
抗鼠疫抗原F1的嵌合抗体制备及特性分析
2022-01-05 08:14周东明周婷婷汪茂荣
南京医科大学学报(自然科学版) 2021年12期
匡 衡,蔡 欣,杨 展,周东明,周婷婷*,汪茂荣*,朱 进
1东部战区总医院秦淮医疗区肝病科教研室,安徽医科大学八一临床学院,江苏 南京210001;2东部战区疾病预防控制中心,江苏 南京210002
鼠疫是由鼠疫耶尔森菌引起的一种严重的人畜共患传染病,在过去数千年里造成了约2亿人死亡。有记载的鼠疫大爆发约有3次,分别是查士丁尼瘟疫、黑死病和第3次鼠疫大爆发,均对人类产生了重要影响,其中第3次鼠疫大爆发起源于我国云南并持续至今[1]。
鼠疫耶尔森菌、小肠结肠炎耶尔森菌和假结核耶尔森菌,同属肠杆菌科耶尔森菌属。鼠疫耶尔森菌有极快的繁殖速度和较强的侵袭性,感染者死亡率极高,其侵袭性的强弱与杆菌表面抗原有较大关系。F1蛋白是鼠疫杆菌荚膜抗原,由caf1操纵子编码,鼠疫杆菌感染人体后,F1抗原形成了高分子量的聚合物,聚集在细菌表面[2]。F1抗原可与体内红细胞、淋巴细胞及组织细胞黏附,直接损伤组织,且随血液扩散并大量繁殖[3-4]。
目前F1抗原主要用于制备亚单位疫苗,同时也可用于鼠疫鉴定。但鼠源性抗体极易被人体免疫系统所排斥,同时鼠源Fc片段难以激活人体免疫系统。人源抗体的Fc片段,可以与人体内血管内皮细胞上的FcRn受体特异性结合,从而避免在体内被迅速降解[5],鼠源性抗体因为不能与其结合,在人体内半衰期较短,限制了鼠源抗体的临床应用。兰州生物制品研究所有限责任公司甘肃省疫苗工程技术研究中心实验室前期制备并获得了稳定表达抗鼠疫F1抗体的杂交瘤细胞株4C6,并完成了抗体的保护性试验[6]。本研究拟在前期研究的基础上,制备抗鼠疫保护性抗原F1嵌合抗体,以期在临床检验和鼠疫治疗中发挥作用。
1 材料和方法
1.1 材料
F1-4C6杂交瘤细胞及抗F1抗体由兰州生物制品研究所有限责任公司甘肃省疫苗工程技术研究中心实验室前期制备,TOP10感受态细胞、胶回收试剂盒(上海生工生物工程股份有限公司),ProteinG纯化柱(GE公司,美国),CHOMaxA培养基(上海迈邦生物有限公司),HRP标记山羊抗人IgG二抗、Millipore超滤离心管(10 000 kDa)(Sigma公司,美国),限制性内切酶NotⅠ和EcoRⅠ(NEB公司,美国),TMB显色液及终止液(上海碧云天),ProteinG琼脂糖珠、总RNA提取试剂盒(Thermo公司,美国)。
1.2 方法
1.2.1 人源化质粒构建
将生长良好的F1-4C6细胞消化裂解后提取细胞总RNA,逆转录得到cDNA,以cDNA为模板分别扩增F1抗体轻、重链可变区基因,切胶回收后连接于pMD-18t载体上,并转化入TOP10感受态细胞中,涂板培养挑取单克隆后测序,通过比对和分析,获得抗F1抗体轻、重链可变区基因序列。将抗体的可变区序列人源抗体的恒定区基因序列拼接,序列优化后全基因合成并克隆进表达质粒pMH3中,分别命名为pMH3-f1-k和pMH3-f1-h。
1.2.2 稳定表达真核细胞系建立与筛选
取5×106个生长良好的CHO-S细胞与pMH3-f1-k和pMH3-f1-h两种质粒混匀后加入0.2 mm电转杯中进行电转染,转染后的细胞在含10%血清的DMEM/F12培养基中培养,48 h后取培养液进行斑点分子杂交法(Dot blot)检测其表达。取100 μL上清,加入含50 ng F1抗原进行ELISA检测,37℃孵育1 h,用PBST溶液清洗3遍,随后用辣根过氧化物酶(horseradish peroxidase,HRP)标记的抗人Fc段抗体孵育1 h,磷酸盐吐温缓冲液(phosphate buffered saline tween,PBST)溶液清洗5遍后,四甲基联苯胺(tetramethylbenzidine,TMB)显色液显色5 min,观察显色情况。
