
基底核在类别学习中的纽带作用
2022-01-08 13:36卡罗尔塞格CarolSeger陈剑平王宇情刘志雅
华南师范大学学报(社会科学版) 2021年6期
卡罗尔·塞格(Carol A. Seger), 陈剑平, 王宇情, 刘志雅*
(1.华南师范大学 心理学院, 广州 510631;2. 科罗拉多州立大学 心理学系,美国科罗拉多州柯林斯堡 80523)
类别学习(category learning)是人类通过学习把纷繁复杂的世界知识进行有序的组织。类别学习的实验范式通常会预先创设某个新的类别,例如一类新的物种或者几何图形,然后让被试进行分类学习。在每个试次中,一般先呈现某个类别的样例,要求被试判断该样例属于A类还是B类,然后对其判断的正误给予反馈。这样,通过多次刺激反应的学习,直到被试达到预先设定的学习标准,被试就习得了一个新的类别知识。最后,通过迁移探测,了解被试学习后形成的类别表征,或者记录被试的神经生理数据,探索类别学习的神经机制[1-2]。
一、类别学习的认知与神经研究
当前,关于人们如何表征类别知识,研究者们已经提出了一些较为成熟的认知模型[3],如规则模型(rule-based theory, RT)、原型模型(prototype-based theory, PT)和样例模型(exemplar-based theory, ET)。原型模型认为[4-6],类别学习实质上是学习类别的原型。当遇到一个新的样例时,哪个类别的原型与之更相似,人们就会把它归类到这个类别中。样例模型认为[7-9],类别学习的实质是学习类别中的每个样例。当遇到一个新样例时,哪个类别有更多与之更相似的样例,人们就会把它归类到哪个类别之中。规则模型认为[10-12],类别学习的实质是习得决定性的特征,即“规则”,当某个新样例符合这个“规则”时,人们就会把它归类到这个类别中。
当前,对三个主要认知模型的争论仍然激烈,近年来随着认知神经科学的兴起,研究者们纷纷开始寻求神经生理的数据来进行验证。然而,由于类别学习是一个高级的心理过程,涉及的神经机制具有层次深、范围广的特点,因此一直未能取得一致的观点。首先在认知与神经机制两方面获得较大进展并形成系统性观点的,是以阿什比(Ashby)和马多克斯(Maddox)为首提出的“多重类别学习系统”方面的研究。他们主张类别学习具有两个不同的学习系统,分别是基于规则的系统和信息整合的系统[3, 13]。基于规则的类别学习指的是学习者学习一种可以用言语表述的规则。例如,让学习者分类学习一些光栅图,具体的刺激如图1中的(1),学习的材料结构有(2)、(3)、(4)三种。对于图 1中的(2)类别结构材料,学习者会习得一种单维度规则,即“所有光栅密度疏的图形都是A类,密度密的图形都是 B 类”;对于图 1 中的(3)类别结构材料,学习者会习得一种双维度规则,即“所有光栅密度疏且倾斜较陡的图形都是A类,除此之外的图形都是B类”。这两种都是基于规则的类别学习,它可以是单维度规则,也可以是多维度的联合规则,关键是能否使用言语把规则描述出来。另外一种是信息整合的类别学习,指学习者学习一种难以用言语表述的规则,如图1中的(4)。学习者虽然能够很好地对两类刺激进行分类,但无法用言语来描述分类规则[2]。他们同时指出,基于规则的类别学习是一种外显学习,容易受工作记忆和执行注意的影响;而信息整合的类别学习是一种内隐的、自动化的学习,不容易受工作记忆和执行注意的影响。阿什比和马多克斯提出的基于规则和信息整合的类别学习任务成了当前探索类别学习认知神经机制最典型的任务范式之一,并引发了大量的临床和脑成像的研究[14-18]。
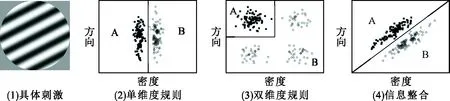
图1 基于规则和信息整合的类别学习材料
