
农田土壤中真菌群落在时间序列对铬胁迫的响应机制*
2022-01-15 03:09赵鑫宇景秀清赵晓东燕平梅赵鹏宇
中国生态农业学报(中英文) 2022年1期
白 雪,赵鑫宇,景秀清,赵晓东,燕平梅,赵鹏宇**
(1.太原师范学院 榆次 030619; 2.山西农业大学 太谷 030800)
随着工业的发展,如制革、冶金等,铬 (Cr) 已成为地下水与土壤中最常见的重金属污染之一。研究表明我国农田土壤环境的重金属污染总超标率为16.1%,其中山西省土壤重金属Cr污染的平均含量高于全国均值。由此,Cr已经成为我国土壤主要重金属污染物之一,因其具有高移动性和毒害性等特征,可通过质膜进入机体导致遗传物质改变,因此对农田土壤、植物生长、粮食安全生产乃至人体健康都产生了严重威胁。重金属Cr在自然界中存在不同的氧化价态,其中三价 (Ⅲ) Cr是人体所必须的微量元素,可调节血糖; 而六价Cr (Ⅵ)却具有严重的毒害作用,较高的化学活性使其更易被植物吸收。有研究表明0.5 mg·L的Cr (Ⅵ)溶液就会对植物产生毒性作用。根际土壤是生物代谢的活跃位点,Cr在植物体内的迁移与积累也大多发生在根际土壤。而土壤中重金属积累必然会导致土壤中原始微生物群落结构发生变化,显著影响土壤生态系统的结构与功能。
微生物虽占土壤有机质较少一部分,但土壤微生物是决定土壤生物化学特性的重要组分,参与土壤的养分循环、污染物降解等过程,对重金属敏感性极强。因此,其结构与功能变化是评价土壤环境的有力指标。鉴于农田土壤环境污染的严峻性以及重金属污染的长期危害性,近年来农田土壤重金属污染、土壤微生物群落适应并响应重金属污染的机制已成为国内外学者研究的热点。Wang等发现微生物对重金属敏感性依次为Cr>Pb>As>Co>Zn>Cd>Cu,不同重金属对微生物群落结构也有不同影响。目前研究多集中于细菌对重金属污染的响应上,特别是对细菌群落多样性的影响。如Orwin等、于皓等研究表明,高浓度的Cu、Cr污染显著影响土壤细菌群落的结构组成与α多样性; Chai等研究表明从污染土壤中分离出的细菌菌株具有明显的Cr抗性和Cr还原能力; 但Wang等研究表明湿地植物风车草 () 在不同栽培模式下,Cr胁迫并未引起根际细菌群落结构的显著差异。
由于真菌具有孢子、菌核等耐性较强的组织结构,因此真菌广泛存在于各种类型的土壤,具有较高的系统发育和功能多样性,且对重金属污染耐性较强。真菌作为土壤生态系统的重要组分,对不同重金属的反应也有所不同。如Hu等研究表明土壤真菌群落通过复杂的相互作用以适应长期Cr污染;刘晋仙等研究发现重金属复合污染的尾矿废水中环境因子对真菌群落的α多样性有显著影响,而群落的β多样性主要受种间相互作用关系的影响。目前关于Cr胁迫土壤微生物群落的研究相对较少,且研究多集中于土壤细菌,对于Cr胁迫时间序列上农田土壤中真菌群落响应机制的研究还有所缺乏。
此外,本研究将Cr胁迫视为一种“生态扰动”,它会引起瞬时的土壤微生物群落破坏,导致微生物群落重建和功能恢复过程的差异,进而探究农田土壤生态系统中真菌群落的动态变化过程。‘晋谷21号’是山西省农业科学院经济作物研究所最新培育的优质高产谷子 () 新品种,出籽率高、结实性好、幼苗生长势强、早熟,且抗病稳产。本研究以种植过‘晋谷21号’谷子的土壤为研究对象,通过添加外源Cr探究Cr胁迫时间序列上农田土壤中真菌群落的响应机制、群落构建驱动机制及功能预测分析,揭示土壤中真菌群落在Cr胁迫时间序列上的动态变化规律,为‘晋谷21号’谷子安全栽培及Cr污染环境治理以及恢复提供参考。
