
封育年限对典型草原大针茅无性系构件组成与生长的影响
2021-12-29 11:23:06平伟梦赵凌平梁方晖谭世图赵芙蓉
草业科学 2021年11期
平伟梦,赵凌平,梁方晖,谭世图,王 冬,孙 平,赵芙蓉
(1. 河南科技大学动物科技学院,河南 洛阳 471000;2. 河南大学生命科学学院,河南 开封 475001)
黄土高原典型草原在维持生物多样性、发展畜牧业和调节气候等方面发挥着重要作用[1]。但近年来在气候变暖、不合理放牧和滥垦滥伐等因素的影响下,草地退化加剧,生物多样性降低,生态环境恶化严重[2]。在退化草地实施封育是缓解放牧压力及进行生态恢复的重要举措。封育对草地物种组成[3]、生物多样性和土壤养分[4]等影响显著,可对退化草地恢复起到一定的积极作用,已成为黄土高原地区草地管理的主要措施。封育引起草地环境条件变化,必然会改变各个种群在群落中的地位和作用。生态系统中的某一种群,特别是优势种群的消长必定会影响甚至改变整个生态系统的结构和功能[5]。
20 世纪70 年代初,Harper 和White[6]提出构件理论,Harper[7]依托构件理论对种群进行了群落及个体水平的划分,个体水平下的种群也可称为构件植物种群。构件是植物基本的形态学单位,既可以是由营养株繁殖形成的单个分株,也可以是单个植株的不同器官[8]。多年生无性系禾草的构件由生殖株和营养株组成[9],另外其无性系生长是指植物通过无性繁殖形成独立生存个体及占有物质资源的过程,此时构件结构及外部形态特征均会随复杂环境变化而变化,揭示了无性系植物表型可塑性对外部环境的响应规律。表型可塑性是指植物外部形态结构、生理特性及草地生物量对复杂环境的响应,通过研究无性系构件对不同环境的表型可塑性调节,有助于阐明无性系植物对环境的生态适应能力,了解草地生态系统中不同种群生长和繁殖策略变化,对草地群落发展动态和演替方向的研究具有重要意义[10]。近年来,构件理论在草地生态系统中已有大量研究,如林缘草地羽茅(Achnatherum sibiricum)[9]、松嫩平原根茎冰草(Agropyron michnoi)[10]、荒漠草原沙芦草(Agropyron mongolicum)[11]和内蒙古草原羊草(Leymus chinensis)[12]等植物,因此,以植物构件数量特征和生物量来研究资源分配特性已成为生态学的研究热点,且在无性系植物研究中应用广泛[10-13]。
