
基于GC-MS分析不同品种小米代谢产物及代谢途径
2022-01-06 05:01于润众宗恩祥张丽媛张东杰
食品科学 2021年24期
于润众,宗恩祥,张丽媛,,张东杰,3,4,
(1.黑龙江八一农垦大学信息与电气工程学院,黑龙江 大庆 163319;2.黑龙江八一农垦大学食品学院,黑龙江 大庆 163319;3.黑龙江省农产品加工与质量安全重点实验室,黑龙江 大庆 163319;4.国家杂粮工程技术研究中心,黑龙江 大庆 163319)
杂粮是具备“天然、绿色、营养、健康”品类特征的一类粮食。杂粮营养丰富[1],其营养价值大多优于主粮(小麦和水稻)。当前,我国政府实施的“健康中国”战略为健康食品市场发展提供了十分有利的政策机遇和空间,杂粮作为很好的营养素来源也是未来健康食品的主力军[2-3]。随着我国居民膳食要求的安全化向营养均衡的深入,对“杂粮”食品的要求也更“深入化、精细化、营养均衡化”[4]。小米(Setaria italicaL.)是杂粮的一种,是北方广泛种植的杂粮作物之一,它具有一定的保健功能,深加工后能生产出易消化、高营养、高附加值的产品[5-6]。
代谢组学是研究生物体内内源性代谢产物及其变化规律[7-8]的一门科学。代谢组学在食品中的应用主要是研究食品代谢成分[9-10],日渐成为探索食品独特品质最方便的方法之一。气相色谱-质谱结合代谢组学技术已被广泛应用于食品、医药、化工等各个领域[11],如大米代谢产物[12]的产地鉴别、绿豆[13]代谢产物分离鉴定等。
本实验利用气相色谱-质谱(gas chromatography-mass spectrometry,GC-MS)联用技术分离鉴定同一产地2 个不同品种小米的代谢产物,分析2种小米的差异代谢产物和各自的优势代谢产物,进一步探讨代谢途径的差异。本研究旨在为小米的选育和品种改良提供参考,同时相应数据也可作为小米产品精深加工利用的依据。
1 材料与方法
1.1 材料与试剂
在肇源县古龙镇进行的田间试验获得小米样品,品种分别为古龙贡米、禾绅小米,在相同田间管理条件下同时生长,同时收获,收获后储藏在-80 ℃冰箱备用,3 个生物学重复用于代谢产物测定。
N,O-双(三甲基硅基)三氟乙酰胺 美国麦克林公司;甲氧胺盐酸盐(分析纯)、吡啶(色谱级,纯度≥99.9%) 美国Sigma-Aldrich公司;甲醇、乙腈、异丙醇(均为色谱级) 美国赛默飞世尔科技公司;代谢产物标准物质 美国Sigma-Aldrich公司和国家药品和生物制品控制研究所(北京,中国);色谱级用水由美国米利波尔公司的Milli-q水净化系统制备。
1.2 仪器与设备
TQ8040三重四极杆型GC-MS联用仪(配备电子电离源)、AOC-5000自动进样器 日本岛津技术有限公司;CR3i multifunction型离心机、移液枪(1 000 μL和200 μL) 美国赛默飞世尔科技公司;DGG-9140A型电热恒温鼓风干燥箱、DRP-9082型电热恒温培养箱上海森信实验仪器有限公司;MTN-2800D氮吹浓缩装置天津奥特塞恩斯仪器有限公司。