1 材料和方法
1.1 材料
供试材料为山西省农业科学院经济作物研究所最新培育的优质高产谷子新品种‘晋谷21号’。研究所用土壤为褐土,土壤pH平均为7.65,总氮为0.13 g·kg,总磷为1.32 g·kg,不含Cr,土壤有机质(SOC)为32 g·kg。
药剂为重铬酸钾(KCrO,1 mmol·LCr)。
1.2 试验及取样的设置
本研究在实验室中进行盆栽试验。挑选颗粒饱满、大小均匀的‘晋谷21号’种子若干,用自来水洗去包衣后,置于垫有湿纱布的盒中为种植做预处理。准备15个花盆,每个花盆直径15 cm、盆深20 cm,种植前先用灭菌蒸馏水调整土壤湿度至田间持水量的40%,以满足谷子苗期的生长需求; 每盆500 g土壤,均匀点放40~50颗‘晋谷21号’种子; 随后用保鲜膜包裹花盆,放在智能光照培养室中培养,直至种子发芽(包裹保鲜膜是为防止空气中的杂菌污染土壤)。控制培养温度在26 ℃促进种子的萌发与生长。培养15 d左右待谷子长势均一时,在试验组托盘中每盆施加300 mL的1 mmol·LCr。培养期间种子出苗率为97%。
试验分3个处理 (对照和两个Cr胁迫处理): 对照 (CK)、Cr胁迫6 h (Cr_6 h)、Cr胁迫6 d (Cr_6 d)。每个处理6个重复,共18个样本。本研究参考肖文丹等研究,设置两个Cr胁迫处理 (Cr_6 h和Cr_6 d),原因在于当Cr胁迫约6 h,土壤中真菌群落对Cr所造成的生态扰动会产生一定的应激反应; 而当Cr胁迫约6 d时,群落逐渐修复并呈现出稳定的状态。但由于重金属污染周期长、难降解,因此未对6 d后土壤样本做后续分析。
取样尽量保持根际土壤的完整性,不破坏取样。所取土样储存在—80 ℃冰箱待用,用于后续序列测定。
1.3 测定方法
1.3.1 DNA的提取
称取1 g土壤,用Power Soil试剂盒提取土样DNA。利用琼脂糖凝胶电泳检测DNA提取物是否降解,提取物纯度与浓度用酶标仪测定,当A260/A280吸光度在1.8~2.0时,表明所提取的DNA纯度较高,可用于下一步分析。
1.3.2 PCR扩增与测序
将提取的基因组DNA进行PCR扩增,PCR扩增体系如下 (25 μL): Taq DNA聚合酶 (5 U·μL) 0.5 μL,10×PCR 缓冲液5.0 μL,dNTPs溶液4.0 μL,模板DNA 5.0 μL,引物 (50 μmol·L) 0.5 μL,用无菌 水补至25 μL。PCR反应条件: 预变性98 ℃ 1 min; 98 ℃ 10 s,50 ℃ 30 s,72 ℃ 30 s,共30个循环; 终延伸72 ℃ 5 min。混匀的DNA样品送至上海美吉生物技术工程有限公司上机测序,以ITS1F_ITS2R为序列扩增区域,通过Illumina Miseq测序平台对ITS进行测序,测序完成后进行数据分析处理。
1.4 生物信息学与统计学分析
