
黄土区不同恢复年限草地群落生物量及根冠比对氮添加的响应
2022-01-17 08:47:56樊勇明温仲明杨玉婷姜艳敏
生态学报 2021年24期
樊勇明,李 伟,,温仲明,,*,郭 倩,刘 晶,杨 雪,郑 诚,杨玉婷 ,姜艳敏,张 博
1 西北农林科技大学草业与草原学院, 杨陵 712100 2 中国科学院教育部水土保持与生态环境研究中心,杨陵 712100
自工业革命以来,随着工业用氮的增加和农业生产中氮肥的大量使用,大气中氮含量迅速增加,导致了氮沉降在世界范围内的发生[1- 3]。氮素是限制陆地生态系统植物生长发育和生产力的主要因素之一[4],随着氮沉降的增加,陆地生态系统的结构和功能受到严重影响[5- 6]。草地生态系统作为陆地生态系统的重要组成部分,其生产力约占陆地总初级生产力的1/3[7],具有重要的生态系统服务功能[8]。草地生态系统中的生产力是生态系统服务功能中供给功能的主要部分[9]。
草地植物群落的生物量是生态系统初级生产力的重要组成部分。其对草地生态系统结构和功能的形成具有重要作用[10],是研究草地状况的量化指标之一,也是草地生态系统管理的重要依据[11]。草地生态系统的地上最大生物量通常近似等同于当年的净地上部分生产力。可以反映该草地的健康状况及其生产潜力[12]。为预测氮沉降对草地生态系统的影响,已发表了大量的文章,有研究认为,对于养分贫瘠的草地,一定量的氮添加能提高草地生态系统的净初级生产力[13]。但是对于成熟稳定的同类草地,氮输入增加会致使植物群落净初级生产力降低[14- 15]。前期研究广泛被接受的结论是氮添加对草地生态系统生产力的促进作用存在阈值[16],即低氮输入会提高草地群落净初级生产力,长期高氮输入不仅不会增加草地群落产量,还可能会导致净初级生产力降低。
草地群落的生物量80%以上集中在地下[17- 18],地下生物量是草地群落碳蓄积的重要组成部分,在草地生态系统碳循环中起着关键作用[19],准确测定草地群落地下生物量对预测草地生态系统与氮沉降的关系以及草地资源的管理和合理利用具有重要意义[20-21]。目前地下生物量对氮添加响应的相关研究较多,但结果存在很大差异,景明慧等[22]研究认为,氮添加对群落地下生物量没有显著影响,薄正熙等[23]认为氮添加显著增加了群落地下生物量。而陈文年等[24]研究表明随着氮添加量的增加,群落地下生物量呈先增大后减少的趋势。群落地下生物量与周围土壤环境息息相关,氮添加后环境的改变必然对植物地下部分产生影响。
植物群落根冠比不仅是植物光合作用产物分配的重要体现,也是草地生态系统碳循环的重要特征,对草地群落地下生物量和植被碳储量的估算具有重要的价值,在氮沉降的背景下,也可用于评估植物对生态环境的适应性[25],以期实现对草地资源更好的管理和利用。目前关于氮添加对草地群落根冠比的影响已经进行了相关的研究,但结果并不一致;习励玮等[26]研究认为,群落地下生物量随着氮添加有降低的趋势,地上、总生物量及根冠比则有增加的趋势。也有研究认为,氮添加使群落地上生物量增加,而对地下生物量没有影响,根冠比随氮添加量的增加而降低[27-28]。还有研究认为,氮添加会同时促进群落地上地下生物量的增加,但对地上生物量的促进效果大于地下,根冠比增大[29]。在草地生态系统中,根冠比的精准测定,将有利于中国草原碳储量的估算,对草地生产也具有十分重要的意义。