1.3 方法
1.3.1 小米样品预处理
选取完整、成熟的小米样品,放入研钵研磨,研磨好的粉末样品储存于-80 ℃冰箱待分析。
1.3.2 代谢产物提取
分别称取50 mg粉末小米样品于2 mL EP管,加入800 μL 80%甲醇溶液和10 μL内标(2-氯苯丙氨酸)移入EP管快速涡旋30 s混匀,均质放入超声波清洗机35 ℃超声9.0 min,超声提取过程每1 min剧烈振摇1 次,而后置于4 ℃离心机,12 000 r/min离心10 min,离心后取200 μL上清液转移至GC进样小瓶(1.5 mL自动进样瓶),氮吹。
1.3.3 衍生化处理
取30 μL甲氧铵盐酸吡啶溶液至浓缩后的样品中,快速混匀至完全溶解,置于37 ℃恒温箱1 h,取出后加入30 μL三氟乙酰胺在70 ℃烘箱1 h进行衍生化处理。
1.3.4 GC-MS分析
色谱条件:用自动取样器注入1 μL样品溶液。进样口温度280 ℃,四极杆温度150 ℃,高纯氦气(纯度大于99.999%)作为载气,不分流进样,进样量1.0 μL。升温程序:初始温度80 ℃,维持2 min,10 ℃/min升至320 ℃,并维持6 min,然后80 ℃平衡6 min,然后再注入下一次样品。
质谱条件:电子电离源;离子源温度230 ℃;接口温度300 ℃;溶剂切割时间2 min;采集模式Q3 Scan,采用全扫描。
1.4 数据处理
实验结果与美国国家标准与技术研究所(National Institute of Standards and Technology,NIST)标准谱库对比分析,得到的代谢产物信息在京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)数据库[14]中搜索比对,确认代谢产物和代谢途径,运用Excel制表,采用Origin绘图。
2 结果与分析
2.1 代谢产物的GC-MS分离与鉴定
如图1、2所示,由代谢谱图中分离度、分辨率以及基线的稳定性看,本次实验分离鉴定方法的条件选择有效,两谱图的总离子流图大致相似,但是在25.0~27.0 min时间段存在不同。

图1 古龙贡米的GC-MS总离子流图Fig.1 GC-MS total ion current chromatogram of extracts from‘Gulong’ millet

图2 禾绅小米的GC-MS总离子流图Fig.2 GC-MS total ion current chromatograms of extracts from‘Hesen’ millet
