
黄连素通过激活AMPK改善同型半胱氨酸诱导的大鼠痴呆和Tau蛋白过度磷酸化*
2021-12-30 05:22胡泽波周新文
中国病理生理杂志 2021年12期
王 林, 丁 见, 吴 超, 张 翠, 李 曙, 胡泽波, 周新文
(1皖南医学院基础医学院,安徽芜湖241002;2华中科技大学病理生理学系,湖北武汉430030)
阿尔茨海默病(Alzheimer disease,AD)是一种不可逆、且进行性的神经退行性疾病,约占所有老年痴呆症病例的 60%~80%[1]。AD 两大主要特征性病理改变为β 淀粉样多肽(amyloid beta peptide,Aβ)沉积形成的老年斑和神经原纤维缠结(neurofibrillary tangles,NFTs),NFTs 主要由过度磷酸化的 Tau 蛋白形成[2]。Tau 蛋白可以在多个丝氨酸(serine,Ser)或苏氨酸(threonine,Thr)位点被磷酸化,例如 Ser396(pS396)和 Thr205(pT205)等。高同型半胱氨酸血症、高血糖、神经炎症、氧化应激和高胆固醇血症等都与AD 发生发展密切相关[3],我们前期建立的尾静脉注射同型半胱氨酸(homocysteine,Hcy)大鼠AD样模型显示,模型大鼠的认知功能受损,海马区Tau蛋白过度磷酸化[4]。AD 的病因和发病机制复杂,药物开发至今为止多以失败告终。
中药黄连素(berberine,BBR)是一种广泛应用的异喹啉生物碱,具有抗病毒、抗菌和抗炎等多种生物活性[5]。研究表明黄连素对多种中枢神经系统疾病,如帕金森病、亨廷顿病和AD 等具有神经保护作用[6-8]。在糖尿病大鼠中,黄连素可抑制炎症、氧化应激和改善脑中胰岛素抵抗进而改善大鼠的认知功能[9];在 APP/Tau/PS1 AD 转基因鼠中给予黄连素治疗后,可降低Aβ1-42的水平和改善认知障碍[10]。以上研究结果表明黄连素在AD 发病机制的多个环节起作用,但是黄连素如何调控Tau 蛋白磷酸化水平的具体机制尚不明确。
腺苷酸活化蛋白激酶(AMP-activated protein kinase,AMPK)是一种丝氨酸/苏氨酸蛋白激酶,由一个催化亚基α、两个调节亚基β 和γ 组成的异源三聚体复合物,上游可通过磷酸化AMPK α 亚基上Thr172 位点(p-AMPK)激活AMPK[11]。AMPK 可调节细胞内的一些代谢途径,如葡萄糖的摄取和运输,脂肪酸和蛋白质的合成,以及脂肪酸的β-氧化等。本课题组前期研究已证实,随着年龄增加,小鼠海马中AMPK 的活性逐渐降低,而激活AMPK 可降低Tau 蛋白的磷酸化水平[12]。在细胞水平上黄连素可激活AMPK[13],在 AD 动物模型中,黄连素能否通过激活AMPK 改善认知功能障碍和降低Tau 蛋白磷酸化水平值得研究。因此,本研究利用尾静脉注射Hcy 建立AD 样大鼠模型,探究黄连素的神经保护作用及机制。
材料和方法
1 实验动物及处理方法
