
阔叶红松林不同演替阶段灌木器官的N、P分配特征
2022-01-17 08:47:54吴一博金光泽
生态学报 2021年24期
吴一博,金光泽,2,*
1 东北林业大学生态研究中心, 哈尔滨 150040 2 东北林业大学森林生态系统可持续经营教育部重点实验室, 哈尔滨 150040
N、P是生态化学计量学的重要研究内容[1]。N、P作为植物体内主要的限制元素[2],是核酸、蛋白质、脂质和细胞结构的重要组成部分[3]。N、P的分配对植物的生长、代谢以及生活史对策起重要的调控作用[4-5]。不同的植物器官承担各自的生理功能,对N、P有不同的需求,因此植物对各个器官的营养分配权衡存在差异[6]。研究显示植物的N、P元素在不同器官中的分配具有显著差异,较高活跃度的叶和根等器官具有优先的分配倾向[7]。关于植物限制元素的判断依据,化学计量学中通常依靠N∶P作为指标[8]。当N∶P小于14时,N是影响植物的限制因子;当N∶P大于16时,植物主要受到P的限制;N∶P在两者之间时,植物则受到两种元素的共同限制[9]。目前养分限制的研究焦点多集中于叶片[10-11],对于其他器官如枝条、茎干缺乏详尽研究。另一方面,灌木群落在森林生态系统中占有重要地位,在维持生物多样性、养分循环和改善林下环境等方面发挥着关键作用[12-13],然而相关研究对其有所忽视[14]。因此,针对灌木不同器官的化学计量特征,以及营养限制研究需要更加深入的探索。
植物对限制元素的分配在器官水平上有所差异,同样在群落尺度上会受到周围环境的影响,尤其是在不同的演替阶段中[15]。演替不仅是植物群落动态变化的过程,同时也是植物对环境改变的适应过程[16]。Odum的经典演替理论认为生态系统在演替前期和演替后期的群落构建、物种适应对策以及土壤营养条件上有较大差异[17]。Huang等[18]通过对南亚热带森林的不同演替阶段研究发现P是生态系统主要的限制元素,对植物生长起到重要的调控作用。刘兴诏等[19]以南亚森林演替的3个阶段为研究对象,发现叶片的N、P随演替阶段的进展而减少,所有器官的N∶P随演替在不断增加。在温带森林演替的进展过程中,有关灌木限制元素的研究依然匮乏;灌木如何适应环境变化,不同器官之间的分配权衡是否受到影响,此类研究缺乏充足的数据支持,仍需进一步的实验分析。