其他数据经NIST标准谱库对比分析,确定了代谢产物的结构,分析得出2 个不同品种的小米共有代谢产物53种,如表1所示,其中古龙贡米代谢产物占48种,禾绅小米代谢产物占32种。可将代谢产物分为5 类:有机酸、脂肪酸、糖及其衍生物、氨基酸及其衍生物和中间代谢产物。在被鉴定的全离子分析物中,乙酰酸、N-甲基丙酰胺、乙胺、巴豆醇、叔丁醇、硅醇以上化合物为衍生化试剂形成,在此不作讨论。

表1 2种小米中的代谢产物Table 1 Metabolites identified in two different varieties of millet

续表1
2.2 2 个品种小米相同代谢产物分析
从图3可以看出,相同代谢产物中古龙贡米含量较高(>10%)的有亚油酸;相对含量中等(2.5%~10%)的有甘油、丙酸酯、棕榈酸、油酸、正颉氨酸、醋酸盐;相对含量较低(<2.5%)的有β-谷甾醇、甘油亚油酸酯、硬脂酸、2-亚麻酰基-rac-甘油、磷酸、D-葡萄糖、花生酸、D-山梨醇、乳酸、2,2-二甲基乙酰胺、氨基甲酸酯、氨基甲酸二乙酯、苹果酸、D-吡喃半乳糖、α-D-吡喃半乳糖、D-呋喃果糖、L-谷氨酸、木糖醇、蔗糖。

图3 2种小米相同代谢产物的相对含量Fig.3 Relative contents of shared metabolites in two varieties of millet
禾绅小米中相对含量较高(>10%)的有甘油、亚油酸;相对含量中等(2.5%~10%)的有油酸、棕榈酸、丙酸酯、正颉氨酸;相对含量较低(<2.5%)的有硬脂酸、D-山梨醇、醋酸盐、2-亚麻酰基-rac-甘油、β-谷甾醇、甘油亚油酸酯、花生酸、磷酸、氨基甲酸酯、乳酸、氨基甲酸二乙酯、D-葡萄糖、D-吡喃半乳糖、2,2-二乙基乙酰胺、β-D-半乳糖、D-呋喃果糖、苹果酸、L-谷氨酸、α-D-吡喃半乳糖、木糖醇、蔗糖。
其中古龙贡米的优势代谢产物(相对含量>15%)为亚油酸,禾绅小米的优势代谢产物(相对含量>15%)为亚油酸;古龙贡米中2-亚麻酰基-rac-甘油相对含量高于禾绅小米;禾绅小米中甘油相对含量高于古龙贡米亚油酸可能经过脂肪水解产生或经乙酰CoA和乙酰CoA羧化酶的作用合成,其含量高可能受栽种地气候和环境因素影响代谢途径中酶的活性等原因。有研究表明到目前为止已发现共轭亚油酸(conjugated linoleic acid,CLA)有20余种,而双键位于碳链9、11和10、12的同分异构体含量最大、生理活性较强[15-16],其中本实验中优势产物亚油酸就属于CLA的一种。CLA能诱导宿主免疫调节实现对炎症性疾病肥胖、糖尿病、呼吸道炎症、肠炎等均有较好治疗效果[17],CLA能改善肥胖糖尿病小鼠的糖脂代谢[18]。2-亚麻酰基-rac-甘油和甘油的相对含量在2种小米中不同可能因为机体内的代谢速率不同,可见古龙贡米的代谢速率快于禾绅小米,可以推测古龙贡米的代谢产物多是因为其代谢速率快于禾绅小米。2-亚麻酰基-rac-甘油为不饱和脂肪酸,是人大麻素1型受体的部分激动剂,可通过甘油三亚油酸酯合成或通过亚麻酰氯合成,能调节其他内源性大麻素包括花生四烯酸甘油酯的活性,花生四烯酸甘油酯能降低促炎因子白介素和肿瘤坏死因子-α,因此推测2-亚麻酰基-rac-甘油可以作为潜在的免疫调节剂,参与神经炎性反应[19-20]。
