
烟草IMK基因家族的鉴定与表达分析
2022-01-08 09:26陈千思刘萍萍李泽锋杨小宁武明珠郑庆霞周会娜
烟草科技 2021年12期
陈千思,刘萍萍,李泽锋,杨小宁,武明珠,郑庆霞,周会娜
中国烟草总公司郑州烟草研究院,郑州高新技术产业开发区枫杨街2号 450001
类受体激酶(Receptor-like kinase,RLK)是真核生物中保守的蛋白激酶,通过信号转导途径感受植物细胞膜外信号,并将信号传递到植物细胞核[1],参与激活生长[2-3]、发育[4-5]、应激反应[6-8]和抗病[9-11]等过程。RLK 通常由氨基末端的膜定位信号、细胞外的配体结合结构域、跨膜结构域和细胞质激酶结构域构成[12]。对多种RLK 的功能研究表明,配体结合域在感知细胞外的刺激中起着重要作用,而胞质激酶结构域是通过磷酸化下游底物来实现对下游的调控[13]。在模式植物拟南芥中,多项研究表明,RLK 通过多种信号传导途径[14-16]参与调节不同类型的生物学过程[17-21]。其中,一类命名为花序分生组织类受体激酶(Inflorescence meristem receptor-like kinase,IMK),属于LRR 类受体激酶[22]。拟南芥基因组中共有2个IMK 基因,分别为IMK2(AT3G51740)和IMK3(AT3G56100)。拟南芥中,IMK2 蛋白能与另外一种非典型的LRR 类受体激酶SCM 互作并参与根的上皮细胞排布;IMK3 主要在茎顶端和根尖表达,在胚中也能检测到,编码的蛋白质具有体外激酶活性且能与MADS-box 蛋白AGL24 互作并使其磷酸化。
在烟草中,前期研究发现NtIMK 的表达与干旱响应高度相关,暗示IMK 基因除了参与调控生长发育外,还可能参与了对逆境的响应过程[23]。然而,烟草中IMK 蛋白家族的相关研究还未见报道。本研究中利用生物信息学手段分析、鉴定了NtIMK 家族成员,研究了NtIMKs 在不同组织器官和不同激素处理前后的表达模式,旨在为进一步探讨NtIMK 在生长发育及逆境胁迫中的功能提供基础,并为烟草重要性状的分子调控机制研究提供潜在的理论参考和基因信息。
1 材料与方法
1.1 材料、试剂和仪器
材料:栽培烟草品种K326(Nicotiana tabacum L.)种植于国家烟草基因研究中心人工气候室,温度为26~28 ℃,相对湿度60%,光周期为16 h 光照/8 h 黑暗[24]。
试剂:6-苄氨基腺嘌呤(6-Benzylaminopurine,6-BA,Lot No.B3408)、赤霉素(Gibberellin,GA,Lot No.48870)、3-吲哚乙酸(3-Indoleacetic acid,IAA,Lot No.I2886)、茉莉酸甲酯(Methyl jasmonate,MeJA,Lot No.392707)、水杨酸(Salicylic acid,SA,Lot No.S7401)购于美国西格玛奥德里奇公司;独脚金内酯(Strigolactones,GR24,Lot No.41012ES03)购于翌圣生物科技(上海)有限公司;MS Medium(Lot No.MS9043)购于北京密码子生物科技有限公司;Agar(Lot No.A7921)购于美国西格玛奥德里奇公司;植物RNA快速提取试剂盒(Lot No.RN3802)购于北京艾德莱生物科技有限公司;反转录试剂盒Transcriptor First Strand cDNA Synthesis Kit(Lot No.04897030001)和qPCR试剂盒FastStart Essential DNA Green Master(Lot No.06924204001)购于瑞士罗氏公司。