关于外显类别学习和内隐类别学习的神经机制的探索取得了两方面的研究进展[19]。第一,研究指出前额叶皮层(prefrontal cortex)在外显类别学习中的重要性,如起到选择、检验和转换规则,忽略无关规则等作用。雷伯(Reber)等人探讨了点模式任务下的脑活动,发现基于规则的类别学习显著地激活了前额叶,前额叶是类别学习中进行抽象活动的重要神经系统[20]。许多研究表明,前额叶损伤患者和帕金森氏症病人在基于规则的类别学习方面受损[21-22]。几乎所有用于神经心理评估的分类学习任务都是基于规则的,包括广为人知的威斯康辛卡片分类测验(Wisconsin card sorting test, WCST),在WCST中重复出错是额叶功能障碍的典型症状[23]。第二,研究指出基底核(basal ganglia)在内隐类别学习中的作用,如起到将感知空间的区域与特定的运动反应联系起来等作用。诺姆拉(Nomura)等人比较了基于规则和信息整合类别学习的认知神经机制,结果发现,相比基于规则学习,信息整合学习与尾状核身体(caudate body,基底核重要组成部分之一)的激活有关[14]。菲洛蒂奥(Filoteo)等人发现,基底核损伤的病人,如帕金森氏症病人和杭廷顿氏症病人,在非线性的信息整合任务中的表现不如控制组[24]。
随着研究的进一步深入,研究者发现,基底核通常在涉及试错的类别学习任务中都会激活。在这些任务中,被试需要观看一个刺激,判断该刺激属于哪个类别并作出反应,然后收到反馈。因此,基底核在类别学习中的作用引起了研究者的重视[25-30]。
二、基底核和皮质—纹状体回路
塞格(Seger)和米勒(Miller)的综述指出[31],类别学习依赖于多种神经系统,而不是只依赖于单一神经系统。类别学习涉及的脑功能区包括前额叶皮层、视觉皮层(visual cortex)、顶叶皮层(parietal cortex)、运动皮层(motor cortex)等大脑皮层区域,还包括基底核、内侧颞叶(medial temporal lobe)和中脑多巴胺能系统(midbrain dopaminergic systems)。在类别学习的整个过程中,包括知觉刺激、特征识别、规则形成、决策判断、规则转换、归纳推理、规则使用和知识迁移等多个过程。这些不同的认知过程引发了不同脑区的活动,激活了多个神经系统间的连接。不同的类别学习任务类型激活不同的神经系统,而这些神经系统在类别学习过程中相互作用。他们进一步归纳指出,连接大脑皮层和基底核的皮质—纹状体回路(corticostriatal loops)在类别学习过程中扮演着非常重要的角色。
基底核是大脑皮层下一系列神经核团的集合,与大脑皮层、丘脑和脑干相连。基底核主要包括纹状体(striatum)、苍白球(globus pallidus)、丘脑下核(subthalamic nucleus, STN)和黑质(substantia nigra)。其中,纹状体分为尾状核(caudate nucleus)、壳核(putamen)和腹侧纹状体(ventral striatum),腹侧纹状体有时也叫伏隔核(nucleus accumbens, NAcc)。苍白球分为内侧部(globus pallidus internal segment, GPi)和外侧部(globus pallidus external segment, GPe)。黑质分为网状部(substantia nigra pars reticulata, SNr)和致密部(substantia nigra pars compacta, SNc)[32]。