测序平台所得的原始数据通过序列拼接、质控等得到高质量序列。按照97%的序列相似性将序列聚类为OTU,以OTU丰度数据为基础进行物种注释分析,统计土壤样本中真菌群落的组成状况。
用软件Excel 2007辅助数据分析与作图。利用单因素方差分析 (ANOVA) 评估不同处理土壤真菌群落的组成结构、α多样性、群落构建过程及功能预测分析的差异。利用Sobs指数、Shannon指数等探究真菌群落的α多样性。利用非度量多维尺度分析 (NMDS) 对Cr胁迫后不同时间真菌群落的时空分布格局进行排序分析,以Stress值验证分析结果的好坏,通常认为stress<0.2时图形具有一定解释意义。利用FUNGuild功能预测,划分对环境资源吸收与利用相似的类群。根据营养方式,真菌中Guild种类划分为3类: 病原营养型 (pathotroph)、共生营养型 (symbiotroph) 和腐生营养型 (saprotroph),进而对扩增子测序结果中真菌群落的类别及具体功能进行预测分析。
1.5 群落构建分析
群落构建分析过程中,种间网络分析的参数运算用R语言igraph程序包,并在Gepih软件中实现可视化 (https://gephi.org/)。用R语言的Hmisc等程序包构建中性群落模型 (NCM),进一步判断土壤真菌群落的构建机制。参考Stegen等提出的基于系统发育结构的分析方法,判断土壤中真菌群落构建过程中的驱动因素。当所有样品中NTI的中位数在—2~2之间,说明随机过程主导微生物群落结构的变化,当NTI的中位数大于2或小于—2,说明确定过程主导微生物群落结构的变化。
2 结果与分析
2.1 Cr胁迫时间序列上的真菌群落结构
2.1.1 真菌群落组成
通过高通量测序,共获得1 286 712个有效序列,比对出1304个真菌分类单元(OTUs)。观测Rank-Abundance曲线,曲线走势趋于平缓,表明测序量基本能达到分析要求。观察韦恩图(图1),3种处理(CK、Cr_6 h、Cr_6 d) 共检测出578个共有OTUs(占所有OTUs的44.33%)。CK、Cr_6 h和Cr_6 d阶段特有OTUs分别为170个 (13.04%)、155个 (11.89%)和118个 (9.05%)。
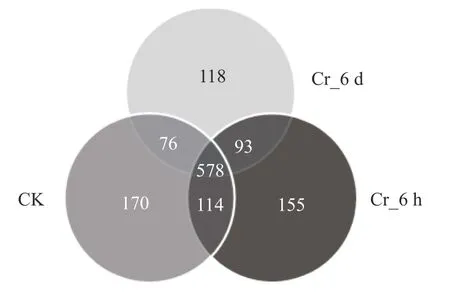
图1 Cr胁迫时间序列上的真菌群落OTUs Venn图Fig.1 OTUs Venn diagram of fungal communities in the Cr stress time series
2.1.2 真菌群落的结构分析
门水平相对丰度大于10%的真菌门共比对出5种 (图2A),其中子囊菌门(Ascomycota) 平均相对丰度最高(87.64%),其次是担子菌门(Basidiomycota,3.02%)、壶菌门(Chytridiomycota,1.56%)、被孢菌门(Mortierellomycota,1.13%) 和罗兹菌门 (Rozellomycota,0.27%)。子囊菌门相对丰度在Cr胁迫时间序列上先升后降,担子菌门和壶菌门先降后升,且壶菌门在Cr胁迫时间序列上丰度变化显著 (<0.01); 被孢菌门(Mortierellomycota) 在Cr胁迫时间序列上丰度下降极显著 (<0.001),其他菌群则无显著差异(>0.05)。