仪器:Biometra Tone 96 PCR 仪器(德国Biometra 公司);LightCycler96 实时定量PCR 仪(Lot No.05815916001,瑞士罗氏公司);移液器(德国艾本德公司)。
1.2 方法
1.2.1 烟草IMK 家族基因的检索和鉴定
在拟南芥种质信息数据库(http://www.arabidopsis.org/)中下载拟南芥IMK2 和IMK3 蛋白质序列。在烟草基因组数据库(ftp://ftp.solgenomics.net/genomes/Nicotiana_tabacum/)普 通烟草K326 的蛋白质序列中进行同源搜索,获得烟草品种K326 的IMK 蛋白质的氨基酸序列。利用TBtools 软 件(https://github.com/CJ-Chen/TBtools/releases)比对获得的NtIMK 蛋白质的氨基酸序列。
1.2.2 烟草IMK 基因亚家族的生物信息学分析
通过MUSCLE 软件对获得的烟草IMK 蛋白和2 个拟南芥IMK 蛋白进行氨基酸序列比对。通过MEGAX 软件采用邻接法生成IMK 蛋白家族的系统进化树,其中,校验参数(Bootstrap)设置为1 000,其余均为默认参数。通过MEME 在线网站(http://meme-suite.org/tools/meme)对IMK蛋白进行保守基序分析。利用在线软件GSDS 2.0(http://gsds.cbi.pku.edu.cn/)对基因结构进行分析,并绘制基因结构分布图。通过TMHMM 软件(http://www.cbs.dtu.dk/services/TMFMM/)对NtIMK蛋白家族进行跨膜结构域预测。
1.2.3 不同组织的表达模式分析
在烟草生长到盛花期时,对第5 叶位叶片、叶芽、第10 叶位叶片、第15 叶位叶片、茎节、茎、侧根、须根、花、花蕾这10 个不同部位组织分别进行取样。样品剪下后,迅速液氮冷冻,-80 ℃冰箱保存。样品经液氮研磨成粉末,使用植物RNA 快速提取试剂盒和反转录试剂盒进行RNA 抽提和反转录,将反转录的cDNA 稀释至400 ng/μL。通过选取基因特异引物,利用qRT-PCR 方法,取1 μL cDNA 用于基因定量分析的模板,用烟草EF1α基因(GenBank ID:NM001326165)作为内参,3 次技术重复。依照qRT-PCR 试剂盒(Lot No.06924 204001,瑞士罗氏公司)说明书上的操作步骤和反应程序,在qRT-PCR 仪器上运行PCR 程序和收集荧光信号。数据结果采用2-ΔΔCT方法进行分析,待测基因的相对表达量参照同一基因在第5 叶位叶片中的表达量。各个组织及处理独立进行3 次生物学重复,每组生物学重复为独立样本并单独提取RNA。
1.2.4 不同激素处理后的表达模式分析
烟草K326 种子用体积分数为70%的酒精浸泡60 s,无菌水清洗2~3 次,然后用质量体积分数为5%的次氯酸钠表面消毒15~20 min(晃荡),无菌水洗3 遍,取出种子放在滤纸上晾干水分,将消毒的种子平铺于MS 固体培养基上(培养基pH=5.8,琼脂浓度0.8%),进行烟草无菌苗培养。待长出幼苗后,将幼苗转移到培养盒中继续无菌培养,直到长到5 叶期时备用。挑取长势一致的烟草无菌苗,用配好的质量体积分数为10%的6 种激素(6-BA、GA、GR24、IAA,meJA 和SA)[25-26]母液分别浸泡无菌苗3 h 和24 h,相应的未处理叶片作为对照,剪下叶片后迅速液氮冷冻,-80 ℃冰箱保存。通过qRT-PCR 方法对6 个NtIMK 基因的表达情况进行分析。RNA 提取、cDNA 的制备、qRT-PCR 体系及反应程序同章节1.2.3。
2 结果与分析
2.1 普通烟草中NtIMK 基因鉴定