36 只 SPF 级 3 月龄 SD 大鼠,雄性,体重 200~250 g,购自长沙市天勤生物技术有限公司,许可证号为SCXK(湘)2019-0014。实验前适应性饲养1 周,自由饮食、饮水,温度(23±1)℃,湿度(50±10)%,光照和黑暗交替各12 h。36 只大鼠随机分为3 组:对照组(NS 组)、模型组(Hcy+NS 组)及治疗组(Hcy+BBR组),每组12 只。模型组和治疗组大鼠尾静脉注射Hcy(400 μg·kg-1·d-1)14 d,对照组大鼠尾静脉注射相同剂量的生理盐水(normal saline,NS)14 d。随后治疗组大鼠用黄连素灌胃(100 mg·kg-1·d-1)10 周,对照组和模型组大鼠用相同体积的NS 灌胃10 周。具体流程图见图1A。所有动物实验均经过皖南医学院实验动物福利与伦理委员会批准。
2 细胞培养
HEK293-Tau 细胞(稳定转染人类全长Tau 蛋白的人胚肾母细胞瘤细胞)由华中科技大学同济医学院病理生理学系周新文老师课题组惠赠,细胞于5%CO2、37 ℃恒温恒湿的细胞培养箱中,用含10%胎牛血清和G418 的DMEM 高糖培养液培养。先检测不同浓度(2.5、5、10 和 20 μmol/L)BBR 对细胞活力和AMPK 活性的影响,最终确定用于后续实验的浓度为 5 μmol/L。细胞实验分组为 3 组:对照(control,CON)组:单独 DMSO 处理 24 h;BBR 组:5 μmol/L BBR 和 DMSO 共同处理 24 h;BBR+Compound C 组:5 μmol/L BBR 和 20 μmol/L Compound C 共 同 处 理24 h。
3 主要药品试剂和仪器
Hcy 和 Compound C 购自 Sigma;BBR 购自上海源叶生物科技有限公司;兔抗AMPK、p-AMPK、PSD93、PSD95、糖原合成酶激酶3β(glycogen synthase kinase-3,GSK-3β)和GSK-3β Ser9 位点磷酸化(GSK3β-S9)多克隆抗体购自Cell Signaling Technology;兔抗pT205 和pS396 多克隆抗体购自Biosource;鼠抗蛋白磷酸酶2A 催化亚基(protein phosphatase 2A catalytic subunit,PP2Ac)和 PP2Ac 亮氨酸 309 位点去甲基化(DM-PP2Ac)单克隆抗体购自 Sigma;鼠抗 PP2Ac 的酪氨酸307 位点磷酸化(p-PP2Ac)单克隆抗体购自Santa Cruz Biotechnology;鼠抗 β-actin 和 Tau5 单克隆抗体购自Abcam;CCK-8 试剂盒、羊抗兔IgG 和羊抗鼠IgG购自碧云天生物技术有限公司;高尔基染色试剂盒购自FD Neuro Technologies;胎牛血清、DMEM高糖培养基、G418 和胰酶购自Gibco。培养瓶和培养板等细胞培养耗材购自NEST;SDS-PAGE 电泳仪及成像分析仪购自Bio-Rad;倒置显微镜购自Olympus;水迷宫实验设备购自泰盟公司。
4 主要实验方法