属水平相对丰度大于20%的真菌属共比对出15个(图2B),其中丰度前5的真菌属包括毛壳菌属(,平均相对丰度8.54%)、赤霉菌属 (,7.31%)、镰刀菌属 (,6.63%)、金孢子菌属(,5.61%) 和瓶毛壳属(,4.66%)。在Cr胁迫时间序列上,镰刀菌属相对丰度先下降后显著增加 (<0.05),赤霉菌属与噬热直菌属() 分别呈极显著(<0.01) 和显著下降(<0.05),毛壳菌属、金孢子菌属、瓶毛壳属与地丝霉属() 先升后降,其中瓶毛壳属变化下降(<0.05)。另外,粉褶蕈属() 在Cr_6 d阶段单独存在,相对丰度为3.19%。上述结果表明Cr胁迫导致真菌群落组成结构变化显著。
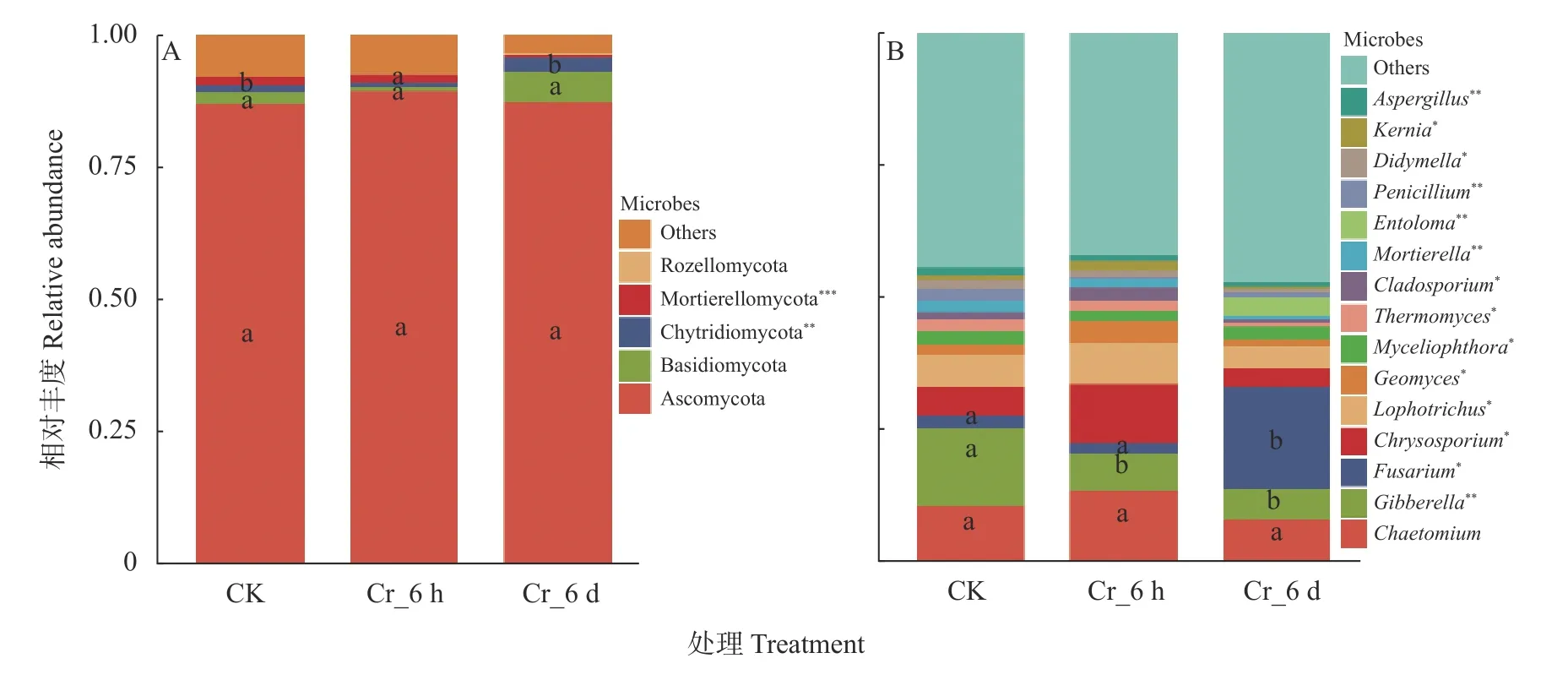
图2 Cr胁迫时间序列上门水平(A)和属水平(B)真菌群落结构与差异性Fig.2 Phylum level (A) and genus level (B) fungal community structure and differences among treatments in the Cr stress time series
2.1.3 真菌群落的多样性
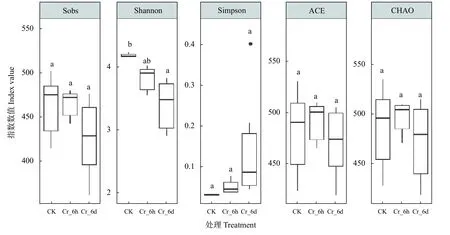
图3 Cr胁迫时间序列上真菌群落多样性指数分析Fig.3 Analysis of fungal community diversity indexes in the Cr stress time series
通过基于Bray_Curtis距离的非度量多维尺度分析 (NMDS),探明土壤中真菌群落的时空分布格局(图4)。土壤中真菌群落结构在Cr胁迫相同时间内更相似,而在不同胁迫时间间差异较大。通过组间差异显著性分析 (ADONIS,=0.001),土壤中真菌群落的时空分布格局在Cr胁迫时间序列上差异显著。
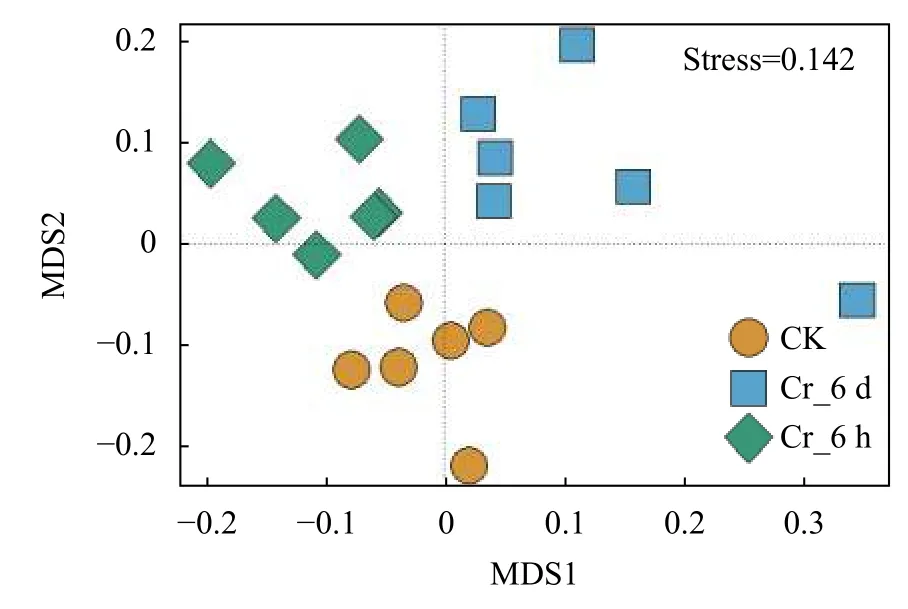
图4 Cr胁迫时间序列上真菌群落的非度量多维尺度分析Fig.4 Non-metric multidimensional scale analysis of fungal communities on the Cr stress time series
2.2 Cr胁迫时间序列上真菌群落构建机制
根据Cr胁迫不同时间段土壤真菌群落的NTI值 (图5),结合Stegen等的群落构建的划分方法,推断Cr胁迫时间序列上群落构建过程中随机过程占主导地位 (NTI|<2),NTI值 (即箱线图中的中位数) 在CK、Cr_6 h、Cr_6 d阶段分别为—0.16、—0.71、—0.23。
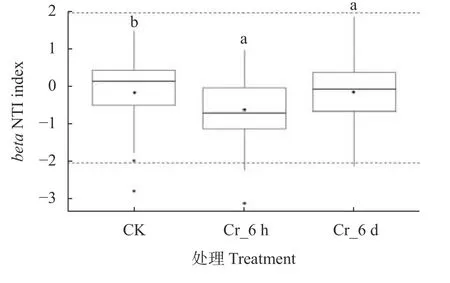
图5 Cr胁迫时间序列上土壤真菌群落的beta NTI值Fig.5 beta NTI values of soil fungal communities in the Cr stress time series
2.2.1 真菌群落种间共生网络