前期对植物群落生物量和根冠比对氮添加响应的研究,主要集中在同一草地对长期施氮或不同放牧强度的草地对施氮的响应[15, 30- 34],而很少涉及不同恢复限的草地对氮添加的响应。本研究依托宁夏云雾山国家级草原自然保护区于2013a建立的长期氮添加实验平台,通过分析3个不同恢复年限天然草地群落地上生物量、地下生物量和总生物量以及根冠比对6个氮添加处理的响应,以期为该区域草地资源后续的合理利用和管理提供理论指导,并进一步为草地资源可持续利用及草地生态环境建设提供决策支持。
1 材料与方法
1.1 研究区自然概况
研究区位于宁夏固原市东北部的云雾山草原自然保护区 (106°21′—106°27′E,36°10′—36°17′N),海拔1800—2100m。该区域气候属于中温带半干旱区,年平均气温7℃,最冷月1月(平均最低气温-14℃),最热月7月(22—25℃),年降水量400—450mm,降雨季节分配不均,全年65%—75%的降水集中在7—9月,蒸发量1330—1640mm。年日照时数为2500h,无霜期112—140d。
该研究区植被主要是温带典型草原物种,其原有建群种和优势种是本氏针茅(Stipacapillata)、白莲蒿 (Artemisiasacrorum)、大针茅(Stipagrandis),伴生种类型以猪毛蒿(A.scoparia)、百里香(Thymusmongolicus)、星毛委陵菜(Potentillaacaulis)为主,样地基本概况见表1。
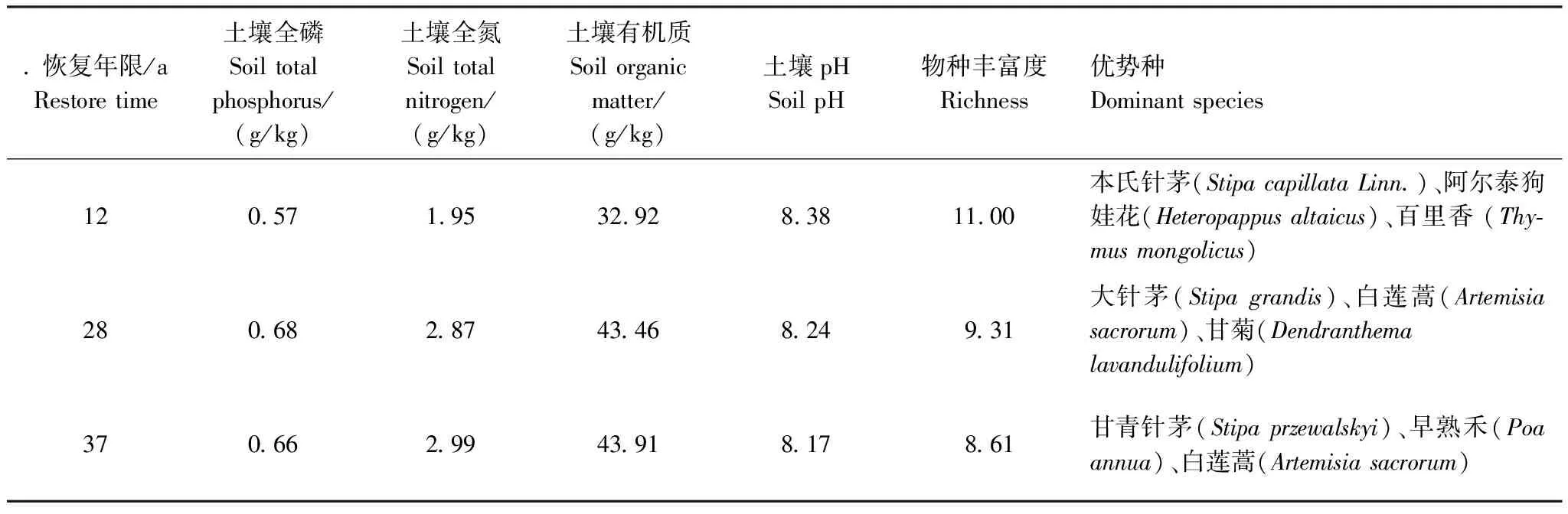
表1 实验样地的土壤和植被基本属性
1.2 实验设计
实验设计于2013年5月,选取地势、土壤类型一致,地形、地貌相似的,不同恢复年限的天然草地进行施肥试验,分别为恢复12年(2007年封禁)、恢复28年(1991年封禁)和恢复37年(1982年封禁)草地,三块草地在封禁前都是自由放牧草地,放牧强度为重度放牧,载畜率为5—8只羊单位/hm2。草地群落密度仅3—8株/m2,主要以蒿属(冷蒿、茵陈蒿等)为主,植株生长高度为10—13cm,覆盖度为30%左右,生物量不足750kg/hm2。每个恢复年限6个施肥处理,所施肥料为尿素(CO(NH2)2,施肥梯度分别为CK(0)、N1(2.34g m-2a-1)、N2(4.67g m-2a-1)、N3(9.34g m-2a-1)、N4(18.68g m-2a-1)、N5(37.35g m-2a-1)。2013年开始施肥时,1982a封育草地群落密度为54株/m2,植株生长高度为17cm,覆盖度为86%,生物量为328.44g/m2;1991年封育草地群落密度为146株/m2,植株生长高度为21cm,覆盖度为90%,生物量为281.66kg/m2;2007年封育草地群落密度为109株/m2,植株生长高度为13cm,覆盖度为83%,生物量为198.32kg/m2。根据样地大小和统计学要求,采用随机区组设计来布局,每个小区大小为4m×6m,每个处理重复6次,每个小区缓冲带为2m。为了保证试验的一致性,避免相邻小区因为施肥梯度的不同,而造成局部微环境对植物的筛选效应,从而影响群落物种多样性和生物量,试验设计中不同恢复年限的天然草地试验布局和施氮处理都一致。施肥于每年生长季初进行,即每年4月中下旬(大部分物种已返青),选择阴雨天气一次性将肥料撒播于不同的试验小区内,一直持续至今。从2013年开始施肥至2019年,7年间的平均降水量为516mm,高于多年间的平均值,7年间总体年降水量呈增加趋势。自2013年施氮实验以来,每年都进行植物群落物种数据的调查采集,同时用收获法进行地上生物量的测定,2019年采集地上生物量时,方框内的植物在前3a内未被收割过。