4.1 Morris 水迷宫实验 用Morris 水迷宫实验评价大鼠的空间学习记忆能力。前5 d 训练大鼠在水迷宫设备中寻找一个隐藏的平台(低于水面1 cm),每天训练4 次,在每次实验中,当大鼠爬上平台并保持15 s 时,实验结束;如果大鼠在90 s 内没有找到隐藏的平台,它们会被引导到平台上,并停留15 s。记录大鼠找到平台的时间(逃避潜伏期)。在检测记忆能力之前,让大鼠休息1 d,第7天移走平台进行空间探索实验,大鼠自由游泳90 s,记录大鼠游过原平台的次数、大鼠在原平台所在象限的时间及大鼠游泳的速度。采用装置自带跟踪软件记录大鼠的运动轨迹。实验期间保持固定的实验人员和安静的实验环境。
4.2 Western blot 于Morris 水迷宫实验后收集大鼠脑组织,BBR处理24 h后收集细胞样品,BCA 法检测各样品蛋白浓度,随后加入上样缓冲液,沸水中煮10 min使蛋白充分变性。变性后的蛋白(20 μg)进行SDS-PAGE 分离蛋白,将凝胶中的蛋白转移至NC 膜上,并用脱脂奶粉溶液进行封闭,封闭完成后用Ⅰ抗孵育,4 ℃过夜,再加入Ⅱ抗室温孵育2 h,最后在凝胶成像仪上曝光显影。显影后以β-actin 为内参照,用ImageJ软件量化目的蛋白的表达水平。
4.3 免疫组化实验 4%多聚甲醛灌流后快速取脑组织,继续浸泡4%多聚甲醛溶液48 h,随后用30%蔗糖溶液脱水,冰冻切片(片厚30 μm),将脑片放入4℃冰箱保存。挑取脑片PBS 漂洗后加入3%H2O2溶液消除内源性过氧化物酶活性,加入枸橼酸盐缓冲液进行抗原修复,5% BSA 室温封闭60 min,加入Ⅰ抗pS396(稀释比为1:100),4℃孵育过夜。回收Ⅰ抗,PBS 漂洗后加入生物素标记的组化Ⅱ抗(羊抗兔IgG),37℃孵育1 h。最后加入DAB 显色液,避光显色3 min 左右用PBS 冲洗终止显色。贴片,室温干燥后,梯度乙醇脱水、二甲苯透明和中性树胶封片,显微镜下观察和拍照。
4.4 高尔基染色 生理盐水灌流后立即取脑组织,浸泡于试剂盒中A、B 混合液中,24 h 后更换新鲜A、B混合液,室温,避光放置21 d,隔2 d更换AB混合液一次。21 d 后脑组织放入C 液中,24 h 后更换新鲜C液,继续避光放置2 d。震荡切片机(50 μm)切片,脑片室温避光晾干。脑片用双蒸水冲洗后放入D、E 混合液中染色10 min,染色过程中始终保持脑片湿润。梯度乙醇脱水、二甲苯透明和中性树胶封片。低倍镜下先确定海马CA3 区神经元,在油镜下选择距胞体90~110 μm(近端)和190~210 μm(远端)的2 个树突段进行拍照,用ImageJ 计算每个神经元10 μm 树突长度上的树突棘数量,即树突棘密度。
4.5 CCK-8 法检测BBR 对细胞活力的影响 将制备好的细胞悬液接种到96 孔板中,每孔约100 μL,将96孔板放入培养箱中培养24 h。各孔中加入不同浓度的黄连素,同时设置空白对照组和正常对照组,每组样本做6 个复孔,继续孵育24 h 后更换含10%CCK-8 的新鲜培养基,加样的过程中尽量不要产生气泡。最后再放入培养箱中孵育30 min,用酶标仪测定450 nm波长处吸光度(A)值并计算细胞活力。