通过真菌群落共发生网络分析,利用Spearsman相关性分析评估不同菌群间的相关性,反映特定生境下真菌菌群的共存格局; 通过构建种间共生网络图,探究Cr胁迫时间序列上土壤真菌群落的种间关系。据表1,Cr_6 d的共生网络的边数量、平均度和平均路径长度均较CK和Cr_6 h更高,表明Cr_6 d的真菌网络规模更大,种间相互作用的关系更复杂。
据种间共生网络图 (图6) 分析可知,Cr胁迫时间序列上的关键物种不同: CK阶段的关键物种为金孢子菌属和毛壳菌属等,Cr_6 h阶段为赤霉菌属和镰刀菌属等,Cr_6 d阶段为赤霉菌属、青霉菌属() 和瓶毛壳属等。不同阶段真菌群落多病原、病理型真菌,且真菌群落模块变化趋势与表1一致;同一模块中以正相关为主,表明Cr胁迫处理后真菌群落以共生关系为主,而竞争关系较弱。
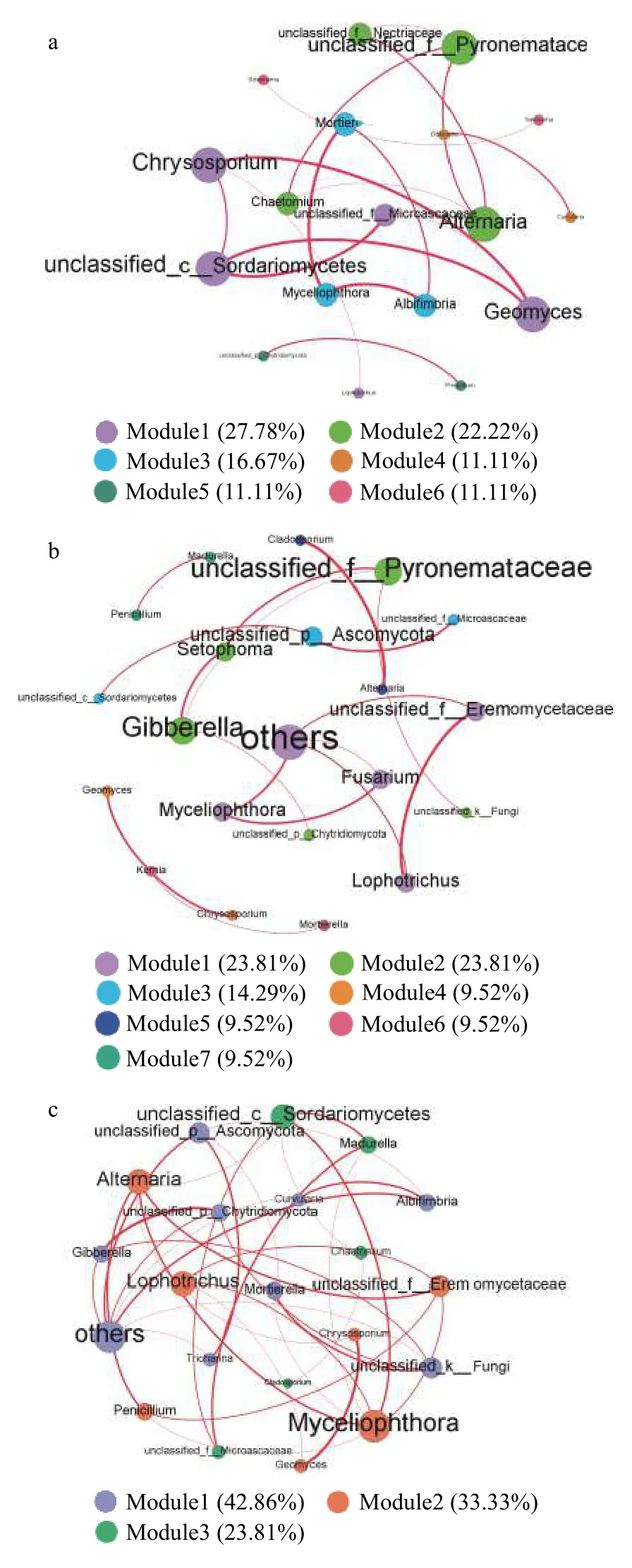
图6 Cr胁迫时间序列上的真菌种群间共生网络图(a:CK; b: Cr_6 h,Cr胁迫后6 h; c: Cr_6 d,Cr胁迫后6 d)Fig.6 Inter-population symbiosis network diagram of fungi in the Cr stress time series (a: CK; b: Cr_6 h,Cr stress for 6 hours; c: Cr_6 d,Cr stress for 6 days)
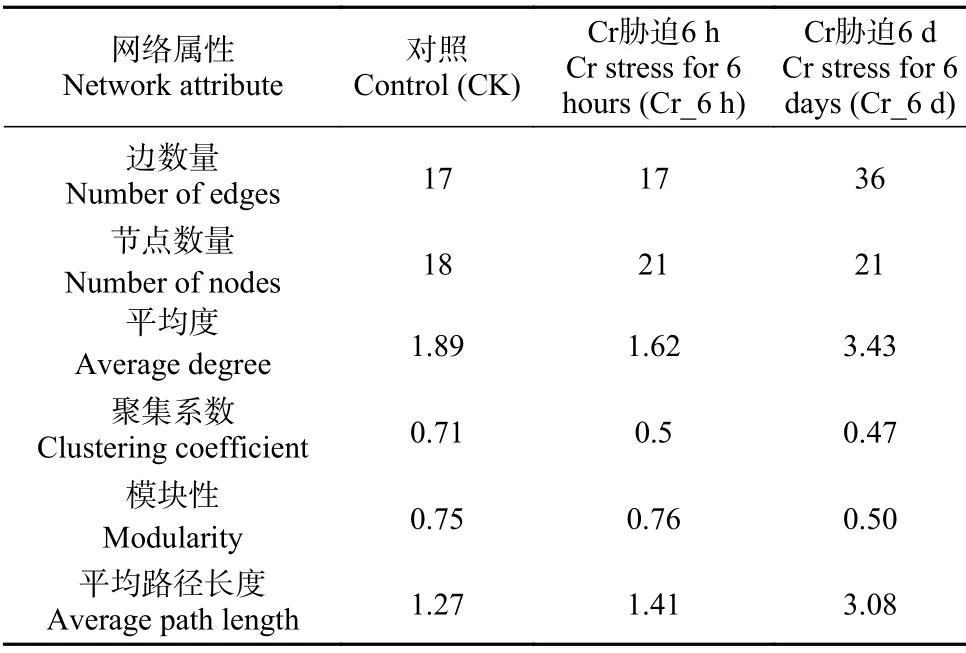
表1 Cr胁迫时间序列上的真菌群落网络属性表Table 1 Network attributes of fungal communities on the Cr stress time series
2.2.2 群落构建的随机过程
图5显示Cr胁迫时间序列上真菌群落|NTI|<2,表明真菌群落构建由随机过程主导。因此本研究通过构建Sloan等提出的中性群落模型进一步量化随机过程。
在变电站汇控箱安装模块化的智能温湿度测控器,实施监测、显示、记录每台抽湿机的工作状态,并可通过终端监控软件平台手动对抽湿机的启停进行远程控制,将湿度参数与抽湿机启停进行联动,当湿度超高时,自动启动抽湿机,消除湿气对设备的腐蚀危害,提高了电气绝缘性能,保障设备的运行安全[1]。
基于OTU矩阵绘制中性群落模型 (NCM,图7),估算OTU发生频率与其相对丰度变化间的关系。CK、r_6 h和Cr_6 d处理的模型分别解释了群落方差的59.3%、61.4%和63.8%,总体解释度为73.9%。NCM在Cr胁迫时间序列上较高的解释率 () 说明该模型可较好地拟合土壤中真菌群落的分布模式,且随机过程在土壤真菌群落的构建中起重要作用。在Cr胁迫时间序列上,真菌群落值[元群落规模 () 与迁移率 () 的乘积]在CK (=70 723.00;=0.066)、Cr_6 h (=71 839.83;=0.132)、Cr_6 d(=70 951.83;=0.163) 阶段逐渐变大,表明真菌群落受到的扩散限制作用变小,分布变广泛。
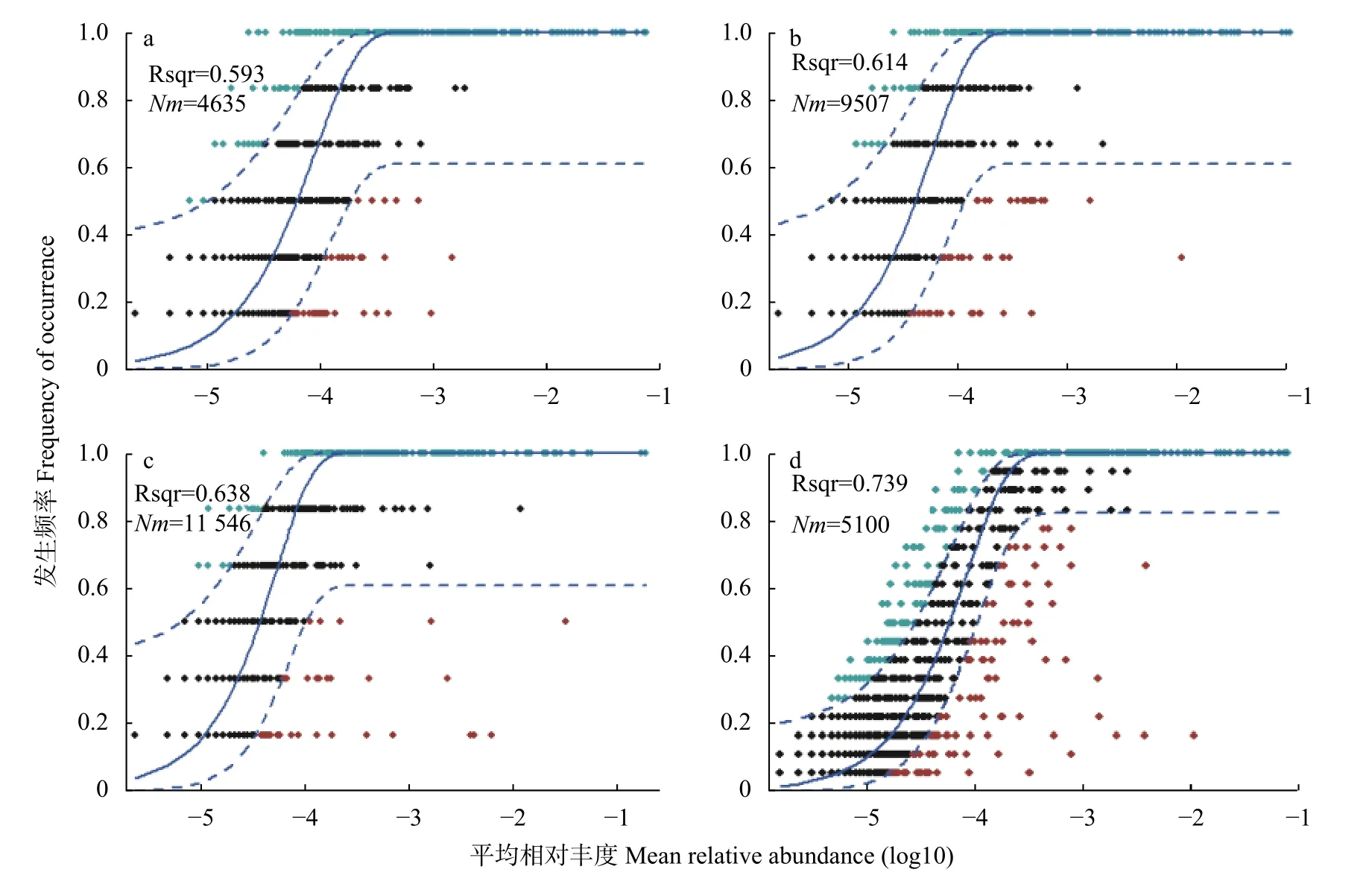