1.3 样品采集与测定
长期定位监测表明,研究区8月中下旬生物量达到较大值,本实验于2019年生长季高峰期(8月中下旬)进行群落调查。在每个4m×6m的实验样方内随机放置一个0.5m×0.5m的样方框,进行群落调查,为了避免边缘效应,样方距边缘的距离大于50cm,然后测定样方内物种数,以及每个物种的高度、盖度和个体数。地上生物量采用收获法测定。将每个样方内所有植物分种齐地收割后装入信封,带回实验室置于65℃烘箱烘干至恒重。称量各物种的质量,累加后计算单位面积内群落地上生物量。地上生物量收获后,用5cm直径的根钻在0—60cm各取3钻,放入根袋带回实验室。用水将植物根系冲洗干净后,于65℃烘干至恒重,称重并记录干重。
根冠比计算方法:
根冠比=地下生物量/地上生物量
近年来,有研究提出了一种新的方法来检测植物地上生物量(ANPP)对氮添加的响应,既氮响应效率(nitrogen response efficiency, NRE:ANPP对每单位氮添加的响应)和相邻氮添加水平之间的NRE差异(ΔNRE)[35-36],其计算公式如下:
其中NRE2和 Nrate2表示较高水平的氮响应效率和氮添加量,NRE1和 Nrate1表示较低水平的氮响应效率和氮添加量。
这种检测方法表明:如果NRE>0 植物生物量高于对照组,NRE<0植物生物量低于对照组,NRE=0植物生物量与对照组相同;ΔNRE≠0 表示非线性响应。ΔNRE=0 表示线性响应,具体情况如下:如果NRE2>NRE1,且ΔNRE>0,则植物ANPP对氮添加的响应为快于线性响应的非线性响应(图1),如果NRE2
1.4 数据分析
利用SPSS软件进行数据分析,利用Sigmaplot软件进行作图。不同恢复年限草地群落的地上生物量、地下生物量、总生物量和根冠比对氮添加水平的响应采用单因素方差分析(One-way ANOVA),采用Duncan′s多重比较的方法进行差异显著性检验(P=0.05)。以氮添加量为连续变量,进行回归参数估计。
2 结果与分析
2.1 不同恢复年限草地群落地上生物量对氮添加的响应
恢复年限不同,草地群落地上生物量对氮添加梯度的响应不同(图2)。随着氮梯度的增加,恢复12年时,群落生物量呈现总体上升增加的趋势,增幅范围为47%—88%;恢复28年时,呈现总体下降的趋势,降幅范围为4%—36%;恢复37年时,呈现总体上升增加的趋势,增幅范围为15%—33%。在CK至N4五个水平间,群落地上生物量均表现为恢复28年大于37年大于12年,既草地在恢复初期生产力逐渐提高,在恢复的中期达到最大,恢复后期生产力会下降,在N5水平上则表现为恢复37年(373.21g/m2)大于28a(346.54g/m2)大于12a(346.15g/m2),是由于高氮对恢复中期的地上生物量有限制作用。方差分析结果显示,在恢复12年时,N1、N2、N3的地上生物量与对照相比无显著差异(P>0.05),N4、N5的地上生物量显著高于对照和N3 (P<0.05),在恢复初期12年时,氮素是植物群落的限制因素,高氮梯度可以促进植物地上生物量的生长;在恢复28年和37年时,对照组地上生物量和不同氮添加水平间均无显著差异(P>0.05),各氮添加水平处理间也无显著差异(P>0.05),经过长期的恢复和7a的氮肥添加实验,土壤肥力得到改善,氮肥不作为植物生长的主要限制因素,因此氮添加处理对地上生物量的影响较小。