5 统计学处理
使用SPSS17.0 统计软件处理数据,数据用均数±标准差(mean±SD)表示,多组间比较采用单因素方差分析(one-way ANOVA),组间两两比较采用最小显著性差异法(LSD 法),以P<0.05 为差异有统计学意义。
结 果
1 黄连素对Hcy大鼠空间学习记忆能力的影响
Morris 水迷宫结果显示,在训练第5 天,与NS 组相比,Hcy+NS 组大鼠的逃避潜伏期显著延长(P<0.05);与 Hcy+NS 组相比,Hcy+BBR 组大鼠逃避潜伏期显著缩短(P<0.05),见图 1B;与 NS 组相比,Hcy+NS 组大鼠穿越原平台次数显著减少(P<0.05),在原平台所在象限停留时间也显著缩短(P<0.05);与Hcy+NS 组相比,Hcy+BBR 组大鼠穿越原平台次数显著增加(P<0.05),并且在原平台所在象限停留时间也显著延长(P<0.05),见图1D、E;同时记录大鼠的游泳速度,结果显示3 组大鼠游泳速度并无显著差异(P>0.05),见图1C;各组大鼠运动轨迹见图1F。
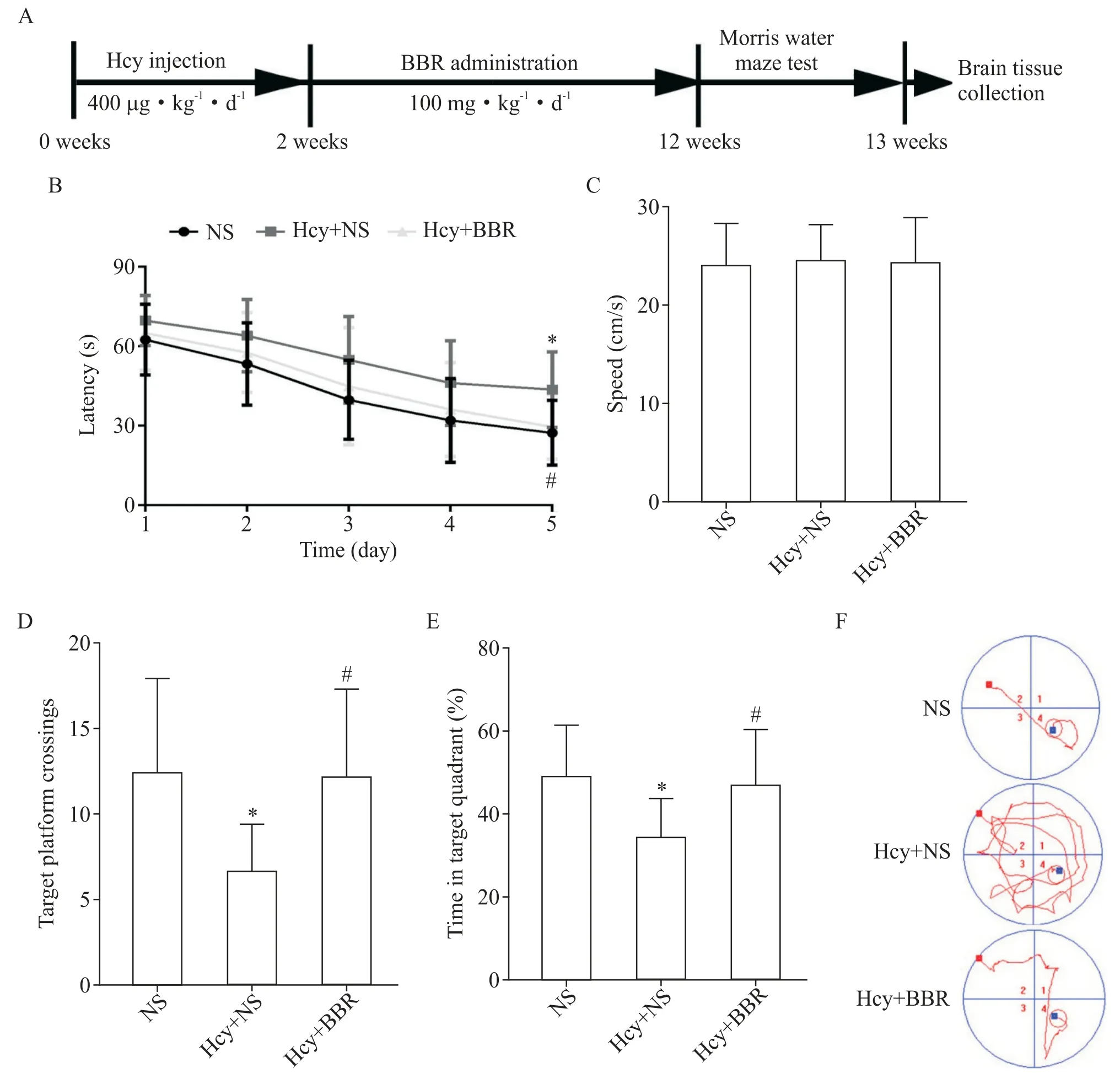
Figure 1. The therapeutic effects of BBR on the cognitive decline of Hcy rats. A:diagram of the experimental procedure;B:the escape latency to find the hidden platform;C:the swim speed on day 7;D:the number of crossing the platform region;E:the time spent in the target quadrant;F:the representative swimming paths on day 5. Mean±SD. n=12.*P<0.05 vs NS group;#P<0.05 vs Hcy+NS group.图1 黄连素对Hcy大鼠认知功能的影响
2 黄连素对Hcy 大鼠海马区突触蛋白和树突棘密度的影响
Western blot 实验结果显示,与 NS 组相比,Hcy+NS 组大鼠海马区突触蛋白(PSD93 和PSD95)的表达水平显著降低(P<0.01);与Hcy+NS 组相比,Hcy+BBR 组大鼠海马中PSD93 和PSD95 的表达水平显著升高(P<0.05),见图2A。高尔基染色结果显示,与NS 组相比,Hcy+NS 组大鼠海马CA3 区树突棘密度显著降低(P<0.01);与Hcy+NS 组相比,Hcy+BBR 组大鼠海马CA3 区树突棘密度显著升高(P<0.05),见图2B。