阔叶红松(Pinuskoraiensis)林是我国东北东部山区的地带性顶极植被,物种丰富、生产力高,是温带森林的典型代表[20]。本研究以典型阔叶红松林4个演替阶段的6种主要灌木为研究对象,分析其不同器官的N、P的化学计量特征,旨在解决以下问题: (1)不同灌木器官之间的化学计量有何差异;(2)不同演替阶段之间灌木群落的N、P含量如何变化;(3)灌木不同器官如何分配与权衡N、P元素,是否存在保守性?研究重点关注不同演替阶段灌木的营养分配模式与适应对策,以期为该地区生态系统的养分循环与群落构建研究提供数据支持和科学依据。
1 材料和方法
1.1 研究区概况
本实验样地位于伊春市大箐山县的黑龙江凉水国家级自然保护区内(47°10′50″N,128°53′20″E),地处小兴安岭南坡达里带岭支脉东坡。该地区属于温带大陆季风气候,冬季寒冷干燥,夏季湿润炎热,年平均降水量676 mm,6—8月降雨量占全年的60%以上;年平均气温-0.3℃,年均最高气温7.5℃,年均最低气温-6.6℃,无霜期100—120 d,积雪期130—150 d。保护区海拔高度在280—707 m,土壤类型为地带性暗棕壤。阔叶红松林为本地区的主要优势群落,红松为建群种,紫椴(Tiliaamurensis)、色木槭(Acerpictumsubsp.mono)、枫桦(Betulacostata)等为主要伴生种[21]。
1.2 实验设计与样品采集
2019年7月,按照演替序列选择白桦(Betulaplatyphylla)次生林、阔叶混交林、针阔混交林和阔叶红松林等4个林型,每个林型设置3个20 m×20 m的样地,样地间隔大于20 m,与道路或者林缘间隔大于20 m,地形因子(海拔、坡位、坡向、坡度)相似。