图7 Cr胁迫时间序列上的中性真菌群落模型(a: CK; b: Cr_6 h,Cr胁迫后6 h; c: Cr_6 d,Cr胁迫后6 d; d: 整体水平)Fig.7 Neutral community model in the Cr stress time series (a: CK; b: Cr_6 h,Cr stress for 6 hours; c: Cr_6 d,chromium stress f or 6 days; d: overall level)
2.3 Cr胁迫时间序列上真菌群落功能
利用FUNGuild功能预测,分类鉴定真菌群落的营养类型 (表2),并进行单因素方差分析筛选出真菌群落在Cr胁迫时间序列上的营养类型(丰度前14)。大致分3种: 腐生营养型、病原-腐生营养型、病原营养型。结果表明,未定义的营养类型 (unknown) 丰度居首位 (20.77%); 腐生营养型 (0.88%~18.78%) 与病原-腐生营养型(1.10%~14.37%) 居多,同时含有病原营养型 (1.24%~9.67%) 也较多。部分动物病原营养型、植物病原营养型以及未定义的腐生营养型在Cr_6 h、Cr_6 d中显著下降(<0.05),而部分植物病原营养型在CK到Cr_6 h呈显著上升趋势(<0.05)。
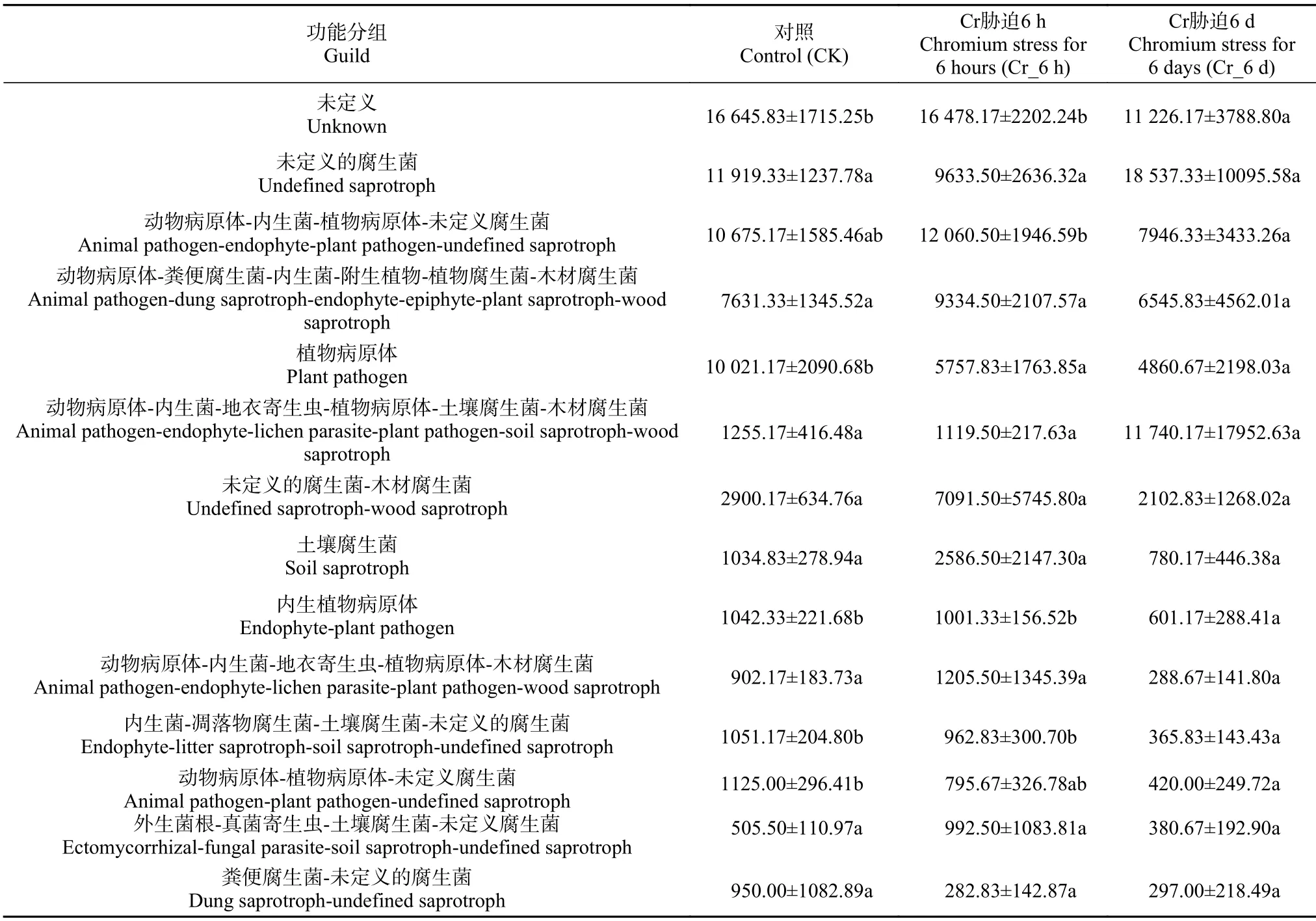
表2 Cr处理时间序列上真菌功能丰度Table 2 Abundance of fungal functional diversity in the Cr stress time series
3 讨论
3.1 土壤真菌群落组成结构对Cr胁迫时间序列的响应
本研究发现Cr胁迫时间序列上,土壤中真菌群落的组成结构差异显著,且不同群落生长策略与营养摄取能力有所不同。1)门水平中,子囊菌门丰度最高,其次是担子菌门,这是健康土壤特性的指标。原因可能在于子囊菌门能产生降解酶以高效利用底物,从而最大程度地将资源转化为自身生物量。而担子菌门丰度先降后升,分别在Cr_6 h、Cr_6 d阶段呈现出由应激到稳定的动态变化过程。担子菌门中许多菌种与重金属有良好的结合能力,如Basidiomycete BDT-14具有很高的除Cr潜力,对极端环境适应性较强。2) 属水平中,子囊菌门的曲霉属 () 丰度随Cr胁迫时间增长而显著下降,而青霉属 () 丰度呈现显著性的由应激到稳定的动态变化过程。曲霉属与青霉属广泛存在于土壤中,有研究表明二者具有抗Cr性,可将Cr (Ⅵ)降低到Cr (Ⅲ),且对Cr具有极大的生物吸附潜力。但丰度下降表征Cr胁迫对真菌群落产生了较大的毒害作用。被孢霉属()、赤霉菌属等对Cr较为敏感,在Cr_6 h阶段丰度下降极显著。镰刀菌属呈现出与担子菌门一致的,由应激到稳定的动态变化过程,可能是由于其病理型营养方式通过破坏植物根系细胞,获取生长所需营养物质,且对较低浓度的Cr有一定抗性。但同时有研究表明某种耐受重金属的内生镰刀菌可促进植物积累重金属,因此其在Cr_6 d阶段丰度升高可能对重金属Cr污染的土壤起到一定修复作用。毛壳菌属作为生物防治菌,可抑制黄瓜菌核病等蔬菜类疾病,其丰度变化可能由于Cr胁迫及土壤生态系统的复杂性不利于其生长。