使用终浓度为1 000 μg/mL G418培养液培养阳性细胞,2周后挑取单克隆于96孔板中培养,1周后做Dot blot检测,分别从6个阳性孔中挑取110个细胞加入96孔板中培养,1周后挑取单克隆并做Dot blot检测,挑选表达量最高的6株细胞进行扩增。
将6株细胞稳定培养10代并冻存细胞,观察生长情况并取上清进行ELISA检测,并测定每株细胞450 nm吸光度值,挑选一株生长迅速且蛋白表达量高的细胞进行大量培养。
1.2.3 抗体表达与纯化
对大量培养的细胞进行计数,取3×107个细胞加入30 mL CHOmaxA培养基中,在37℃条件下悬浮培养,每隔2 d检测细胞密度,观察细胞生长状况,15 d后离心收集上清。将收集的上清经0.22 μm的滤膜过滤后,使用Protein G纯化柱纯化。浓缩后用磷酸盐缓冲液(phosphate buffered saline,PBS)溶液调整其浓度为1 mg/mL。取20 μL抗体加入带有还原剂的加样缓冲液,于100℃水浴锅中水浴5 min,随后进行十二烷基硫酸钠聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate polyacrylamide gel electrophoresis,SDS-PAGE),并进行考马斯亮蓝染色。
1.2.4 抗体特性分析
1.2.4.1 间接ELISA检测
每孔50 ng F1抗原包被酶标板,将纯化抗体从1∶200开始梯度倍比稀释后加入酶标板,37℃孵育1 h后洗涤3遍,加入HRP标记二抗,继续于37℃孵育1 h后洗涤3遍,用TMB显色液显色,测定D(450 nm)值。
1.2.4.2 竞争性ELISA检测
50 ng/孔F1抗原包被酶标板,以1∶2 000浓度加入抗F1抗体孵育1 h,洗涤后,将纯化后的人源化F1嵌合抗体从1∶50开始倍比稀释,加入酶标板孵育1 h,PBST洗涤5遍,加入HRP标记的抗人Fc段抗体孵育后显色,测定D(450 nm)值。
1.2.4.3 亲和力检测
使用Biacore X100分析仪进行亲和力检测。选取CM5芯片,并在芯片上固定F1抗体,将抗F1的嵌合抗体和抗F1鼠源抗体梯度稀释后进样分析。使用Biacore X100软件分别计算其亲和力。
1.2.4.4 免疫沉淀
将抗F1抗原嵌合抗体与含有裂解液的F1抗原混合,于4℃下摇床孵育过夜,加入Protein G琼脂糖于4℃下摇晃过夜。离心后取沉淀进行SDS-PAGE凝胶电泳,考马斯染色后切割条带做质谱分析。
2 结果
2.1 抗F1抗体轻链、重链真核表达载体的构建
PCR获得4C6杂交瘤细胞中抗F1轻、重链可变区基因,琼脂糖凝胶电泳检测条带大小符合预期,轻链为300 bp左右,重链为350 bp左右(图1),分别将轻、重链可变区基因片段克隆到pMD-18T载体中,并转化至Top10感受态细胞,培养并挑取单克隆测序。分别将抗F1抗体轻链、重链可变区基因与人源抗体恒定区基因拼接后,全基因合成并克隆进pMH3质粒中,质粒双酶切结果显示轻链条带为700 bp左右,重链条带为1 400 bp左右,与设计条带大小相符(图2)。
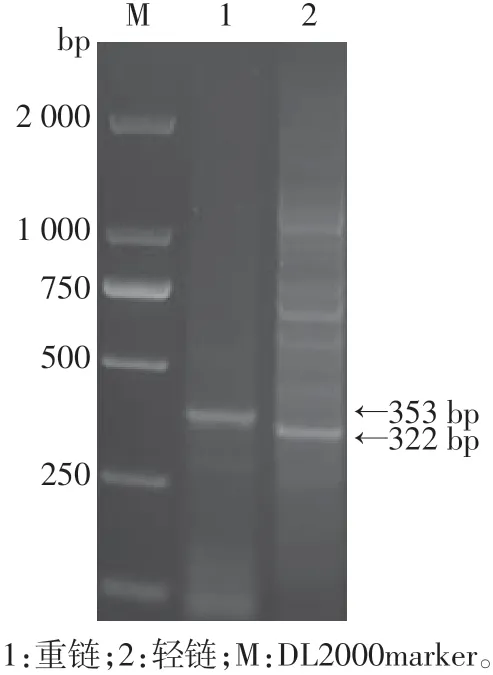
图1 PCR获得轻重链条带Figure 1 Light and heavy chain obtained by PCR