Figure 2. The expression of synaptic protein and spine density in the hippocampal of the rats in each group. A:comparison of PSD93 and PSD95 protein levels in the hippocampal of rats in each group(n=6);B:representative shaft dendrites in hippocampal neurons of the CA3 region in each group by Golgi staining and quantitative analysis(n=3). Mean±SD.**P<0.01 vs NS group;#P<0.05 vs Hcy+NS group.图2 各组大鼠海马区突触蛋白的表达和树突棘密度
3 黄连素对Hcy 大鼠海马区Tau 蛋白磷酸化水平的影响
Western blot 实验结果显示,与NS 组相比,在Hcy+NS 组大鼠的海马区pT205 表达水平显著升高(P<0.01);与Hcy+NS 组相比,Hcy+BBR 组大鼠海马中 pT205 表达水平显著降低(P<0.01),见图 3A。Western blot 和免疫组化实验结果均显示,与NS组相比,Hcy+NS 组大鼠海马区pS396 表达水平显著升高(P<0.05);与Hcy+NS 组相比,Hcy+BBR 组的大鼠海马区pS396 表达水平显著降低(P<0.05),见图3B。但是总Tau 蛋白(Tau5)的表达水平在各组大鼠海马中无显著差异(P>0.05),见图3。
4 黄连素对Hcy 大鼠海马区Tau 蛋白激酶和磷酸酶的影响
Western blot 实验结果显示,与 NS 组相比,Hcy+NS 组大鼠海马区p-AMPK 表达水平显著降低(P<0.01);与 Hcy+NS 组相比,Hcy+BBR 组的大鼠海马区p-AMPK表达水平显著升高(P<0.01);但是AMPK的表达水平在各组大鼠海马中并无显著差异(P>0.05)。GSK3β-S9和GSK3β 的表达水平在各组大鼠海马中并无显著差异(P>0.05)。与NS组相比,Hcy+NS 组大鼠海马区p-PP2Ac 和DM-PP2Ac 表达水平显著升高(P<0.05);与Hcy+NS 组相比,Hcy+BBR 组的大鼠海马中p-PP2Ac 和DM-PP2Ac 的表达水平显著降低(P<0.05);但是在各组大鼠海马中,PP2Ac 的表达水平无显著差异(P>0.05),见图4。

Figure 3. The expression level of phosphorylated Tau protein in the hippocampal of the rats in each group. A:the levels of total Tau(Tau5)and phosphorylated Tau(pT205,pS396)in the hippocampal of rats in each group were measured by Western blot;B:representative images of brain slice immunostained with pS396 and quantitative analysis of pS396 staining. Mean±SD.n=6.