2.3 2 个品种小米差异代谢产物以及代谢途径分析
2.3.1 古龙贡米差异代谢产物的种类与含量分析
根据对比古龙贡米和禾绅小米二者代谢差异产物可知,古龙贡米差异代谢产物有21种,分别属于氨基酸、糖类、植物甾醇、脂肪酸及其他中间代谢产物。如表2所示,其中棕榈酸乙酯、油酸丙酯两物质相对含量较高,其他产物的相对含量均小于1%。
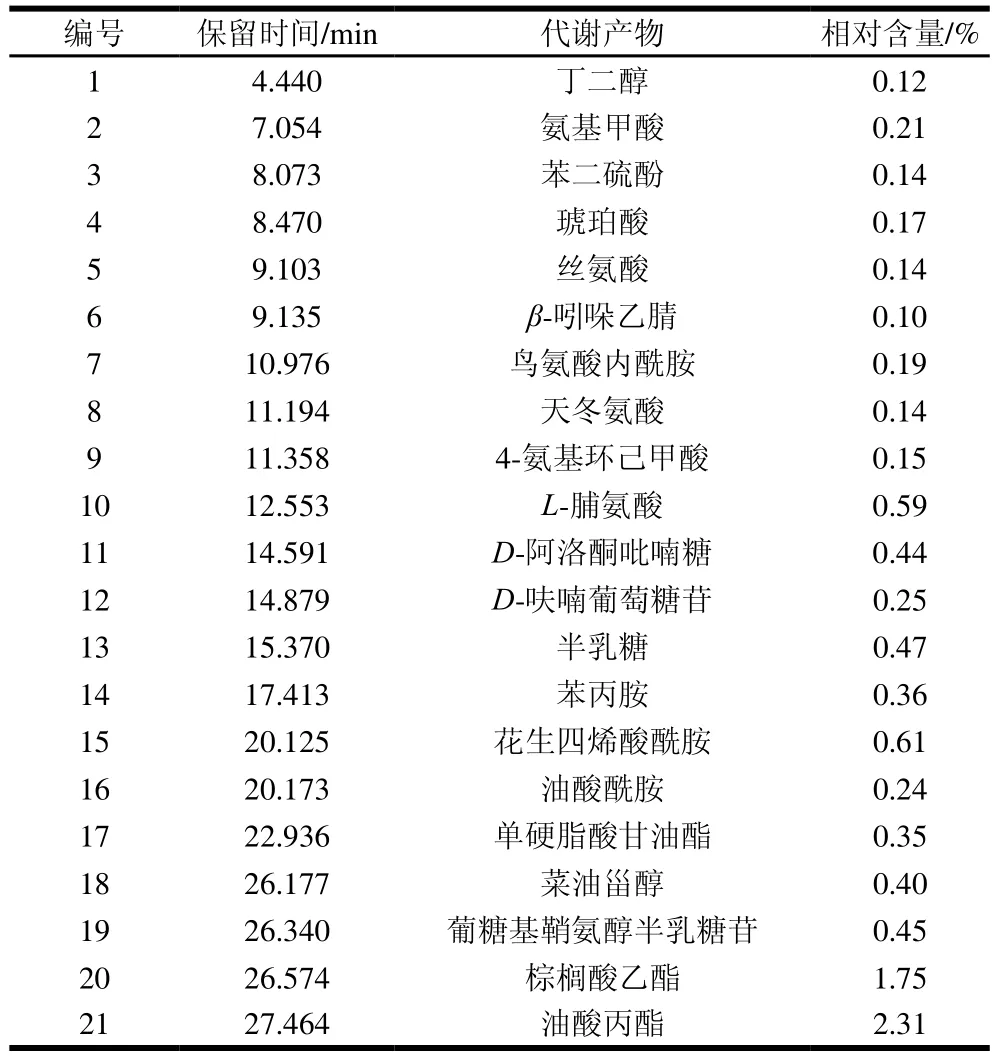
表2 古龙贡米差异代谢产物Table 2 Contents of differential metabolites in ‘Gulong’ millet
2.3.2 古龙贡米差异代谢产物代谢途径
通过KEGG数据库搜索确定2 个品种小米差异代谢产物主要来自于氨基酸代谢、糖代谢、三羧酸循环、植物甾醇代谢、脂肪酸代谢。
2.3.2.1 氨基酸代谢途径分析
丝氨酸可通过丝氨酸消旋酶与D-丝氨酸相互转化,丝氨酸消旋酶的直系同源基因为SRR;可通过甘氨酸羟甲基转移酶和丙氨酸-乙醛酸转氨酶与甘氨酸和羟基丙酮酸相互转化,2种酶的直系同源基因为GLyA、SHMT、AGXT;可由右磷丝氨酸通过磷酸丝氨酸磷酸化酶代谢而成,磷酸丝氨酸磷酸化酶的直系同源基因为serB、PSPH;也可由色氨酸合酶α链代谢成色氨酸,色氨酸合酶α链的直系同源基因为trpA。天冬氨酸可与L-天冬酰胺通过天冬酰胺合成酶相互转化,天冬酰胺合成酶的直系同源基因为asnB、ASNS;可与N-氨基甲酰基-L-天冬氨酸盐通过天冬氨酸氨基甲酰转移酶相互转化,天冬氨酸氨基甲酰转移酶的直系同源基因为CAD;可通过精氨琥珀酸合酶代谢成L-精氨酸琥珀酸,精氨琥珀酸合酶的直系同源基因为argG、ASS1;可通过腺苷琥珀酸合酶代谢成腺苷琥珀酸酯,腺苷琥珀酸合酶的直系同源基因为purA、ADSS;也可通过高丝氨酸脱氢酶1代谢成L-4-天冬氨酰磷酸,高丝氨酸脱氢酶1的直系同源基因为lysC。L-脯氨酸可通过脯氨酰4-羟化酶与羟脯氨酸相互转化,脯氨酰4-羟化酶的直系同源基因为P4HA;可通过吡咯啉-5-羧酸还原酶和脯氨酸脱氢酶与1-吡咯啉-5-羧酸盐相互转化,2种酶的直系同源基因为proC、PRODH、faDM、putB;也可由肽通过脯氨酸亚肽酶代谢而成,脯氨酸亚肽酶的直系同源基因为pip。
2.3.2.2 糖代谢途径分析
D-阿洛酮糖作为D-果糖的差向异构体,可通过微生物来源的酮糖3-差向异构酶催化D-果糖C-3差向异构化获得[21]。葡萄糖苷参与苯并嗪生物合成,由磷酸吲哚甘油通过吲哚-3-甘油磷酸裂解酶代谢而成。半乳聚糖可通过β-半乳糖苷酶代谢出D-半乳糖,或者由乳糖通过β-半乳糖苷酶代谢出D-半乳糖,β-半乳糖苷酶的直系同源基因为lacZ。在葡糖基鞘氨醇半乳糖苷中,葡糖基鞘氨醇则是通过鞘脂类代谢过程中的碎片形成的,在代谢途径中,多条通路可以代谢出鞘氨醇,而葡糖基则可以通过乳糖神经酰胺直接代谢而成,或鞘氨醇间接代谢而成,可能与半乳糖苷连接而成。