为了鉴定烟草中的IMK 基因,利用拟南芥已报道的2 个IMK 类型蛋白IMK2(AT3G56100.1)和IMK3(AT3G51740.1)的氨基酸序列在烟草基因组数据库中比对搜索烟草品种K326 的IMK 蛋白序列,得到6 个编码IMK 蛋白的基因(表1)。氨基酸比对分析结果显示,这6 个IMK 蛋白的氨基酸序列一致性在61%~97%之间。其中Nitab4.5_0011886g 0020.1 与Nitab4.5_0000230g0240.1,Nitab 4.5_00003 75g0060.1 与 Nitab4.5_0003402g0040.1,Nitab4.5_0002854g0020.1与Nitab4.5_0004167g0030.1 两两之间的序列一致性分别高达97%、92%和94%,暗示IMK 基因可能存在功能冗余。
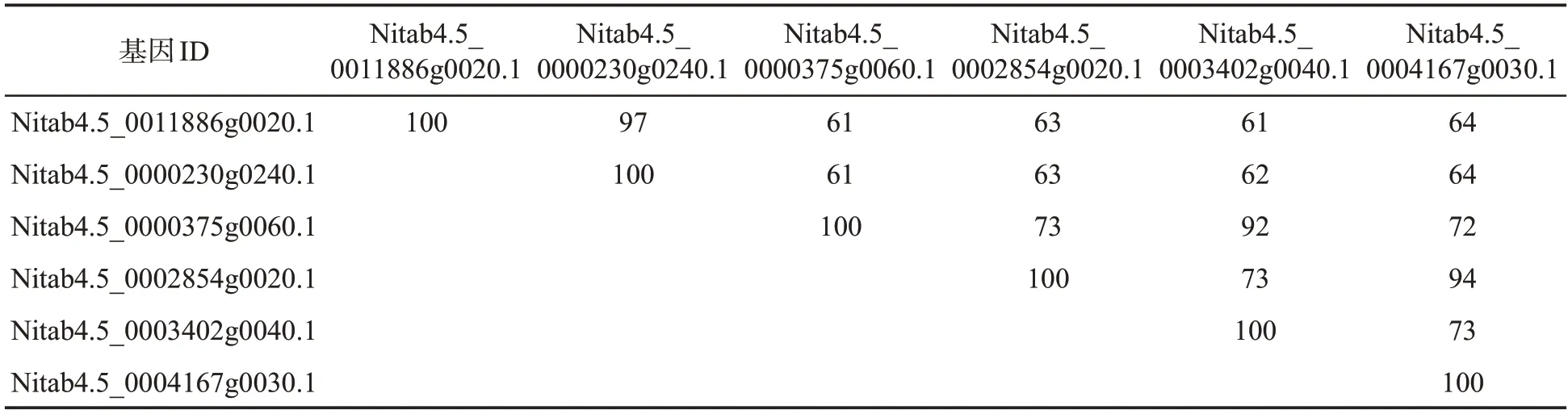
表1 NtIMK 蛋白质的氨基酸序列比对Tab.1 Amino acid sequence alignment of NtIMK protein (%)
2.2 烟草IMK 基因亚家族的聚类、基因结构和motif 分析
使用MUSCLE 软件对6 个烟草IMK 蛋白和2个拟南芥IMK 蛋白进行氨基酸序列比对,在MEGAX 中通过邻接法生成了IMK 蛋白家族的系统进化树(图1A)。由图1A 可见,6 个烟草IMK 蛋白可以分为2 类,其中2 个与拟南芥IMK3 距离较近(IMK3 类),4 个与拟南芥IMK2距离较近(IMK2 类)。基因结构分析显示,这6 个烟草IMKs基因均含有2 个外显子和1 个内含子,且外显子长度较为一致,只有内含子长度变异较大,进一步暗示这些基因可能存在功能冗余(图1A)。
利用MEME 软件对这些IMK 蛋白的保守Motif 进行分析,结果如图1B 所示,共鉴定出12 个分布于IMK 蛋白上的保守Motif,所有IMK 蛋白均包含这12 个保守的Motif。其中不同类型IMK 蛋白的主要区别在于重复motif 12(GSIP motif)个数的差异。烟草IMK2 类蛋白均含有2 个重复的motif 12,而IMK3 类蛋白包含3 个重复的motif 12,暗示motif 12 在这2 类IMK 蛋白的功能分化过程中可能起着重要作用。进一步分析每个motif中氨基酸的保守性(图1C),结果表明motif 10、motif 11 和motif 12 富含Leu,表明烟草IMKs 属于LRR 类受体激酶。

图1 NtIMKs 的进化树(A)、保守基序分布(B)和氨基酸保守性(C)Fig.1 Phylogenetic analysis of NtIMKs (A),distributions of conserved motifs of NtIMKs (B) and amino acid conservation analysis of motifs (C)
2.3 跨膜结构域和翻译后修饰位点分析
通过TMHMM 软件对NtIMK 蛋白家族进行了跨膜结构域预测,结果显示 Nitab4.5_0011886g0020.1 与Nitab4.5_0000230g0240.1 存 在2个跨膜结构域,而其他4 个则可能存在3 个跨膜结构域,说明NtIMK 蛋白也可能是膜蛋白(图2A)。此外,对烟草IMKs 的翻译后修饰位点进行分析,结果表明,在拟南芥IMK2 鉴定出的11 个修饰位点中,仅2 个位点在烟草所有IMKs 蛋白中均存在且保守,包含一个N-glycosylation(天冬酰胺糖基化)和T-phosphorylation(苏氨酸磷酸化)。类受体激酶通常通过自身磷酸化状态的变化来行使功能,因此推断这2 个位点可能在IMKs 蛋白发挥功能过程中起着重要作用,其中苏氨酸磷酸化位点可能是其激酶活性位点(图2B)。