*P<0.05,**P<0.01 vs NS group;#P<0.05,##P<0.01 vs Hcy+NS group.图3 各组大鼠海马区Tau蛋白磷酸化水平
5 不同浓度的黄连素对HEK293-Tau 细胞活力和AMPK活性的影响
在HEK293-Tau细胞中,给予不同浓度BBR作用24 h,CCK-8法检测结果显示,与对照组相比,BBR 分别在 10 μmol/L 和 20 μmol/L 时对细胞的活力抑制有统计学差异(P<0.05),见图5A。Western blot 实验结果显示,BBR(2.5 μmol/L)组p-AMPK 表达水平与对照组相比无显著性差异,而BBR(5 μmol/L)组中p-AMPK 表达水平较对照组显著升高(P<0.01),见图5B。AMPK 的表达水平在各组中并无显著差异(P>0.05),见图5B。
6 黄连素通过AMPK/PP2Ac降低HEK293-Tau细胞中Tau蛋白磷酸化水平
实验结果显示,与 CON 组相比,BBR 组 pT205 和pS396 表达水平显著降低(P<0.01),与BBR 组相比,BBR+Compound C 组pT205和pS396表达水平显著升高(P<0.05),Tau5 的表达水平在各组间无显著差异(P>0.05)。与CON 组相比,BBR 组中 p-AMPK 表达水平显著升高(P<0.05),与BBR 组相比,BBR+Compound C 组中p-AMPK 表达水平显著降低(P<0.05),AMPK 的表达水平在各组间无显著差异(P>0.05)。与 CON 组相比,BBR 组 p-PP2Ac 和 DM-PP2Ac 表达水平显著降低(P<0.05),与BBR 组相比,BBR+Compound C 组 p-PP2Ac 和 DM-PP2Ac 表达水平显著升高(P<0.05),PP2Ac 的表达水平在各组间无显著差异(P>0.05),见图6。
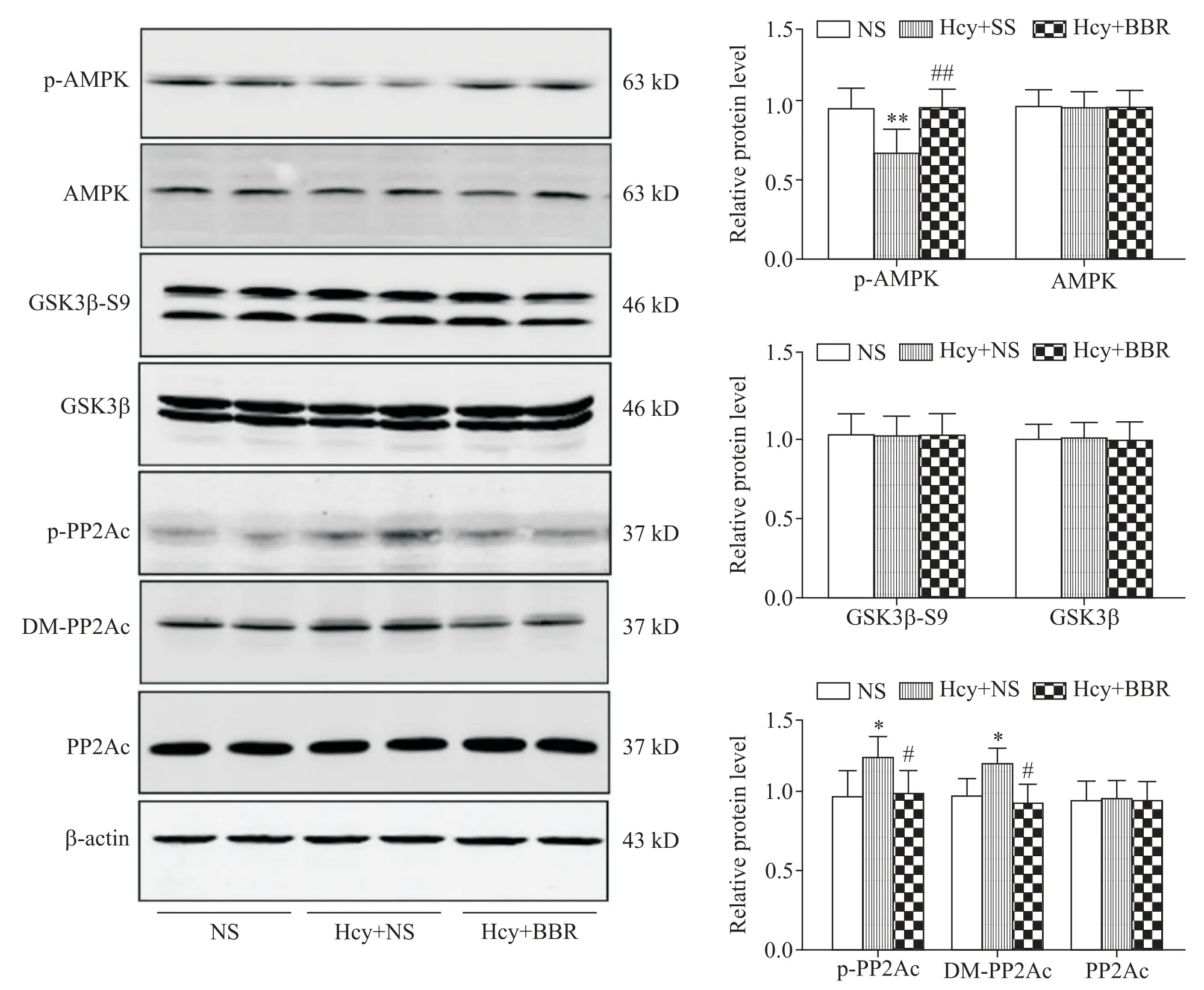
Figure 4. Comparison the activity of AMPK,GSK3β and PP2Ac in the hippocampal of the rats in each group. Mean±SD. n=6.*P<0.05,**P<0.01 vs NS group;#P<0.05,##P<0.01 vs Hcy+NS group.图4 比较各组大鼠海马区AMPK、GSK3β和PP2Ac的活性
讨 论