图2 轻重链质粒双酶切条带Figure 2 Light and heavy chain plasmid by double enzyme digestion
2.2 稳定表达细胞系建立与筛选
使用电穿孔技术,将轻、重链表达质粒共转入CHO-S细胞中,转染48 h后,取上清进行Dot blot与ELISA检测。结果表明,转染后的细胞可以表达抗F1嵌合抗体IgG,并与酶标板包被的F1抗原结合(图3)。

图3 转染后培养液Dot blot检测Figure 3 Dot blot detection of culture medium after transfection
使用G418筛选2周,将细胞挑取2次单克隆,取上清做Dot blot检测,挑选目的蛋白表达量最高的6株单克隆细胞,稳定传代培养10代,取上清进行ELISA检测,挑选表达蛋白量最高的一株命名为CHMIgG-F1C6-5,扩大培养进行下一步实验(图4)。

图4 6株细胞培养10代后培养液ELISA检测Figure 4 Culture medium after 10 generations of 6 cell lines detected by ELISA
2.3 抗体表达与纯化
将3×107个细胞悬浮培养于30 mL CHOmaxA培养基中,每2 d测定细胞密度。细胞培养15 d后,离心并收集上清进行Dot blot与ELISA检测均为阳性,结果提示,CHMIgG-F14C6-5细胞可以于悬浮培养基中稳定表达抗F1嵌合抗体。
上清经ProteinG亲和纯化后,得到抗F1嵌合抗体,加入PBS溶液调整浓度为1 mg/mL。经SDSPAGE凝胶电泳检测,显示抗体轻、重链大小符合预期,且浓度较高(图5)。
2.4 抗体分析
2.4.1 间接ELISA检测
将抗体从1∶200稀释度开始稀释,加入含F1抗原的酶标板中进行ELISA检测。结果显示,抗体稀释度在1∶400以上时,D(450 nm)值与抗体浓度呈正相关,结果提示,嵌合抗体与F1抗原特异性结合,D(450 nm)值与抗体浓度呈正相关(图6)。

图6 抗体间接ELISA检测Figure 6 Indirect ELISA detection of antibodies
2.4.2 竞争性ELISA
将抗F1鼠源抗体加入带有F1抗原的酶标板中,随后加入人源化的嵌合抗体与其竞争结合F1抗原。当嵌合抗体稀释度为1∶50时,D(450 nm)值接近间接ELISA实验中嵌合抗体饱和时D(450 nm)值(图7)。该结果提示,当嵌合抗体稀释度为1∶50时,酶标板上F1结合位点大部分结合嵌合抗体。随着嵌合抗体的浓度逐渐降低,D(450 nm)值也随之不断减小。该实验证明,抗F1嵌合抗体能够与鼠源抗体竞争结合F1抗原上的相同表位。