图2 氮添加对地上生物量的影响Fig.2 Effects of nitrogen addition on aboveground biomass6个氮添加水平分别为:CK(0)、N1(2.34g m-2a-1)、N2(4.67g m-2a-1)、N3(9.34g m-2a-1)、N4(18.68g m-2a-1)、N5(37.35g m-2a-1);每个处理的地上生物量表示6个重复的平均值(误差棒表示 SE)。不同小写字母表示所有处理在P<0.05水平差异显著
NRE和ΔNRE在不同恢复年限间对氮添加的响应结果(图3)所示,恢复28年时,随着氮添加水平的升高,NRE呈现上升的趋势,且ΔNRE先大于0后小于0,表明恢复28年草地群落对氮添加的响应为非线性的负相应,即氮添加对群落地上生物量产生了抑制作用,随氮添加水平升高,抑制作用越强,但每单位氮对生物量的抑制作用减弱。恢复12年和37年时,群落地上部分的NRE均大于0,随着氮添加水平的升高,NRE总体呈下降的趋势,表明每单位氮对群落地上生物量的促进作用减弱,恢复37年时ΔNRE始终小于0,恢复12年时ΔNRE先为负后为正又变为负,表明恢复12年和37年群落对氮添加为非线性的正响应,氮添加促进群落生物量增加,但随氮添加水平升高,每单位氮对生物量的促进作用减弱。

图3 地上部分NRE和ΔNRE在不同氮添加水平的变化Fig.3 Changes in aboveground NRE and ΔNRE with different ranges of N addition rate图中的点表示平均值,线段表示标准误差
2.2 不同恢复年限草地群落地下生物量和总生物量对氮添加的响应
氮添加对地下生物量和总生物量的结果(图4)如下:恢复12、28年时,对照组地下生物量和总生物量对不同氮添加水平间均无显著差异(P>0.05),恢复37年时,对照组地下生物量对不同氮添加水平间均无显著差异(P>0.05),但总生物量对照组与N4添加水平下存在显著差异(P<0.05),表明在不同恢复年限间,地下生物量对氮添加的响应较小,本研究认为这是由于氮添加于土壤,再反馈于根系需要很长时间。恢复12年和恢复37年时地上生物量和总生物量均呈现总体上升趋势,恢复28年时则相反,呈现总体下降趋势。在恢复12年时,地下生物量和总生物量最高出现于N5添加水平下,分别为对照组的1.20倍和1.21倍;在恢复37年时,地下生物量和总生物量最高出现于N4添加水平下,分别为对照组的1.73倍和2.21倍;但在恢复28年时,地下生物量和总生物量最低出现于N5添加水平下,分别为对照组的0.58倍和0.60倍;在恢复12年和37年时,氮添加对地下生物量和总生物量有促进作用,在高施氮梯度下,促进作用强于低施氮。恢复28年时则表现为相反,即对地下生物量和总生物量有抑制作用,且随氮梯度升高,抑制作用越强。这是由于恢复12年和恢复37年的优势种为禾本科牧草,对氮添加响应敏感,而恢复28年白莲蒿在群落中占比较大,导致出现这种现象。