研究报道,增加大鼠血浆中Hcy 浓度,可以诱导AD 样认知功能障碍、Aβ 沉积和Tau 蛋白过度磷酸化[14-16]。本研究与以往研究结果相似,水迷宫实验结果显示Hcy 大鼠的学习记忆能力受损。Hcy 通过多种途径分解代谢,其中之一是叶酸/维生素B12 依赖的再甲基化。虽然维生素B12 和叶酸的补充可有效的降低血浆中Hcy 浓度,但与安慰剂相比,叶酸和维生素B12 的补充并没有改善健康或认知障碍患者的认知功能[17]。这一结果表明Hcy 可能通过血管或神经毒性等多种途径引起脑功能障碍。
黄连素可改善AD 转基因鼠(3×Tg AD 小鼠)和2型糖尿病大鼠的认知功能障碍[18-19],但黄连素能否改善Hcy 诱导的大鼠认知功能障碍目前并没有被详细探讨。本研究水迷宫实验结果显示黄连素(100 mg·kg-1·d-1)灌胃 10 周后,可显著改善 Hcy 大鼠的空间学习记忆能力。突触是学习和记忆的基本单位,在轻度认知损伤患者和AD 患者的早期阶段已观察到突触功能受损[20];在AD 转基因鼠(APP/PS1 小鼠)海马中,突触蛋白PSD93 和PSD95 的含量显著降低,提高突触蛋白的表达水平可改善认知功能[21]。Hcy 可引起 PSD93 和 PSD95 的表达水平降低[15]。为解释黄连素显著改善Hcy 大鼠的空间学习记忆能力的机制,我们检测了树突棘密度和突触蛋白的表达水平,结果显示黄连素治疗后Hcy 大鼠海马区PSD93 和PSD95 表达水平显著升高,树突棘密度显著增加。表明黄连素改善Hcy 大鼠的认知功能可能与增加突触蛋白和树突棘密度有关。
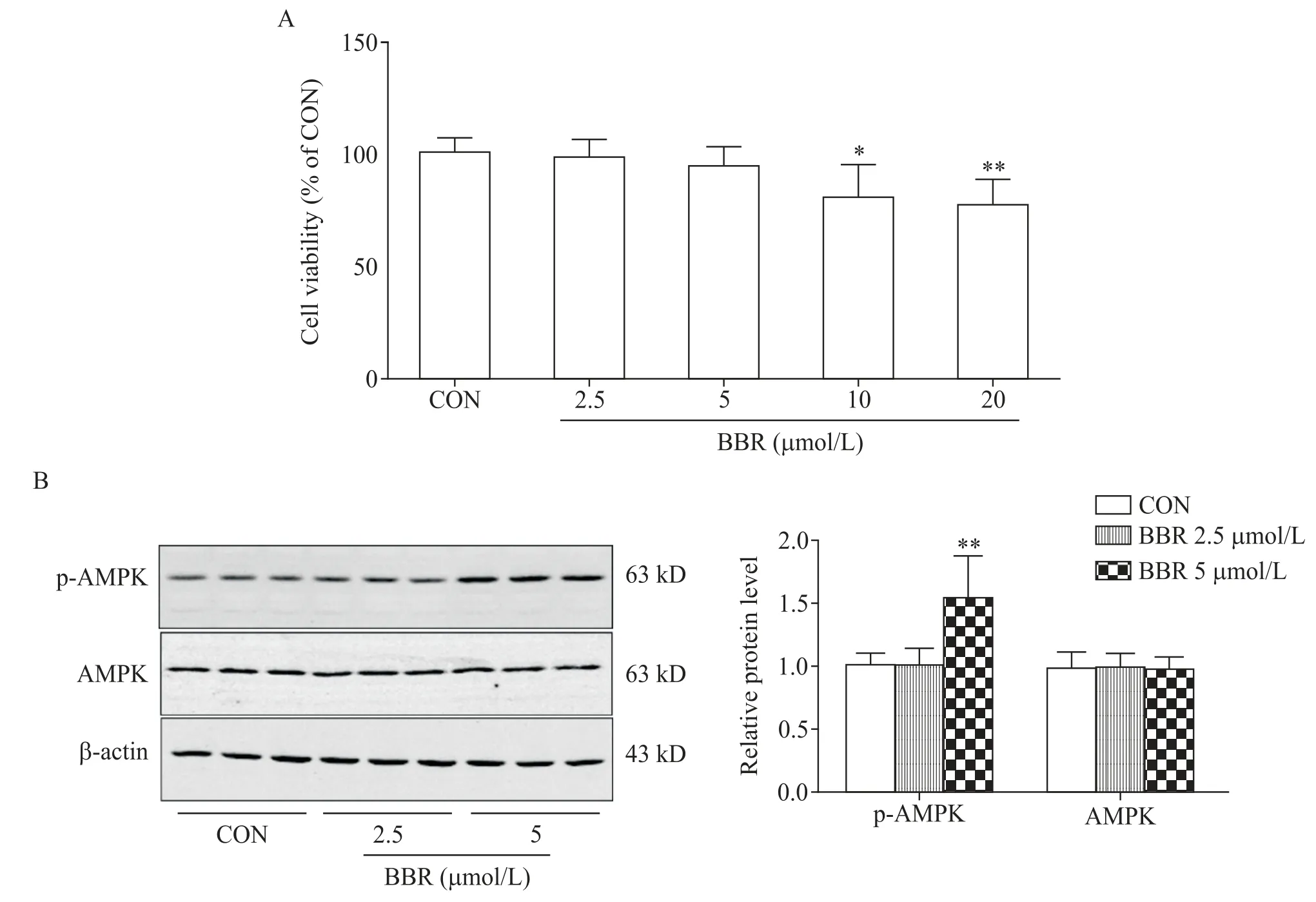
Figure 5. Effect of BBR on viability and AMPK activity in HEK293-Tau cells. A:the viability of HEK293-Tau cells after BBR treatment was tested by CCK-8;B:the levels of AMPK and p-AMPK in HEK293-Tau cells after BBR treatment were measured by Western blot. Mean±SD. n=3.*P<0.05,**P<0.01 vs CON group.图5 BBR对HEK293-Tau细胞活力和AMPK活性的影响
黄连素通过激活糖尿病模型动物的AMPK 活性,从而提高胰岛素的敏感性和降低全身肥胖[22]。神经母细胞瘤细胞和皮层神经元的实验结果显示,黄连素通过激活AMPK 减少Aβ 的生成并降低APP裂解酶(BACE1)的表达[13]。本研究的动物实验结果显示Hcy 模型大鼠的AMPK 活性降低、Tau 蛋白磷酸化水平增加,而黄连素治疗后,AMPK 的活性升高而且伴随着Tau 蛋白磷酸化水平降低。我们前期研究结果显示,激活AMPK 可以显著降低糖尿病小鼠海马区Tau 蛋白磷酸化水平[12]。为了阐明黄连素是否通过激活AMPK降低Tau蛋白磷酸化水平,我们用黄连素处理HEK293-Tau 细胞,同时给予(或不给)AMPK 抑制剂(Compound C),结果显示AMPK 抑制剂特异性的阻断黄连素对抗Tau 蛋白磷酸化的作用。该实验证实黄连素通过激活AMPK 而降低Tau蛋白的磷酸化水平。