对每个样地进行调查后(群落特征详见表1),以刺五加(Acanthopanaxsenticosus)、暴马丁香(Syringareticulatavar.amurensis)、毛榛(Corylusmandshurica)、早花忍冬(Lonicerapraeflorens)、东北山梅花(Philadelphusschrenkii)、光萼溲疏(Deutziaglabrata)6种各演替阶段共有的优势灌木为研究对象,选取长势良好的个体作为标准木,每种灌木在每个样地选取3株标准木进行取样。
取样时将整株标准木挖出,按照叶、枝条、茎干和根等器官划分,每株标准木各器官收集1份足量的样品,共计取样864份。其中根选取直径<2 mm的细根,并除去附着的泥土;叶去除样品中受病虫害影响的部分;选取茎干的直径>2 cm,选取枝条的直径<1 cm。所有样品放在盛有冰块的保温箱中低温保存带回实验室。
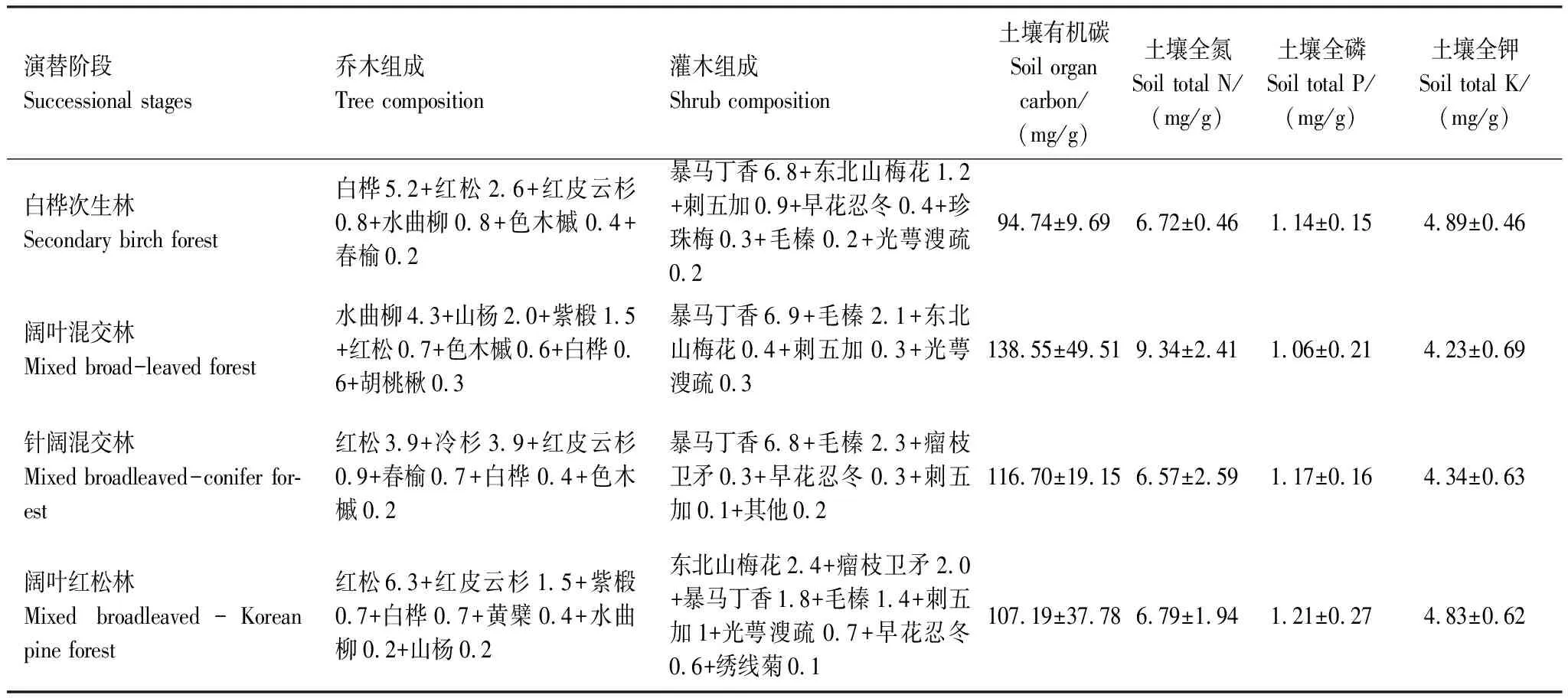
表1 阔叶红松林不同演替阶段群落特征
白桦(Betulaplatyphylla)、红松(Pinuskoraiensis)、红皮云杉(Piceakoraiensis)、水曲柳(Fraxinusmandschurica)、色木槭(Acerpictumsubsp.mono)、春榆(Ulmusdavidianavar.japonica)、山杨(Populusdavidiana)、紫椴(Tiliaamurensis)、胡桃楸(Juglansmandshurica)、黄檗(Phellodendronamurense)、暴马丁香((Syringareticulatavar.amurensis)、东北山梅花(Philadelphusschrenkii)、刺五加(Acanthopanaxsenticosus)、早花忍冬(Lonicerapraeflorens)、珍珠梅(Sorbariasorbifolia)、毛榛(Corylusmandshurica)、光萼溲疏(Deutziaglabrata))、瘤枝卫矛(Euonymusverrucosus)、绣线菊(Spiraeasalicifolia)。数据为均值(Mean)±标准差(SD)
1.3 样品测定与数据处理
所有样品带回实验室后使用烘箱在65℃下烘干至恒重,然后粉碎、过筛(100目)。将过筛的样品进行消煮后使用全自动间断化学分析仪(AQ400,Seal,Germany)测定全氮和全磷。