图2 NtIMK 蛋白质的跨膜结构域(A)和翻译后修饰位点(B)分析Fig.2 Analysis of transmembrane domains (A) and modified sites after translation (B) in NtIMK proteins
2.4 不同组织表达模式分析
通过选取基因特异引物(表2),利用qRT-PCR的方法对这些基因在叶、茎、茎节、花、花蕾、侧根和须根中的表达情况进行了分析(图3)。结果发现,6 个IMKs 基因家族成员均在侧根和花蕾中高表达,其中Nitab4.5_0002854g0020、Nitab4.5_0000375g0060 和Nitab4.5_0003402g0040 在花蕾表达最高,Nitab4.5_0004167g0030、Nitab4.5_0011 886g0020 和Nitab4.5_0000230g0240 则在侧根中的表达高于其他组织。Nitab4.5_0011886g0020 和Nitab4.5_0000230g0240 除了在氨基酸序列高度相似外,在不同组织中表达模式相似,除了在花蕾、茎和侧根中高表达外,在茎节、第10 叶位和第15叶位叶片中也高表达,暗示其除了具有花蕾和侧根的发育功能外,可能还在叶和茎中发挥功能。另外一对高度同源的基因对Nitab4.5_0000 375g0060 和Nitab4.5_0003402g0040 在不同组织中表达模式相似,均在侧根中表达最高,在第15 叶位叶片中表达量最低。表达模式分析结果说明,不同的烟草IMKs成员表现出不同的组织特异性,并且氨基酸序列一致性高的基因其表达模式也较为相似。

表2 引物序列及用途Tab.2 Sequences of primers and their usage
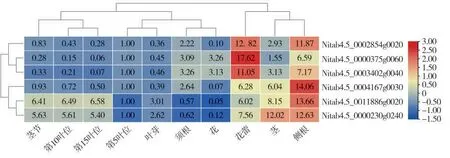
图3 NtIMKs 在不同组织的表达模式Fig.3 Expression patterns of NtIMKs in different tissues
2.5 不同激素处理后的表达模式分析
烟草IMKs 基因家族成员对6 种激素(包括6-BA、GA、GR24、IAA、meJA 和SA)响应的分析结果见图4,与对照相比,GR24 和SA 处理后Nitab4.5_0011886g0020 的表达量显著上调;GA、GR24 和IAA 处理后,Nitab4.5_0000230g0240 的 表达量显著上调;GA 处理后,Nitab4.5_0000375g0060的表达量显著上调,但在6-BA 和meJA 处理后该基因的表达量显著下调;GA、IAA 和SA 处理后,Nitab4.5_0002854g0020 的表达量显著上调;GA 和IAA 处理时,Nitab4.5_0003402g0040 的表达量显著上调,但在6-BA 处理时,该基因的表达值显著下调;GA、IAA 和SA 处理后,Nitab4.5_0004167g0030的表达量显著上调。
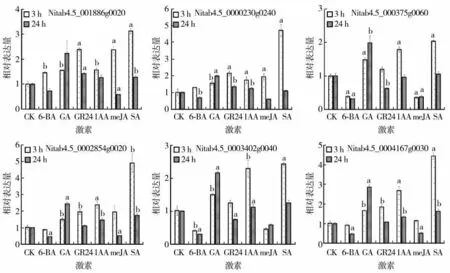
图4 不同激素处理后烟草IMKs 的表达模式Fig.4 Expression modes of NtIMKs treated by different hormones
3 讨论