GSK3β 是Tau 蛋白过度磷酸化的最重要的激酶,其活性位点Ser9 位点的磷酸化水平降低时,GSK3β 活性增加,引起 Tau 蛋白磷酸化水平增加[23]。PP2A 是降低 Tau 蛋白磷酸化水平的重要磷酸酶[24],其活性主要受催化亚基(PP2Ac)的酪氨酸307 位点(Tyr307)磷酸化水平和亮氨酸309 位点(Leu309)甲基化水平调节,磷酸化水平增加和/或甲基化水平降低使PP2Ac 酶活性降低[25]。课题组前期研究结果显示,AMPK 可抑制 GSK3β 的活性,同时降低 Tau 蛋白的磷酸化水平[12]。因此,我们检测 GSK3β 和 PP2Ac表达和其活性依赖的磷酸化水平。本研究结果显示,各组大鼠海马中GSK3β 活性没有变化,说明GSK3β 没有参与黄连素对抗Tau 蛋白磷酸化水平的过程中。动物实验结果显示Hcy 模型大鼠PP2Ac 活性显著降低,黄连素治疗后PP2Ac 活性显著升高,提示PP2Ac 参与黄连素对抗Hcy 诱导的Tau 蛋白过度磷酸化。细胞实验结果显示,黄连素和Compound C共同处理细胞时,AMPK 和PP2Ac 的活性降低,Tau蛋白磷酸化水平也增加,进一步证实黄连素通过激活AMPK 进而增加PP2Ac活性,从而阻止Tau蛋白过度磷酸化。
综上所述,黄连素通过调节AMPK/PP2Ac 通路,显著改善Hcy 大鼠的学习记忆能力和降低Tau 蛋白磷酸化水平,从而发挥神经保护作用。

Figure 6. BBR could reduce phosphorylation of Tau via AMPK/PP2Ac pathway in HEK293-Tau cells. Levels of pT205,pS396 ,p-AMPK,p-PP2Ac and DM-PP2Ac in HEK293-Tau cells after BBR with or without Compound C treating were measured by Western blot. Mean±SD. n=3.*P<0.05,**P<0.01 vs CON group;#P<0.05,##P<0.01 vs BBR group.图6 在HEK293-Tau细胞中,BBR通过AMPK/PP2Ac降低Tau蛋白的磷酸化水平
猜你喜欢
天津医科大学学报(2019年6期)2019-08-13
中国报道(2018年2期)2018-04-20
分析化学(2017年12期)2017-12-25
益寿宝典(2017年34期)2017-02-26
中老年健康(2016年10期)2016-11-19
绿色中国(2016年1期)2016-06-05
安徽医科大学学报(2015年9期)2015-12-16
中国中医药现代远程教育(2014年13期)2014-03-01
遗传(2014年3期)2014-02-28