采用单因素方差分析法(one-way ANOVA)检验不同演替阶段灌木各器官之间的N含量、P含量以及N∶P,若方差齐性则采用最小显著性差异法(LSD)比较差异的显著性;若方差不齐则采用Tamhane′s检验法分析显著性,所有检验显著性设置为a=0.05,以上分析在SPSS 23.0完成。
采用异速生长方程分析不同器官的N和P之间的关系[22]。将不同器官的N含量和P含量进行对数转化后,采用如下公式进行计算分析:
lnN=mlnP+n
式中,m为异速生长指数,即异速生长方程的斜率,n为异速生长归一化常数,即方程的截距。对构建的方程采用标准化主轴分析法(SMA)分析N、P的异速生长关系,使用R语言中的smatr包来计算各参数[23],研究结果使用R语言中的ggplot2包进行绘图[24]。Smart包和ggplot2包均在R 4.0.1中运行。
2 结果
2.1 灌木不同器官的N、P含量和N∶P
如图1所示,灌木不同器官的N、P含量以及N∶P差异均显著(P<0.05),并且N与P分配具有一致性。叶的N、P含量最高,分别为19.06 mg/g和2.64 mg/g;茎干的N、P含量最低,分别为5.49 mg/g和1.04 mg/g;根的N、P含量均显著高于枝条,根和枝条的N含量分别为10.74 mg/g和8.70 mg/g,P含量分别为2.30 mg/g和1.65 mg/g。灌木各器官的N∶P均小于10,其中叶的N∶P显著高于其他器官 (P<0.05);根的N∶P显著高于茎干(P<0.05),而茎干、根的N∶P与枝条间相比无显著差异(P>0.05)。
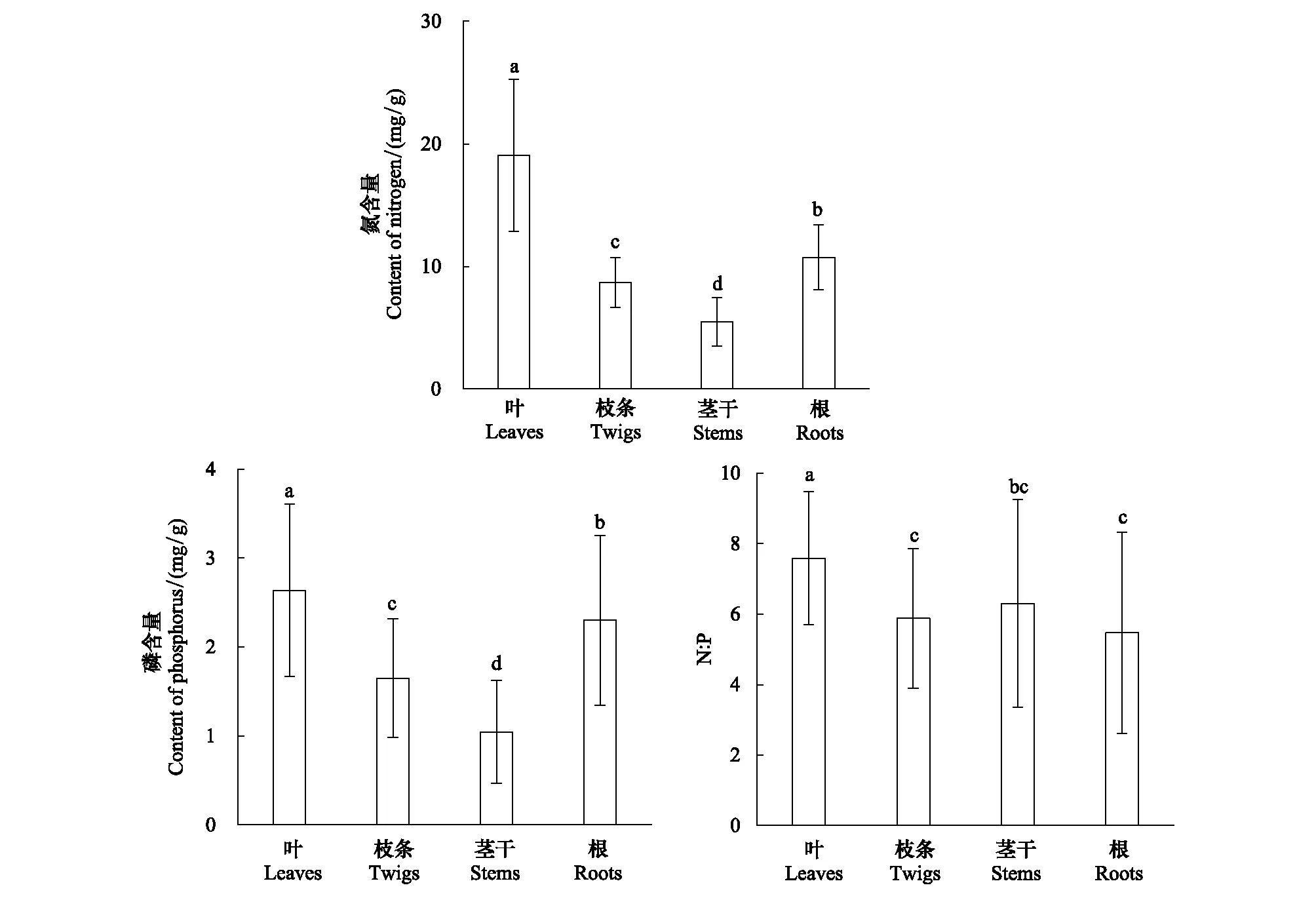
图1 阔叶红松林主要灌木器官的化学计量特征Fig.1 The stoichiometric traits of major shrub organs in the mixed broadleaved-Korean pine forest不同小写字母代表不同器官间存在显著差异(P<0.05)