由此可见,Cr胁迫可能改变了土壤真菌群落中耐性菌与敏感菌的比例,进而影响土壤中真菌群落的组成结构; Cr胁迫处理6 h、6 d与对照的真菌群落存在显著差异,且本研究中Cr胁迫可能会招募土壤中的致病菌如镰刀菌等使土壤中部分病原菌丰度升高,土壤中可能存在病原菌污染。研究结果进一步表明,Cr胁迫时间序列上土壤中真菌群落的多样性指数在CK阶段最高,在Cr_6 d阶段显著下降,与部分真菌属丰度变化趋势相一致。表明短期的重金属污染降低了土壤微生物群落的丰度与多样性,与Hu等结果相类似。可能因为土壤受重金属污染时,会对土壤中真菌群落产生一定的生理生化毒性,而微生物群落的变化是较为直观的表现形式,因此对Cr抗性较低的真菌群落在胁迫条件下很难存活,丰度呈下降趋势,而土壤真菌群落多样性的减少可能会影响土壤生态系统的结构与功能。此外,重金属对微生物群落的毒性除了与重金属本身的特性相关,还可能与土壤类型和理化性质,如土壤颗粒、湿度、pH、有机质含量密切相关。
3.2 土壤真菌群落构建过程对Cr胁迫时间序列的响应
研究表明,土壤微生物群落的构建由确定过程与随机过程的权衡作用驱动,确定过程涉及非随机和基于生态位的机制,一般包括环境过滤和种间关系; 而随机过程主要表征物种相对丰度的随机变化,一般包括漂变、扩散、迁移和种化作用,但两者的相对贡献可能因环境不同而有所差异。微生物的生物地理学观点认为微生物具有显著的扩散能力与环境适应性,但某些微生物群落存在扩散限制。因此本研究进一步探讨了农田土壤中真菌群落的组装机制,以获得对微生物生态学先进机制的理解。揭示Cr胁迫时间序列上土壤真菌群落的组装过程,有助于了解重金属污染生境中真菌多样性的产生与维持机制,对揭示群落组成和功能的差异具有重要意义。本研究发现,Cr胁迫时间序列上,NTI指数显著先降后升,即呈现出由应激到稳定的状态; 且群落迁移率() 在时间序列上不断增长。随机过程在群落构建中占主导地位,土壤中真菌群落受到的扩散限制逐渐降低,分布愈加广泛,可能原因如下:
一方面,群落构建过程中的随机过程相较于确定过程可产生更多样的生态功能,对驱动农业土壤中真菌群落的组装具有较强的效应。随机过程对可持续农田生态系统中功能产生正向反馈机制,缓解Cr胁迫引起的生态扰动,进而维持农田土壤中生态系统的稳定性。Chen等研究表明,高丰度种群易分散在不同样地而具较高的分布性,而稀有种群由于生态迁移在不同样地中易丢失。土壤真菌群落分布性的增长预示真菌群落中的高丰度物种随Cr胁迫时间而广泛分布,以维持土壤生态网络的稳定性与交互性。因此Cr_6 d阶段的共生网络呈现出较CK、Cr_6 h阶段更复杂的结构特征。土壤中生态系统稳定性与微生物密切相关,真菌群落在时间序列上呈现出由应激到稳定的特征,表明Cr胁迫造成较小程度的生态扰动,随机过程占主导地位从侧面表征了本研究中环境变异较小。可能是土壤中真菌群落存在较高程度的功能冗余,包含有互补型分类单元,其功能互补效应在维持土壤结构与功能稳定中发挥重要作用。真菌群落还可利用植物根系分泌物,如类黄酮、氨基酸等抵抗Cr的入侵,并可通过改变Cr的价态、生物吸附、促进甲基化等机制减少Cr对作物的伤害作用。如镰刀菌属中尖孢镰刀菌()对Cr的生物吸附作用良好,可通过真菌寄生或独立新陈代谢两种方式吸附重金属; 植物也可与微生物联合修复重金属污染,如放线菌与植物联合修复Cr污染土壤。