图4 氮添加对地下生物量和总生物量的影响Fig.4 Effects of nitrogen addition on belowground biomass and total biomass
2.3 不同恢复年限草地群落根冠比对氮添加的响应
根冠比对氮添加的响应结果(图5)如下:恢复12年和恢复28年的草地群落根冠比总体有下降的趋势,而恢复37年根冠比则表现为整体上升的趋势。恢复12年时,方差分析结果显示,N1、N4、N5添加水平下的根冠比显著低于对照组(P<0.05),而N3水平下的根冠比显著高于对照和其他氮添加水平(P<0.05)。其中对照组根冠比为7.83,N1、N2、N4、N5的根冠比分别降低了55%、32%、46%、40%,而N3水平比对照增加了17%,表明中等梯度的氮添加,可以促使根冠比增大,过低或过高的氮添加会导致根冠比降低。氮添加后对地下生物量影响较小,这是由于地上生物量对氮添加的响应不同导致的。恢复28年时,对照组根冠比为3.83,N1、N2水平比对照组增加均未超过10%,N3、N4、N5与对照相比,分别降低了11%、29%、3%,表明低氮梯度促使根冠比增大,高氮梯度降低。恢复37年时,各氮添加水平均比对照增加,增加最少为N2水平,增加了58%,最多为N5水平,增加了113%,表明氮添加致使根冠比增大,氮添加梯度越大,增幅越强,恢复37年的土质疏松,优势种为禾草,氮添加会促进须根系的增加来使植物获取更多的养分。
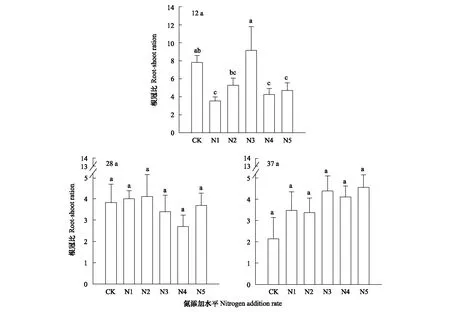
图5 氮添加对根冠比的影响Fig.5 Effects of nitrogen addition on root-shoot ratio不同小写字母表示处理间差异显著(P<0.05)
3 讨论
本研究通过对黄土高原不同恢复年限天然草地的氮添加试验,结果表明,恢复12年群落地上生物量随氮添加呈增加趋势,氮添加后生物量增加了47%—88%,且在氮水平到达N5(37.35g m-2a-1)时,与对照组呈显著相关,即氮添加未达饱和阈值水平。恢复37年,群落地上生物量增加了15%—33%,随氮添加量增加,地上生物量总体呈上升趋势,但和对照均无显著关系,恢复28年则表现为相反的情况,呈下降趋势,降低了4%—36%,和对照无显著关系。但总体上恢复28年的地上生物量高于恢复12年和37年,与以往研究结果存在差异,Bai等[15]在进行了4a的氮添加试验,结果发现地上生物量增加了98%—271%,氮饱和阈值为10.5g m-2a-1;Lan等[38]进行了10a的氮添加实验发现地上生物量的氮饱和阈值为17.5g m-2a-1,而在同一样地上连续18a的氮添加处理下,发现不同氮添加水平下群落地上生物量差异不显著[30]。景明慧等[22]在进行了7a的氮添加试验发现地上生物量增加了37%—117%,氮饱和阈值为22.4g m-2a-1;综合分析其原因,本研究认为在同样的氮添加条件下,恢复年限较短(12年)时长期氮添加会提高地上生物量,与Bai等[15]、景明慧等[22]和Lan等[38]的研究结果一致,恢复年限较长(37年)时长期氮添加会提高地上生物量,但与对照差异不显著,与王玉冰等[30]连续18a的氮添加结果相似;恢复年限处于中间状态(28年)时,虽然长期氮添加后生物量与对照无显著差异,但总体生物量呈下降趋势;以往研究很少出现这种结果,王玉冰等[30]认为初级生产力主要是由群落内优势物种的生物量及功能性状所决定,本研究中,封育12a和37a的优势物种主要为针茅属的禾本科,其根系为须根系,易于吸收养分而提高植物叶氮含量,进而通过提高植物光合速率来提高地上生物量。