2.2 不同演替阶段灌木器官的N、P化学计量特征
由表2发现,不同演替阶段中灌木叶的N含量差异显著(P<0.05),随着演替有增加的趋势;茎干中的N是先减小后增大的趋势,其他器官中无明显趋势;茎干、根和枝条的N含量在不同演替阶段没有显著差异(P>0.05)。不同演替阶段中灌木器官的P含量的变化较为复杂,叶的P含量有上升的趋势;枝条和茎干的P含量呈现先增大后减小的趋势;根的P含量变化无明显规律,整体上各演替阶段灌木器官的P含量差异均不显著(P>0.05)。不同演替阶段中,只有枝条的N∶ P具有显著差异(P<0.05),而其他器官均差异不显著(P>0.05)。
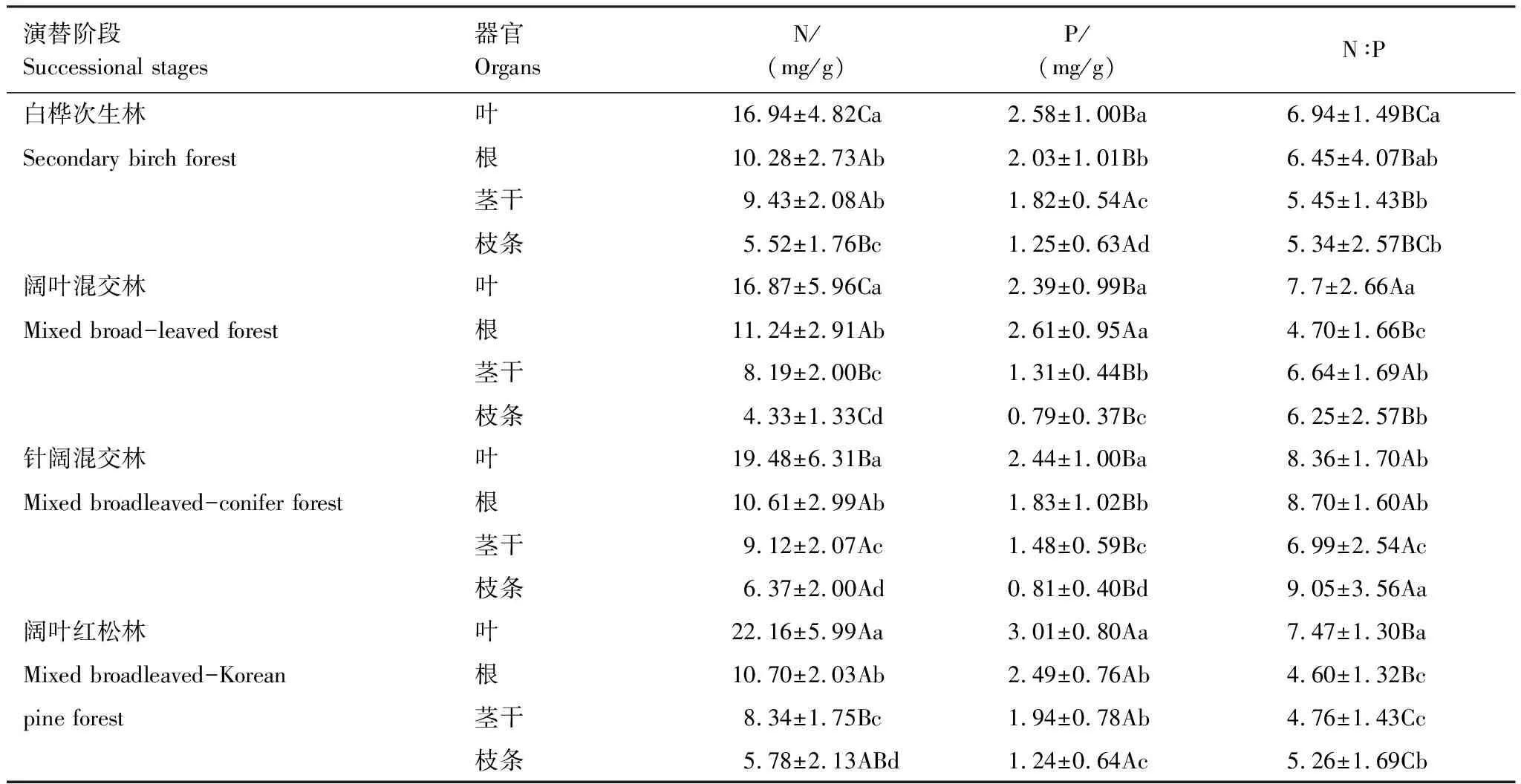
表2 阔叶红松林不同演替阶段灌木器官的化学计量特征
2.3 不同演替阶段灌木器官的异速生长关系
如图2所示,除白桦林的灌木茎干外,灌木器官的N、P含量经过对数转化后呈现显著的线性正相关关系(P<0.05)。不同演替中灌木器官异速生长指数均小于1,说明N、P元素的异速生长关系普遍存在于灌木器官中,并且P含量的增加速度高于N。所有器官中,叶的异速生长指数最高,随着演替有增加趋势,范围为0.71—0.80;根的异速生长指数最小,范围为0.39—0.69;而枝条和茎干的异速生长指数范围分别为0.5—0.72和0.66—0.76(表3)。SMA结果显示,整体上不同演替阶段各器官之间的异速生长指数无显著差异(P>0.05)。

图2 阔叶红松林不同演替阶段灌木器官N、P的异速生长关系Fig.2 Allometric relationships between N and P of shurb organs in different successional stages of the mixed broadleaved-Korean pine forest虚线表示N、P的异速生长关系不显著(P>0.05);白桦次生林中叶和枝条斜率差异不显著(P>0.05);阔叶混交林中四种器官斜率均不显著(P>0.05);针阔混交林中根和枝条斜率差异不显著(P>0.05);阔叶红松林中枝条、根和茎干斜率差异不显著(P>0.05)