另一方面,确定过程同样驱动群落的构建。微生物生态网络属性可表征微生物群落对逆境的反应,及相关生态位功能间的关系。在许多生态系统中不同物种之间的相互作用对群落的生态分布格局与群落功能至关重要。本研究中共生网络图表明真菌群落的不同属之间存在显著的相互作用,且不同真菌间绝大多数为正相关,节点数量稳定。表明Cr胁迫的不同真菌类群之间主要表现为互利的相互作用从而提高各自的抵抗力。共生网络中直接与间接相互作用对生态系统起着重要调控作用,它们对生态扰动做出同步反应,并产生了积极的反馈和共同振荡。Cr_6 d阶段的共生网络规模更大、相互作用更复杂,部分原因可能是Cr胁迫后期,土壤真菌群落呈稳定状态,提高了物种活性以及种间相互作用,而复杂的网络状态有利于提升群落的稳定性,进而提高资源转移的效率; 平均路径长度大可减缓真菌群落对Cr胁迫的响应速度,有利于土壤真菌群落有效利用土壤养分,且连通度较高的网络会产生更强的抵抗性以应对环境扰动。本研究中低多样性的真菌群落的随机过程在装配过程中占主导地位。而与这一现象相反的是,之前有研究表明,在对照培养试验中的土壤低多样性细菌群落的确定性过程占主导地位。这种差异可能源于真菌与细菌的体型差异,而体型这种属性与物种的生长速率、群落组装、扩散能力密切相关。
3.3 群落功能对Cr胁迫时间序列的响应
FUNGuild功能预测表明土壤真菌群落以腐生、病理营养型为主。以腐生营养型为主可能与优势菌门子囊菌相关,子囊菌门中多腐生菌,在土壤中可作为分解者分解难降解的有机物从而促进土壤养分循环。病原营养型次之,镰刀菌属、赤霉菌属等病原真菌主要从宿主获取营养,可能对作物产生病害。可考虑通过调控根际微生物群落来抑制病原菌的繁殖。利用抗逆菌等根际微生物制成微生物菌肥或者菌剂以保证‘晋谷21号’良好生长发育,有助于维持土壤的生态平衡和友好环境,对发展可持续农业具有重要的战略意义。
4 结论
1) Cr胁迫时间序列上,土壤真菌群落组成结构显著改变,不同处理组间群落组成差异显著; 胁迫可能影响抗逆菌与敏感菌等不同菌种的丰度,增强了土壤真菌群落对Cr胁迫的适应性; 菌群丰富度显著下降。2) 土壤真菌群落的构建由随机过程主导,随Cr胁迫时间增长群落所受扩散限制减小,分布更广泛; 种间关系以共生为主,提升了群落稳定性以应对生态扰动。3) 土壤中真菌群落多病理营养型、腐生营养型,病原菌含量较高预示了土壤可能同时存在病原菌污染。
通过研究深入了解Cr胁迫下种植过‘晋谷21号’谷子的土壤中真菌群落在时间序列上的动态变化机制,揭示‘晋谷21号’根际土壤真菌的群落构建、互作网络与生态功能,是利用植物微生物组促进‘晋谷21号’作物健康和推进农业可持续发展的基础与前提。
猜你喜欢
农业灾害研究(2022年2期)2022-05-31
山地农业生物学报(2022年3期)2022-05-13
土壤学报(2022年1期)2022-03-08
学校教育研究(2020年7期)2020-04-09
饮食与健康·下旬刊(2019年9期)2019-03-08
大自然探索(2019年1期)2019-01-24
饮食与健康·下旬刊(2018年3期)2018-04-11
知识就是力量(2017年12期)2018-01-15
中学生理科应试(2017年6期)2017-09-27
大自然探索(2017年3期)2017-04-07