而Bai等[39]和杨晓霞等[40]研究也认为,植物功能群地上净初级生产力对长期氮添加的响应不同,氮添加均显著增加了禾草的地上生物量。杨倩等[41]研究认为,草地土壤性质的差异是造成地上生物量对氮添加响应有所不同的原因之一,Luo等[42]研究发现,在草地恢复过程中,随着根系生物量、根系中的C/N比、微生物C量以及土壤碳库的增加,改变了土壤原有的性质,会致使土壤净N矿化速率和N的生物有效性快速下降。植物与土壤资源有效性的互作效应将表现为负反馈,限制了植物生产力的进一步提高[43]。本研究认为这是造成封育28a优势种为白莲蒿草地群落生物量下降的主要原因。
草地群落生物量大约有80%以上集中分布在地下,在生态系统碳循环中起重要作用[17]。本试验研究样地植物以多年生为主,从实验开始到取样,已经过7a,地下生物量是7a实验处理效果的累积,可以很好的反应地下生物量对氮添加的响应。本研究表明,氮添加对不同恢复年限草地群落的地下生物量均无显著影响,但地下生物量随氮含量增加的变化趋势不同,恢复28年有下降趋势,恢复12年和37年时有上升趋势。与前人研究结果有所差异;刁励玮等[26]研究认为,施氮对群落地下生物量影响不显著,但地下生物量随着氮添加年限的增加有降低的趋势。Xu等[44]研究表明,地下生物量随氮添加量增加有降低趋势,郭璇等[45]研究认为,地下生物量有随着氮添加剂量增加而增加的趋势。这可能与草地群落的结构和恢复演替进程有关,有研究表明,群落地下生物量对氮添加响应的差异与草地群落的优势种有关,氮添加会降低杂类草的地下生物量,增加禾草的地下生物量[39,44]。恢复12年和37年草地群落以禾草为主,因此添加氮后地下生物量有上升趋势。恢复28年时,处于三个恢复期的中间状态,优势物种为菊科的白莲蒿。有研究发现,当草地生态系统植被由深根系向浅根系物种发生转变[46],长期氮添加处理对地下生物量的促进作用在处理后期转为抑制作用[26],其次氮添加对不同根系形态的响应不同[47],白莲蒿为根蘖型粗根系植物,主根不甚发达,水平根发达,早期不定根存在死亡现象,植株老龄后,主根会出现腐烂、枯死等现象[48]。本研究认为这是导致恢复28年草地添加氮后地下生物量有下降趋势的主要原因。除此之外,苏淑兰等[49]研究认为,不同管理方式对草地的地下生物量也会有影响,因此,今后在研究地下生物量时,应充分考虑草地的管理利用方式、群落间的物种组成,以及群落中根系形态的分布比例,系统化的来研究地下生物量对氮添加的响应。