3 讨论
3.1 N、P元素在灌木不同器官间的分配差异
本研究中灌木器官的N含量和N∶P低于19.1 mg/g和14.7的全国灌木平均水平,而P含量高于全国的平均水平(1.11 mg/g)[25]。灌木不同器官的N、P差异显著,说明灌木在生长过程中对各器官的营养分配有所权衡。在灌木所有器官中,叶的N、P含量均为最高,这与Zhang等[26]的研究结果一致。王晓洁等通过对比该地区常见植物的叶片与细根亦得到相似的结果,即叶片的N、P和N∶P显著高于细根[27]。叶作为光合作用的主要器官,承担养分制造与储藏的重要功能,被认为是植物器官中最活跃的器官[7]。N含量直接影响光合作用中相关蛋白质的合成,进而影响光合效率[28]。P是影响植物代谢过程的重要元素,直接参与rRNA的合成[29-30],较高的P含量是植物营养供给的重要保障[31]。因此在养分的分配权衡上,叶的优先分配是植物保证自身生存的重要策略。根作为养分吸收和运输的重要器官,在营养分配中同样占有重要地位[32-33]。茎干和枝条在植物体中具有骨架支撑作用,连接地上和地下部分进行营养物质的运输,同样是不可忽略的重要器官[34]。本研究中根的N、P含量仅次于叶片,高于枝条和茎干,这同样说明灌木器官的营养分配与器官的活跃性有关。Zhang等[14]的结果也证实了这一点,即越活跃的器官具有较高的营养分配比重。对活跃器官的优先分配权衡是灌木合理利用资源、达到自身优化配置的重要生存策略。
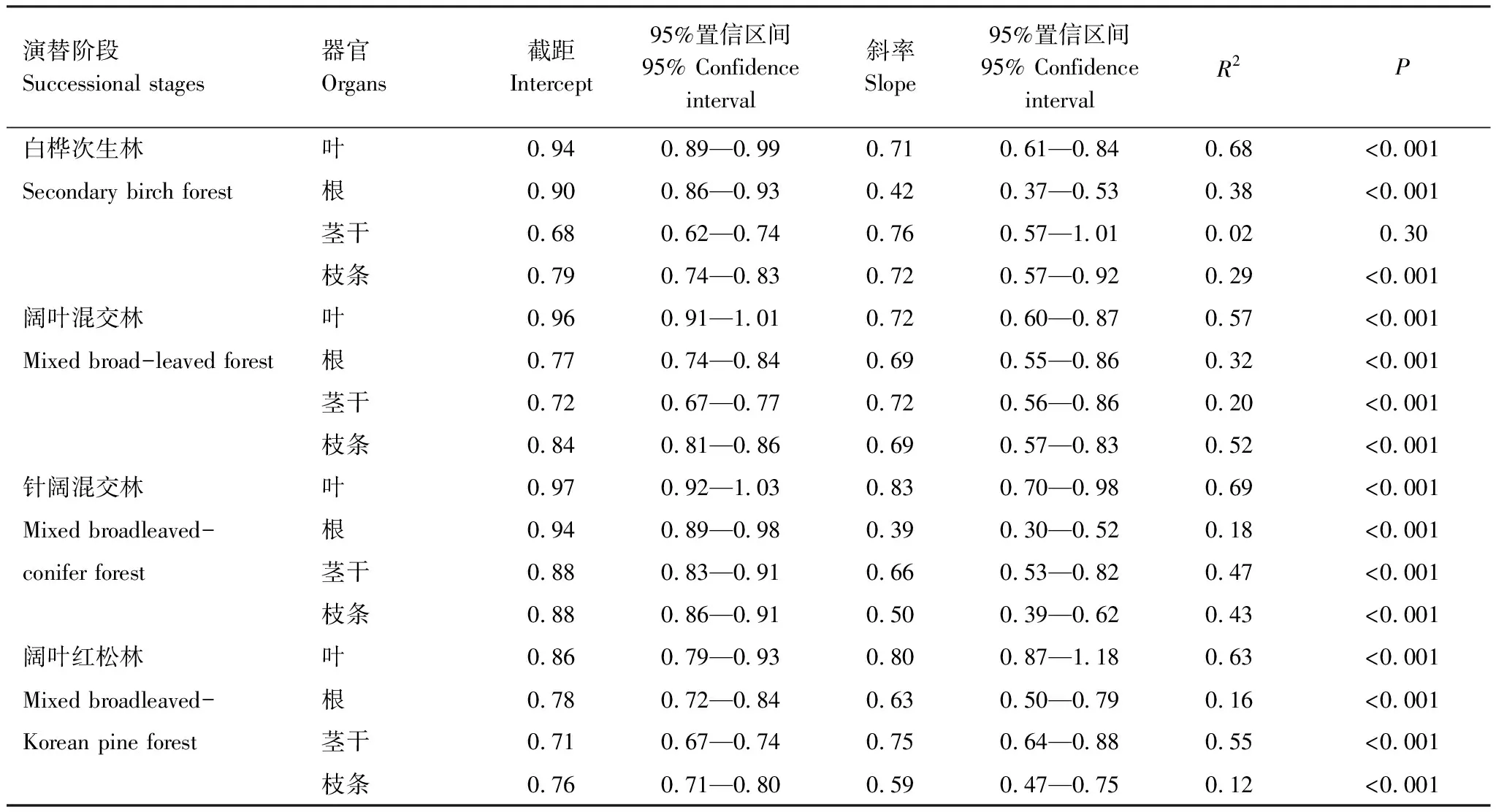
表3 阔叶红松林不同演替阶段灌木器官N、P异速生长方程的主要参数
灌木叶的N含量随着演替阶段不断增加,并且差异显著;其他器官N含量差异在各演替阶段不显著。结果同时显示,不同演替阶段灌木器官的P含量没有显著的差异。相关研究表明,随着演替的进行,环境会发生较大幅度的变化,如土壤养分改变,林下光资源减少,物种组成改变等[35-36],植物需要通过改变性状来适应环境变化。Graven等[37]研究发现叶的N含量会随着演替的进行增加,与本研究得到的结果一致。