2.3.2.3 植物甾醇代谢
菜油甾醇由亚甲基胆固醇通过固醇还原酶代谢而成,菜油甾醇和谷甾醇参与了类固醇生物合成,菜油甾醇通过类固醇22-α-羟化酶代谢出22-α-羟基樟脑醇,具有降胆固醇、促生长、抗氧化等作用[14,22-24]。还有含量较少的物质β-吲哚乙腈参与到吲哚衍生,经查文献可知吲哚乙腈为植物生长素[25],葡糖胺、3-吲哚乙醛肟可通过酶的作用代谢成吲哚乙腈,如图4所示,吲哚乙腈则可通过酶的作用代谢成吲哚-3-乙酰胺和吲哚乙酸,而且色氨酸也可通过吲哚乙腈转变为吲哚乙酸[14]。

图4 葡糖胺(A)和吲哚-3-乙醛肟(B)代谢生成吲哚乙腈Fig.4 Metabolism of glucosamine (A) and indole-3-acetaldehyde oxime (B) into indole acetonitrile
丁二醇可由木糖在微生物[26]的作用下转化形成,丁二醇也可能是在丙酮酸代谢途径中由丙酮酸脱羧脱氢形成乙醇后形成丁二醇,或1-(3-吡啶基)-1,4-丁二醇断键而成。推测鸟氨酸内酰胺可有精氨酸在精氨酸酶的催化作用下生成鸟氨酸再酰基化形成鸟氨酸酰胺。推测花生四烯酸酰胺、油酸酰胺、单硬脂酸甘油酯、棕榈酸乙酯、油酸丙酯等由脂肪的水解或者乙酰CoA经一系列缩合过程中的中间产物。而4-氨基环己甲酸、苯二硫酚、苯丙胺代谢途径未知,其中苯二硫酚是一种重要的医药中间体,可用于抗心绞痛、抗冠心病,也可治疗眼睛红肿、疼痛等[27];而苯丙胺属于精神兴奋剂的一类[28],也可用来治疗帕金森综合征和嗜睡症等[29]。
2.3.3 禾绅小米差异代谢产物的种类与含量分析
对比古龙贡米和禾绅小米二者代谢差异物可知,禾绅小米差异性代谢产物有5种,如表3所示。禾绅小米的差异代谢产物柠檬酸、D-甘露糖、棕榈酸、D-葡萄糖酸、焦性没食子酸相对含量都比较低,均低于1%。

表3 禾绅小米差异代谢产物Table 3 Contents of differential metabolites in ‘Hesen’ millet
2.3.4 禾绅小米差异代谢产物代谢途径
禾绅小米差异代谢产物主要参与了脂肪酸代谢、糖代谢、三羧酸循环。
2.3.4.1 脂肪酸代谢途径分析
棕榈酸可经脂肪水解或乙酰CoA经一系列生物合成,主要是由十六烯酰基-[酰基载体蛋白]通过脂肪酰基ACP硫酯酶A代谢形成,脂肪酰基ACP硫酯酶A的直系同源基因为FATA;也可由棕榈酰蛋白硫酯酶通过棕榈酰辅酶代谢形成,棕榈酰辅酶的直系同源基因为PPT。
2.3.4.2 糖代谢途径分析