大针茅(Stipa grandis)为禾本科针茅属多年生密丛旱生草本植物,杆高50~100 cm。该植物生长期为4 月 − 10 月,花果期为5 月 − 8 月。幼嫩时适口性良好,粗蛋白含量高,营养价值高于多数禾草,开花前家畜均喜食,尤其在春季萌发期,但在秋季成熟期其带稃颖果对牲畜有害。因具有较高的抗寒、耐旱性,适应性极强,大针茅是黄土高原典型优势种及建群种[14],在改善生态系统功能和维持畜牧业稳定方面起着重要作用。以往关于黄土高原大针茅的研究大多集中于生理生态特性[15]、光合特性[16]和根系格局变化[17]等方面。在黄土高原,随着封育年限的延长,退化草地逐渐演替为大针茅群落,这必然与大针茅种群生长和繁殖策略有关。尽管已有研究报道过根茎型羊草[12]、疏丛型狗牙根(Cynodon dactylon)[18]等多种禾草的无性系构件,但关于密丛型禾草大针茅营养种群结构、分株生长和物质分配的研究却鲜有报道。因此,本研究以黄土高原典型草原的放牧地、封育10 年草地和封育20 年草地为研究对象,测定大针茅无性系不同构件数量特征和生物量分配特征,探究封育措施对大针茅构件资源分配的影响,以期为黄土高原退化草地的恢复管理和可持续利用提供依据。
1 材料与方法
1.1 研究区概况
研究区设在海拔1 800 − 2 180 m 的宁夏回族自治区固原市云雾山国家草原自然保护区(36°13′ −36°19′ N,106°24′ − 106°28′ E),属温带大陆性半干旱气候。年均气温5 ℃,1 月气温最低,为−14 ℃左右,7 月气温最高,为25 ℃左右。无霜期112~140 d。年均降水量400~450 mm,其中70%降水量集中在7 月 − 9 月。年均蒸发量约为1 440 mm。土壤为砂壤土,pH 8.0~8.6[19]。植被以多年生无性系草本植物为主,包括优势种大针茅和本氏针茅(Stipa bungeana),伴生种百里香(Thymus mongolicus)、铁杆蒿(Artemisia sacrorum)、厚穗冰草(Agropyron cristatum)、二裂委陵菜(Potentilla bifurca)和阿尔泰狗娃花(Heteropappus altaicus)等[20]。
1.2 研究方法