Liang[38]研究长白山阔叶红松林不同演替阶段主要树种的叶片功能性状发现,叶片结构和叶的N含量对光合作用效率起主要的调控,并且演替后期的植物群落具有更高的N利用效率。较高的N含量可以保证处于林冠下层的灌木具有较高的光合效率,以此应对演替产生的环境变化,维持自身的正常生长。本研究发现演替对灌木各器官的P含量没有影响,这可能与P的性质有关。P元素作为岩石风化沉积的产物,在森林中迁移较慢,不同森林土壤的P含量通常不具特异性[39],造成灌木各器官的P含量差异不显著。
3.2 N是各演替阶段中灌木器官的限制元素
在本研究的结果中,灌木器官的N∶P均小于14,说明阔叶红松林内的灌木普遍受到N元素的限制,这符合温带森林中N元素限制格局[40]。高三平等[41]研究天童山不同演替阶段叶片的化学计量特征时得出了相同的结果,即不同演替阶段均受到N元素的限制。王凯等[42]通过对榆树(Ulmuspumila)幼苗进行氮添加实验发现,氮的输入显著增加了各器官的N含量以及N∶P,进一步证实N对植物的生长发育过程具有重要的限制作用。在不同的演替阶段,土壤养分中N含量差异较大,尤其是演替后期因为生物固氮受限,淋溶、干扰等非受控的氮损失影响,群落更容易受到N元素的限制[43]。本研究中,灌木叶片的N∶P最高,这是由于叶片具有较高的N含量。在受到N元素的限制作用下,灌木倾向于优先分配N元素于叶片中,对于营养器官较多的投入是灌木确保自身生存的一种基本策略[7]。N∶P的结果同时说明P不是主要的限制元素,灌木在P元素的分配上采取保守的分配策略。化学计量学的内稳性假说认为,生物体可以维持自身化学组成的相对稳定[29]。当P没有成为限制元素时,不同演替阶段灌木P含量在各器官间可以保持相对稳定,因此各个器官的P含量在不同演替阶段没有差异。
3.3 N、P的异速生长关系表明灌木器官的保守分配策略