根冠比反映了植物对环境的响应策略,是指示草原生态系统碳循环的关键指标[50],通过氮添加试验,来预测草地生态系统氮沉降变化时,地上、地下生物量分配具有重要意义。本研究经过7a的氮添加处理后,恢复12年草地群落地上地下生物量均增加,地上生物量增幅大于地下,根冠比下降;恢复28年,地上地下生物量均减少,地上生物量减少幅度小,根冠比下降;恢复37年和恢复12年相似,但地上生物量增幅小于地下,根冠比上升。贺星等[29]经过3a试验处理,发现氮添加处理后均使群落的根冠比下降,主要是因为植物生长明显受氮限制,景明慧等[22]认为,根冠比下降,是由于地上生物量对氮添加响应更加敏感。本研究中,经过长期恢复后,不同恢复期的群落结构和土壤理化性质均存在一定差异。恢复12年时,草地群落地上部分垂直空间分布不均,氮添加后会促进植物地上部分生长,进而提高了植物群落垂直空间的密度[51],并因此引发了光照限制,地上部分的光合产物分配增加,进而导致根冠比降低。恢复28年时,粗根系半灌木的白莲蒿在群落中占比较大,对外源氮输入不敏感,经过长期施氮后,土壤养分状况改善,会促使主根数量减少,增加不定根数量,致使地下生物量减少,进而导致根冠比下降。恢复37年时,土质疏松,土壤中营养物质含量高,氮添加后对群落地上生物量的促进作用明显减弱,但氮添加后可能影响植物根系内部的分配[52],促使群落粗根和细根的比例在土层中发生变化[47],增加了地下生物量,致使根冠比升高。本研究通过对不同恢复期根冠比的探讨,可以为未来估测中国草原碳储量提供有力的数据支撑。
4 结论
本研究以黄土高原不同恢复年限天然草地为对象,旨在通过探讨氮添加处理对不同恢复期草地群落地上、地下生物量和根冠比的影响,为该区域草地的后续管理与利用提供理论指导与科学建议。研究结果表明,经过7a的氮添加后,恢复12年的草地,地上地下和总生物量均呈增加趋势,地上生物量对氮添加表现为非线性的正响应,但根冠比降低;恢复28年的草地,地上地下和总生物量表现为相反的趋势,地上生物量对氮添加表现为非线性的负响应,同时根冠比也下降;恢复37年时,地上地下生物量、总生物量和根冠比均呈上升趋势,地上生物量对氮添加表现为非线性的正响应。通过对比不同恢复年限草地生物量对氮添加的响应,揭示了氮添加后生物量的变化趋势,在氮沉降背景下,本研究建议对该区域开展试点实验,实行适应性草地管理,如进行两年一次刈割或轻度放牧(2只羊/hm2),通过科学的控制来管理和监测,实时调整有关措施,使草地实现系统性恢复,进而满足生态系统容量和社会需求的变化。本研究仅仅从氮添加对生物量的影响进行了初步探讨,尚未形成系统的理论支撑,未来仍需对氮添加后影响恢复草地的过程、机制及途径等理论问题进行系统性深入研究,进而实现以草地生态系统稳定平衡和多功能协同为目标的管理利用方式。本研究虽然进行了7a的连续观测,但观测时间尺度还远远不够,需要更长时间尺度的观测,同时应从生态系统整体出发,关注植被-土壤-微生物-环境协同响应氮沉降的内在机制。
猜你喜欢
现代畜牧科技(2021年4期)2021-07-21 06:13:08
今日农业(2020年19期)2020-12-14 14:16:52
中国科技纵横(2018年14期)2018-08-30 09:15:34
浙江农林大学学报(2018年4期)2018-08-22 01:54:12
现代农业科技(2017年16期)2017-09-22 14:47:46
中成药(2017年6期)2017-06-13 07:30:35
动物营养学报(2015年3期)2016-01-07 11:06:10
中国科技信息(2015年2期)2015-11-16 08:18:32
江苏农业科学(2014年1期)2014-07-18 21:21:34
植物营养与肥料学报(2014年1期)2014-03-11 20:24:38