几乎所有的大脑皮层都投射到基底核的输入结构,这些输入结构包括纹状体和丘脑下核;然后,纹状体和丘脑下核投射到基底核的输出结构,这些输出结构主要包括苍白球和黑质网状部;接着投射到丘脑,最后再投射回大脑皮层,这样就构成了皮质—纹状体回路[33-34]。不同的纹状体区域接受不同大脑皮层的投射,虽然有部分脑区重叠,但是皮质—纹状体回路可以分为四条独立的回路,分别是视觉回路(visual loop)、运动回路(motor loop)、执行回路(executive loop)和动机回路(motivational loop),模式图见图2[35]。从模式图中可以看出,基底核在每条回路中都像纽带一样连接着大脑皮层和丘脑,维系着四条回路的稳定运行。这四条回路在类别学习中起一定的作用,很大程度取决于该回路的大脑皮层区域。根据具体的大脑皮层区域,可以推测皮质—纹状体的四条回路在类别学习的知觉识别、反应选择、规则形成和转换、反馈处理等过程中分别承担重要的作用。
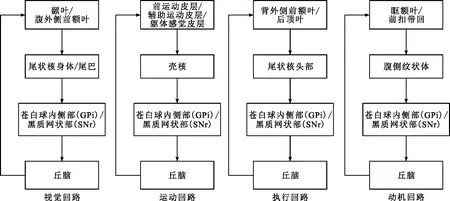
图2 皮质—纹状体回路
第一,视觉回路。这条回路通过尾状核身体和尾部(caudate tail)连接纹外视觉皮层(extrastriate occipital cortex)和颞下皮层(inferior temporal cortex,ITC), 研究表明这条回路可能在类别学习中的刺激知觉和特征识别等视觉信息处理上承担了重要作用[36]。形状知觉归类任务的功能性磁共振成像研究(如原型变形任务)表明,纹外视觉皮层在早期的刺激外部特征变化时就首先被激活了,尤其是BA 18和19区域[37-38]。而颞下皮层与类别的抽象泛化过程无关,与物体特征的高级分析过程有关,对类别间外部特征的知觉差异作出特定的反应。对猴子微电流刺激的研究发现,颞下皮层能够促进对新异图片的视觉归类[39]。此外,德古蒂斯(DeGutis)等的人脸识别研究也指出,颞下皮层下的梭状回(tusiform gyrus)在人脸分类活动中有重要的作用[40]。塞格等人研究发现,尾状核的身体和尾部在视觉分类过程中被激活,其活动与反馈接收无关,与正确分类类别成员有关[41-42]。尾状核身体和尾部的位置接近视觉皮层,并与视觉皮层建立了广泛的联系,我们推测它们与类别学习的特征识别机制有较大的关系。
第二,运动回路。这条回路将前运动皮层(premotor cortex, PMC)、辅助运动皮层(supplementary motor area, SMA)、躯体感觉皮层(somatosensory cortex)与壳核连接起来,研究表明这条回路在类别学习中选择适当行为的反应和自动化上起到了重要作用[43]。在类别学习初期,选择适当行为的反应不仅需要视觉回路将视觉信息输出到早期运动计划脑区,如前辅助运动皮层(pre-supplementary motor area, pre-SMA),而且需要运动回路在前运动皮层和初级运动皮层中选择更具体的运动程序,即选择正确的类别。为了验证这种可能性,塞格等人的研究发现,在分类学习过程中,壳核与尾状核的身体和尾巴同时活跃[41]。随着类别学习过程的深入,刺激反应慢慢成为熟练的技能,前额叶的激活程度逐步下降,转而依靠前运动皮层作出判断,而且壳核变得越来越重要[35]。这些结果表明,运动回路在适当行为的反应选择上发挥着重要作用,并且与默认网络一起共同维持持续性的反应动作,进而形成行为习惯,达到自动化水平。
第三,执行回路。这条回路是通过尾状核头部(caudate head)连接前额叶皮层和顶叶皮层,研究表明这条回路在类别学习中对规则的形成和转换承担了重要作用[44-46]。诺姆拉等人认为,前额叶皮层在复杂的抽象活动中形成了规则,然后从纹状体中获得反馈性的信息来调整或替换原有的规则。而且,规则学习中的刺激变换可以引起腹侧纹状体及尾状核头部的激活, 所需的样例存储功能使海马(hippocampus)和内侧颞叶也参与到学习的活动中来[47]。塞格等人考察了基于规则学习任务下额叶、纹状体和海马的交互情况,结果发现,在规则学习过程中,内侧颞叶与纹状体系统之间存在拮抗效应。在学习前期,内侧颞叶可能调动长时记忆系统资源对新异信息进行编码,而当基底核开始起作用后,内侧颞叶的作用逐渐下降[42]。尾状核头部不仅对反馈敏感,而且能检测到行为情景的变化,允许前额叶皮层更新工作记忆[48]。COVIS模型认为,基于规则的学习由一系列神经网络调节,其中包括前额叶皮层、前扣带回、尾状核头部和海马[3, 13, 19]。这些结果表明,皮质—纹状体回路在规则学习和记忆上可能对相关信息起了上传下达的枢纽性作用。随着功能性磁共振成像研究新技术的发展,特别是与学习相关的脑网络,如认知控制网络(cognitive or executive control network)、突显网络(salience network)等研究的深入,将更有力地揭示基底核在类别信息加工中的重要作用。
