微生物生物量碳氮磷均存在显著的相关性,其活性变化是多因子综合作用的结果,不同酶活性变异的主导因子不同。其中,土壤含水量、微生物生物量碳、氮、磷和全氮是解释度较高的5个环境因子(表1、表2,图6)。研究表明,环境因子综合作用会掩盖了单一环境因子对土壤养分、酶活性和微生物的作用[46],不同生态系统类型、氮肥类型、施肥速率和施肥试验时间都对土壤酶活性和微生物群落组成具有影响,土壤酶化学计量比与土壤养分化学计量比也可能存在动态平衡关系[6,13]。多数研究中,在同一氮水平处理下,水解酶和氧化酶活性变化不同[7- 8,10],而在本研究中,中氮水平(60 kg hm-2a-1)添加在各个季节都提高了土壤水解酶和氧化酶活性,且使大多数酶活性达到最大值,由此可提出进一步的研究方向,目前的氮添加速率和中氮水平的添加是否是增加当地马尾松人工林土壤有效氮和微生物活性的最适浓度?这有待结合区域环境条件,通过控制变量进行长期实验来进一步探讨。
4 结论
(1)氮添加对土壤pH值、有机碳、全氮和全磷含量没有形成显著的季节间差异,但高氮(90 kg hm-2a-1)水平添加增加了土壤有机碳、全氮含量和酸性磷酸酶活性,降低了土壤pH值和全磷含量,且全磷与氮添加水平显著负相关(P<0.05),这预测了氮沉降的持续增加可能会导致当地马尾松人工林土壤酸化,促进磷酸盐的释放,引起磷限制。
(2)各季节土壤酶活性、微生物生物量对不同水平氮添加处理的响应不一致,但不同水平氮添加处理均提高了土壤PPO活性,且中氮水平(60 kg hm-2a-1)的氮添加处理使得各季节土壤BG、NAG、AP、PPO、POD活性均增加,高氮(90 kg hm-2a-1)水平的添加处理提高了各季节土壤微生物生物量碳、氮、磷含量,抑制了BG、NAG活性。水解酶活性下降,氧化酶活性的增加表明氮沉降会加速当地马尾松人工林土壤腐殖质的形成,增加有机碳的积累。
(3)土壤酶活性和微生物生物量存在显著的季节差异,秋季水解酶活性和微生物生物量碳、磷含量显著高于春夏两季,而氧化酶活性和微生物生物量氮含量则是春夏季较高,氮添加对这种季节分异规律没有显著影响。
(4)氮沉降增加背景下,土壤酶活性和季节、土壤含水量、养分含量及微生物生物量碳氮磷含量存在显著的相关性,其活性变化是多因子综合作用的结果,不同酶活性变异的主导因子不同。冗余分析表明土壤含水量、微生物生物量碳、氮、磷和全氮是驱动土壤酶活性的主要环境因子。
猜你喜欢
生物加工过程(2023年5期)2023-10-10 06:41:36
生物信息学(2022年1期)2022-04-01 08:56:50
昆明医科大学学报(2020年12期)2021-01-26 00:44:12
石油化工应用(2018年3期)2018-03-24 14:54:36
现代农业(2016年3期)2016-04-14 12:35:28
中国粮油学报(2016年5期)2016-01-23 02:44:40
土壤与作物(2015年3期)2015-12-08 00:47:01
茶叶通讯(2014年2期)2014-02-27 07:55:39
食品科学(2013年15期)2013-03-11 18:25:38
郑州大学学报(理学版)(2012年4期)2012-03-25 13:59:10