拟南芥中最大的RLKs 家族LRR RLK 大约有235 个成员,在发育、病原体抗性和激素感知中具有多种作用。研究表明,LRR RLKs 在与激素和非生物应激反应有关的多种信号转导途径中发挥作用。ABA 是非生物胁迫响应的关键介质,可以调节干旱、盐和渗透胁迫反应基因的表达。拟南芥RPK1 基因(Receptor-like protein kinase 1 gene)突变体对ABA 不敏感,且干旱胁迫响应基因表达大幅下调。油菜素内酯(BRs)是植物必需的多羟基化甾体类激素,对植物生长发育至关重要。被命名为第一个BR 信号转导基因的BRI1 基因(Brassinosteroid insensitive 1 gene)与其相关受体激酶BAK1 异源二聚化而参与BR 的感知和信号传导。BAK1 相关受体激酶1(BARK1)的过表达促进了根系的生长,表明BARK1 在BR 介导的根系发育中发挥作用[1]。BAK1/SERK3 受IAA 和邻氨甲酰苯甲酸(NPA)调控,RLK902 受NPA 调控,但IMK2 和IMK3 不受激素调节,IMK2 受NPA 和甘露醇调节,IMK3 受盐和甘露醇调节[27]。拟南芥中,IMK2 蛋白能与另外一种非典型的LRR 类受体激酶SCM 互作并参与根的上皮细胞排布;IMK3 主要在茎顶端和根尖表达,在胚中也能检测到,编码的蛋白质具有体外激酶活性且能与MADS-box 蛋白AGL24 互作并使其磷酸化[28]。
利用序列分析、序列比对等生物信息学相关方法,从普通烟草基因组中鉴定出了6 个IMK 家族成员。这些烟草IMK 基因结构和其编码的蛋白质序列均相对保守,主要分为2 类,一类与拟南芥IMK2 相近。激素处理后的基因表达模式分析结果显示,大多数烟草IMKs 能响应不同的激素,其中4 个烟草IMKs 均被GA 和IAA 诱导上调表达,表明Nitab4.5_0000230g0240、Nitab4.5_0002854g0020、Nitab4.5_0003402g0040 和 Nitab4.5_0004167g0030均可能参与了GA 和IAA 介导的调控反应。Nitab4.5_0000375g0060还被6-BA和meJA抑制下调表达。组织表达模式分析结果说明,大部分的烟草IMKs 在侧根和花蕾中呈现高表达,这与拟南芥中IMK2 和IMK3 较为相似,暗示烟草IMKs 基因可能在侧根和花发育过程中起关键作用,不同的烟草IMKs 成员也表现出不同的组织特异性,同源基因对Nitab4.5_0000375g0060 和Nitab4.5_00034 02g0040 在不同组织中表达模式相似,均在侧根中表达量最高,在15 叶位烟叶中表达量最低,表明这些基因在烟草中可能存在功能冗余。表达模式分析结果表明,不同类型的IMKs成员表现出不同的组织特异性,表明它们可能在烟草不同组织发育或逆境响应过程中发挥重要作用。IMK 可能负调控GA 和IAA 激素介导的生理反应,但这些烟草IMKs如何参与激素介导的生理反应,还有待深入研究。
4 结论
在普通烟草基因组中共鉴定出6 个IMK 家族成员,鉴定出的IMKs 大多数可以响应不同的激素。其 中,Nitab4.5_0000230g0240、Nitab4.5_0002854g 0020、Nitab4.5_0003402g0040 和Nitab4.5_0004167g 0030 均被GA 和IAA 诱导上调表达,表明这4 个基因均可能参与了GA 和IAA 介导的调控反应。此外,6 个IMKs 基因家族成员均在侧根和花蕾中高表达,表明IMKs 基因可能在侧根和花发育过程中发挥关键作用。
猜你喜欢
农业现代化研究(2022年3期)2022-06-29
天津医科大学学报(2021年3期)2021-07-21
世界科学技术-中医药现代化(2021年12期)2021-04-19
国际放射医学核医学杂志(2020年4期)2020-07-27
生命科学研究(2018年1期)2018-05-29
上海农业学报(2017年3期)2017-04-10
中国医药生物技术(2015年4期)2015-12-26
植物营养与肥料学报(2014年1期)2014-03-11
中国医学科学院学报(2013年6期)2013-03-11
植物营养与肥料学报(2011年4期)2011-10-26