在视觉回路和执行回路中,尾状核的各部分存在功能分离,即尾状核身体和尾部与视觉信息处理有关,尾状核头部与规则形成和转换有关。尾状核是基底核的重要组成部分之一,和大脑的各部分形成了广泛的联系。探索尾状核不同部位在类别学习中的不同作用,也将更系统地揭示基底核在类别信息加工中的重要作用。
第四,动机回路。这条回路连接腹侧纹状体和眶额皮层(orbitofrontal cortex)、前扣带回(anterior cingulate cortex, ACC),研究表明这条回路在类别学习中对反馈和奖赏的处理承担了重要作用[35]。腹侧纹状体与中脑多巴胺系统有着大量神经联结,前者的激活被认为与错误反馈和奖赏的加工有密切的关系[25, 49]。格兰班德(Grinband)等人考察了在分类边界可变化的类别学习任务中与分类相关的脑区,结果发现,由于难以感知关键特征,腹侧纹状体对正确类别的不确定性非常敏感[50]。来自其他认知领域的研究表明,腹侧纹状体和尾状核头部可能是可分离的,这取决于反馈和奖励各自反映的方面[35]。腹侧纹状体和尾状核头部都对反馈信息敏感,但研究表明前者对学习中的错误反馈信息敏感,而后者则对学习中的奖赏信息敏感[16]。塔纳卡(Tanaka)等人考察了马尔科夫决策任务中不同时间尺度下的奖励预测大脑机制,他们发现,奖赏时间对腹侧纹状体与尾状核头部存在不同的影响,前者对及时强化非常敏感,而后者则对延迟满足表现出敏感反应,这表明二者加工信息的类型和时程存在差异[51]。
皮质—纹状体的四条回路在类别学习中各自起到不同的作用,并且它们之间是交互的,不是独立的,这样才能产生有意义的认知或行为输出。塞格与其同事使用格兰杰(Granger)因果模型检验了分类过程中皮质—纹状体回路之间的相互作用,发现从视觉回路到运动回路,以及从运动回路到执行回路的直接影响模式相一致。该模式与经典类别学习任务的每一步所需过程相一致,分别是加工视觉刺激、准备和执行表明类别归属的运动反应、接收和处理反馈[52]。当人们从一个分类任务中的新手变成专家时,皮质—纹状体回路也会在许多经验或试次中相互作用。执行回路和动机回路在早期是最重要的,此时获取信息的速度最快,反馈处理最有用,而运动回路的重要性随着专业知识的获得而提高[31]。威廉姆斯(Williams)等人让恒河猴完成联想学习任务,通过单细胞记录方法记录恒河猴的纹状体活动情况。结果发现,尾状核头部(执行回路)对学习速度敏感,当学习发生迅速[53]以及预测误差最大时[54],它的激活程度达到最大。相比之下,在掌握类别知识之后,壳核(运动回路)在学习后期变得更活跃[53]。
综上所述,类别学习的神经机制可能是以大脑的基底核为纽带,接受大脑皮层的信息输入,并将信息输出给丘脑,最后回到大脑皮层,构成皮质—纹状体回路,并以皮质—纹状体四条回路为基础实现类别学习所需的各个方面。此外,皮质—纹状体四条回路并不是独立的,而是相互作用的。
三、总结和展望
随着研究的深入,基底核在类别学习中的作用越来越受研究者的重视,并取得了一定的成果。基底核与大脑皮层相互作用形成皮质—纹状体回路,参与类别学习的各个方面,包括知觉识别、反应选择、规则形成和转换、反馈处理等。在整个类别学习的进程中,每条皮质—纹状体回路并非独立运作,而是相互作用以支持分类。例如,在一个典型类别学习任务的试次中,被试需要观察一个刺激,作出反应,以及接受反馈。这个任务的每个方面可能都需要皮质—纹状体回路的参与。首先,观察刺激激活了视觉皮层区域,这些视觉区域投射到视觉回路的尾状核身体和尾部;然后,视觉回路将输出发送到额叶的运动计划区域,与运动回路一起帮助选择适当的分类反应;最后,接收到的反馈由执行和动机回路中的尾状体头部和腹侧纹状体处理,用于更新关于刺激类别的记忆表征,以便在未来的试次中使用。