图7 嵌合抗体与鼠源抗体竞争性ELISA检测Figure 7 Competitive ELISA detection of human chimeric antibody and mouse antibody
2.4.3 亲和力检测
两种抗体进样后分析其亲和力,嵌合抗体及鼠源抗体的平衡解离常数KD值分别为2.303×10-9mol/L与1.830×10-9mol/L,两种抗体与F1抗原均有较高的亲和力(图8、9)。

图8 Biacore T100检测抗F1嵌合抗体亲和力Figure 8 Biacore T100 assay results of human F1 antibody affinity
2.4.4 免疫共沉淀与质谱分析
免疫共沉淀后的样本,经SDS-PAGE电泳和考马斯亮蓝染色后,于15 kDa左右发现目的条带(图10),其大小与F1抗原一致,切胶回收后进行质谱分析,该蛋白为鼠疫F1抗原(图11)。

图10 嵌合抗体IP检测Figure 10 Immunoprecipitation analysis of chimeric antibody
图11 嵌合抗体质谱分析Figure 11 Mass spectrometry analysis of chimeric antibodies
3 讨论

图9 Biacore T100检测鼠源抗F1抗体亲和力Figure 9 Biacore T100 assay results of mouse F1 antibody affinity
人源化抗体常分为嵌合抗体、表面重塑抗体及改型抗体3类,其中嵌合抗体是利用DNA重组技术,将异源单抗的轻、重链可变区基因插入含有人抗体恒定区的表达载体中,转染哺乳动物细胞表达出嵌合抗体[7-8]。表面重塑抗体改变的氨基酸不多,人源化不彻底,在应用上具有安全隐患,而改型抗体改变了抗体FR区基因,会降低抗体亲和度,影响抗体效果[9]。
本研究采用制备嵌合抗体的技术方法,成功获得了抗F1嵌合抗体,该抗体与F1抗原亲和力较强,但嵌合抗体仍保留较长鼠源基因序列,导致抗体在人体内疗效不确定,安全性和实用性有待观察。
CHO-S是一种常于生物制药的悬浮细胞系,该细胞系具有生长迅速,转染效率高,极少分泌内源蛋白,产物蛋白分泌于胞外,又免去了细胞破碎这一步骤,可以有效减少杂质蛋白对抗体的污染,是目前实验室中最适合生产单克隆抗体的细胞系之一[10]。本研究使用化学成分确定的3种培养基CHO MaxA、CHOMaxFA、CHO MaxFB悬浮培养CHO细胞,此3种培养基不含杂质蛋白,有效抑制了Protein G柱纯化过程中杂质蛋白的产生[11],从而实现抗体高表达,同时也利于精简抗体纯化步骤,缩减抗体生产成本,有利于大量生产。而同样被广泛应用于抗体生产的大肠杆菌系统,由于其极易产生内毒素和包涵体的原因,未被本课题采用。
鼠疫的临床治疗通常以链霉素联用其他类型抗生素为主。在预防鼠疫方面,灭活疫苗已投入临床应用近百年,尽管近年来已研制出亚单位疫苗及DNA疫苗,但有待临床试验验证其安全性及有效性。近年来发现多种耐药鼠疫杆菌,单纯的药物治疗效果不明显[12-14],随着恐怖主义势力抬头,耐药鼠疫杆菌被应用于生化武器研发,高效鼠疫治疗方案的开发被提上日程。本研究制备的抗F1嵌合抗体可以与F1抗原特异性结合,可有效应用于鼠疫检测与治疗。在后续研究中,需进行动物实验及临床试验,分析其安全性及保护效率,为人源化中和抗体应用于鼠疫临床救治奠定基础。
猜你喜欢
生殖医学杂志(2022年10期)2022-10-19
交通医学(2022年2期)2022-06-14
昆明医科大学学报(2021年1期)2021-02-07
学生天地(2020年15期)2020-08-25
发明与创新·大科技(2020年1期)2020-04-17
中小学德育(2020年11期)2020-03-18
新民周刊(2019年47期)2019-12-26
生物学教学(2018年10期)2018-11-29
教育界·上旬(2016年12期)2017-05-25
标记免疫分析与临床(2016年9期)2016-11-21