分析结果发现,N、P元素的异速关系在灌木中具有普遍性。不同演替阶段间灌木枝条、根和茎干的异速生长指数整体上没有显著变化,保守性较强。灌木叶片的异速生长指数在0.71—0.80之间,高于灌木其他器官,并且高于全球水平(0.62—0.73)[22]。在演替后期的两个林型中,叶的异速生长指数与其他器官差异显著,保守性较差。这主要是由于叶中N含量升高造成的,可能与环境变化有关[44]。在演替的进展过程中,上层林冠郁闭,减少了下层的光照,对灌木叶的经济性状造成了影响,直接导致叶对N的需求不断增加。Jiang等[45]研究该地区的群落发现叶的经济性状与土壤养分联系紧密。张增可等[46]分析海南岛植物群落发现叶功能性状在演替前期与后期有较大差异,植物的比叶面积和叶P含量在演替前期较高,后期叶干物质含量和叶N含量显著增加。演替前期的灌木为了维持较高的生长率需要维持高的P含量与低的N∶P,对P的需求较高,因此P的增加速率大于N[47];演替后期由于光资源的获取减少、竞争加剧,叶对N元素的需求增加以此提高光合效率。除阔叶混交林外,并非所有灌木器官在分配上均具有保守性,这可能是受地区尺度限制的原因。因此,今后的相关研究中应进一步拓宽尺度,以期寻求更加普适性的规律。
4 结论
阔叶红松林各演替阶段的灌木在养分分配上采取的是保守的策略,即(1)活跃度高的器官具有较高营养分配比重;(2)各器官中N和P的异速生长关系普遍存在。同时,发现N是各演替阶段中限制灌木生长的重要因子,保守的分配策略为灌木适应环境变化,优化资源配置,满足生长发育需求提供了基础的保障。
猜你喜欢
南京林业大学学报(自然科学版)(2022年1期)2022-11-29 10:10:16
小哥白尼(野生动物)(2021年9期)2022-01-17 02:31:00
中国林副特产(2020年5期)2020-10-09 08:24:14
农业机械学报(2020年3期)2020-04-01 08:11:10
种子(2019年5期)2019-07-02 11:37:44
乡村科技(2018年8期)2018-02-13 12:16:18
防护林科技(2017年7期)2017-09-01 01:08:43
诗潮(2017年2期)2017-03-16 10:42:30
湖南林业科技(2017年1期)2017-02-06 05:29:17
天然产物研究与开发(2016年11期)2016-06-15 20:29:15