D-甘露糖可由D-葡萄糖差向异构化形成,可与D-果糖相互转化,也可由D-半乳糖代谢形成。古洛糖通过鸟苷二磷酸-D-甘露糖-3’,5’-异构酶与D-甘露糖相互转化,也可由D-阿拉伯糖通过碳链增长或经葡糖-6-磷酸转化等途径合成。葡萄糖酸可经葡萄糖和尿苷三磷酸反应形成尿苷二磷酸葡糖,接着被氧化成二磷酸尿苷-葡糖醛酸然后水解生成,葡萄糖酸也可通过葡萄糖激酶代谢成D-核糖-5-磷酸。柠檬酸由乙酰CoA在柠檬酸合酶催化作用下与草酰乙酸缩合形成。
没食子酸是一种广泛存在的酚类化合物,以许多形式存在于绿色植物的组织中,尤其是在茶叶中,均以游离的形式存在或作为聚合物的一部分如单宁、鞣花单宁、茶黄素-3-没食子酸酯、表没食子儿茶素-3-没食子酸酯等[30]。焦性没食子酸可由没食子酸脱羧而成,在数据库中未找到焦性没食子酸的代谢途径,推测在实验过程中可能因为加热由没食子酸断键脱羧而成,焦性没食子酸也多应用于农药、合成医药、化妆品、显影剂、热敏剂、高分子材料的助剂以及化学分析试剂等方面[31]。
3 结 论
通过实验探究黑龙江省肇源县同一产地不同品种小米(古龙贡米、禾绅小米)的代谢产物,共检测出59种代谢产物,其中古龙贡米代谢产物占48种,禾绅小米代谢产物占32种。在二者相同的27种代谢产物中,古龙贡米的优势代谢产物(>15%)为亚油酸,禾绅小米的优势代谢产物为亚油酸。
由于小米品种不同,基因也就不完全相同,所得代谢产物也存在一定的差异性。古龙贡米相对于禾绅小米的代谢差异物有21种,其中含多种必需氨基酸、不饱和脂肪酸及其他活性代谢产物,禾绅小米相对于古龙贡米的差异代谢产物只有5种,在分析2种样品差异代谢产物的代谢途径时,古龙贡米的差异代谢产物主要参与了氨基酸代谢、糖代谢、三羧酸循环、植物甾醇代谢、脂肪酸代谢;而在禾绅小米的差异代谢产物主要参与了脂肪酸代谢、糖代谢、三羧酸循环。在古龙贡米代谢产物中发现了苯二硫酚、苯丙胺;在禾绅小米代谢产物中发现了焦性没食子酸。由此可见,古龙贡米的代谢过程相比禾绅小米更为复杂,古龙贡米的葡萄糖代谢途径、脂肪酸代谢途径、氨基酸代谢途径和三羧酸循环都比禾绅小米对应的代谢途径更活跃,推测古龙贡米的代谢产物多是因为其代谢速率快于禾绅小米。这项研究为充分利用小米中生理功能活性成分提供数据支撑,也为小米优选优良种质资源品质分析、分品种开发功能食品或保健品提供理论基础和依据。
猜你喜欢
山东第一医科大学(山东省医学科学院)学报(2021年7期)2021-10-13
昆明医科大学学报(2020年12期)2021-01-26
河北农业(2020年5期)2020-09-07
人物画报(2020年29期)2020-03-14
人物画报(2020年36期)2020-03-13
农家科技中旬版(2020年1期)2020-03-12
新农村(2018年11期)2018-11-15
文苑(2018年20期)2018-11-09
思维与智慧·下半月(2016年4期)2016-04-25
意林·少年版(2014年13期)2014-08-27