学习是一个高级的心理过程,涉及人类大部分的认知过程,如注意、知觉、记忆、思维等,甚至情绪和意志也可能参与进来,以至于难以形成一个统合性的理论观点。刘志雅等主张从学习条件、学习过程和学习结果三方面来系统地解释类别学习的认知机制[1]。探索基底核在上述三个方面起到的作用,将有利于检验以基底核为纽带,皮质—纹状体的四条回路为实现途径的神经机制。
类别学习的条件指的是影响类别学习内部信息加工过程的外部因素。近年来,有研究者从学习的过程,特别是通过控制信息的叠加来研究类别学习的神经机制[16, 28]。在动物研究中也发现,循序渐进的学习方式比尝试错误学习有更高的学习效率,使用渐进学习方法来训练发育迟滞的儿童获得了显著的效果。因此,研究学习材料的不同安排对学习效果的影响,结合高空间分辨率脑成像设备和新的数据分析方法,探索基底核在调整学习条件上的作用,预期能够为人类学习机制带来有价值的研究结果。
类别学习的过程指的是学习者内部信息加工过程,如采用了什么学习策略,信息处理具有什么特点,以及学习的效率如何等。学习策略是实现高效率学习的保障。一些研究者提出了类别学习中存在三种学习策略:其一是单维度规则策略,即寻求某个标准特征来对所有的样例进行归类;其二是规则加例外策略,寻求标准特征来对绝大多数的样例进行归类,同时记住某些例外的样例,最后完全归类正确;其三是信息整合策略,通过对维度规则的整合或特征的叠加,即前者形成联合规则或后者抽象出原型,进而对所有的样例进行正确归类[1, 55-56]。这些特点包括采用了什么学习策略、在学习的早期还是晚期出现、策略间如何转变等,这些问题的研究有重要的理论和实践价值。一些研究也表明,在类别决策界限附近的刺激对大脑有更明显的激活[40]。综上,研究类别学习过程中采用的学习策略,结合高时间分辨率的脑电设备和新的数据分析方法,可以探索学习策略在发展上呈现的阶段性特点,可以发现人类高效率学习的机制。
类别学习的结果一方面指的是学习者学习后形成了什么样的类别表征, 是抽象出原型还是以样例的表征方式来存储。近年来, 研究者倾向采用两种不同学习任务的比较范式来探索其形成的类别表征[57-58]。学习结果的另一个方面,是考察学习者学习后对类别表征的迁移,它反映了学习者掌握知识的巩固程度和对知识的运用能力[59-60]。综上,研究不同类别结构下学习者形成的类别表征和迁移,结合各种已有的或新设计的任务范式,应用认知神经研究方法和技术,可以发现人类学习表征的脑机制。
基底核的不同部位在功能上具有一定的独立性,这对承担学习这项复杂的认知活动提供了一定的解剖基础[32-33, 61]。生理学倾向的研究注重在皮质—纹状体的四条回路上探索学习机制,心理学倾向的研究注重在特征识别、注意策略、刺激反应和反馈上探索学习机制。本文从信息加工的角度出发,主张未来研究可以从学习条件、学习过程和学习结果上研究类别学习的认知机制,并以此进一步探索基底核在类别学习中的作用。
猜你喜欢
中华实验眼科杂志(2022年6期)2022-11-15
中国体育科技(2022年1期)2022-03-10
中国康复(2021年6期)2021-11-30
青岛大学学报(医学版)(2021年5期)2021-11-17
广东教学报·教育综合(2020年15期)2020-03-23
首都体育学院学报(2019年5期)2019-10-18
知识文库(2017年9期)2017-10-20
中国当代医药(2017年17期)2017-07-25
支部建设(2016年18期)2016-11-28
中学教学参考·文综版(2014年1期)2014-03-11