2018 年8 月,选取1 个放牧草地和两个封育草地作为试验样地。封育草地是由云雾山草原管理站分别于1998 和2008 年实施围栏封育的,在取样时已经分别封育20 和10 年。封育20 和10 年草地面积约为10 和5.5 hm2。在每个样地随机挖取大针茅30 丛(株),每丛间距100 m 以上。挖取前测定针茅基部丛径,以3 个不同方向测定的平均值作为丛径大小。编号完成后,挖出置于密封袋内,带回实验室进行处理。生殖枝和营养枝的区分以是否有花序或底部分蘖芽为标准。将针茅分株从基部分蘖处掰开,对分蘖芽、生殖枝及营养枝计数,测定生殖枝和花序高度后,分离生殖枝与花序,分别装袋。75 ℃烘干至恒重,测定生殖枝、营养枝和花序生物量。大针茅根系生物量采用单位面积挖掘法(0.4 m × 0.4 m ×0.3 m)测定,根据相连的大针茅地上部分分辨出大针茅根系,去除大块土壤及其他植物根系后,冲洗干净,烘干并称重。变异系数(coefficient of variation,CV)是标准差与平均值之比,可反映出各草地中大针茅同一构件的相对变异度。根系生物量是根系干重与根系采集面积之比。
1.3 数据分析
采用SPSS 25.0 对大针茅不同构件数量特征丛径、总枝数、生殖枝数、营养枝数、分蘖芽总数、生殖枝高度、花序高度、地上总生物量、生殖枝生物量、营养枝生物量、花序生物量、根系生物量进行单因素方差分析(One-way ANOVA)和Pearson 相关性分析,运用Origin 2018 制图。
2 结果与分析
2.1 不同封育年限下大针茅构件数量特征变化
封育年限对植株丛径、总枝数、生殖枝数、营养枝数、分蘖芽总数、生殖枝高度、花序高度、地上总生物量、营养枝生物量和花序生物量均有显著影响(P<0.05),但对生殖枝生物量和地下根系生物量影响不显著(P> 0.05) (表1)。与放牧地相比,封育10 年仅显著提高了植株丛径、生殖枝高度和花序高度(P<0.05),分别增加了38.9%、66.1%和68.6%;封育20 年草地中的大针茅丛径、总枝数、生殖枝数、营养枝数、分蘖芽数、生殖枝高度、花序高度、地上总生物量、营养枝生物量和花序生物量均显著增加(P<0.05),分别增加了76.5%、294.0%、176.6%、307.2%、159.6%、74.8%、98.0%、414.9%、894.4%和297.9%。与封育10 年草地相比,封育20 年草地植株的丛径、总枝数、生殖枝数、营养枝数、分蘖芽数、地上总生物量、营养枝生物量、花序生物量均显著增加(P<0.05),分别增加了27.0%、283.2%、91.2%、314.2%、83.8%、185.1%、316.3%和133.4%。
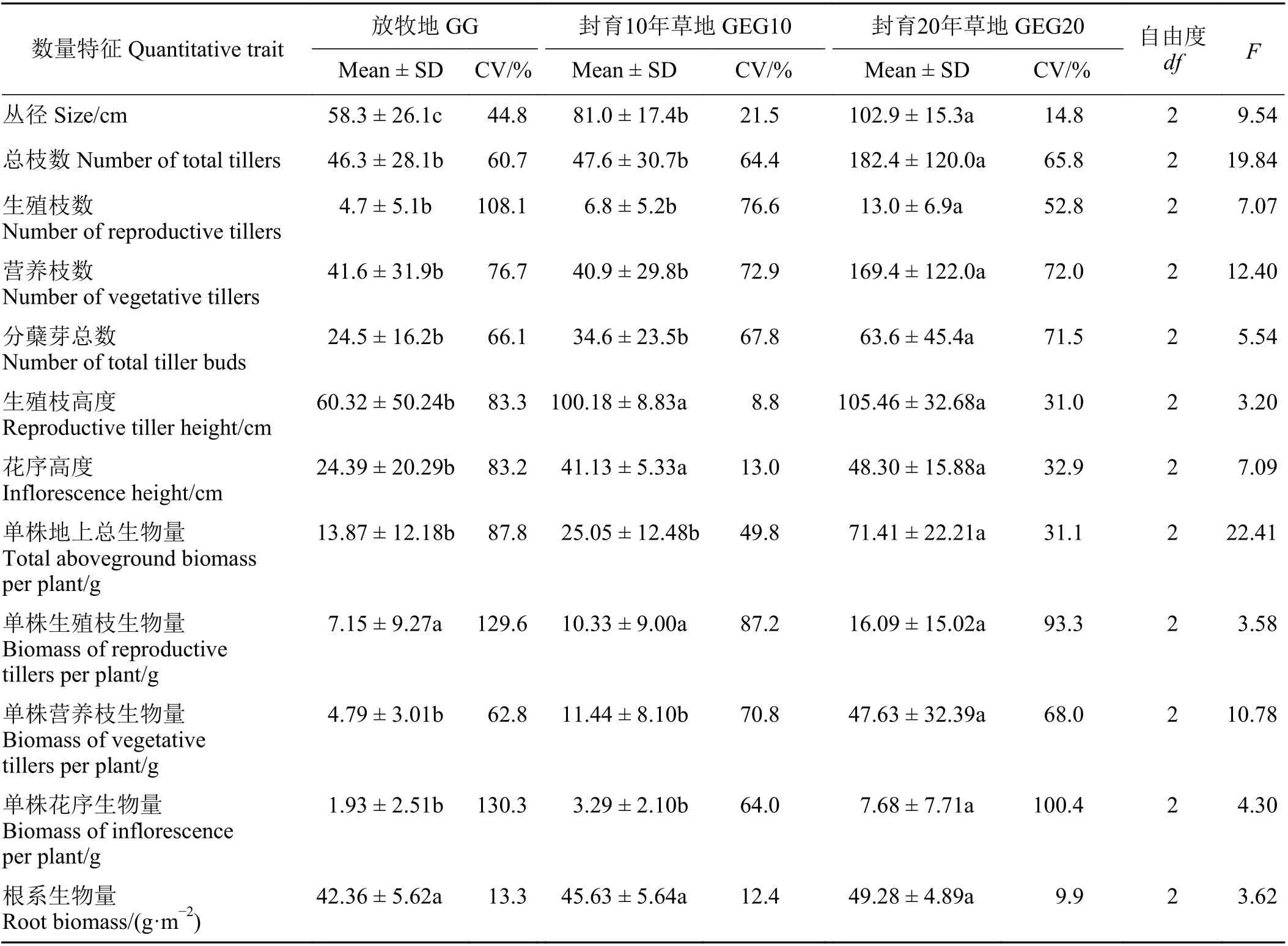
表1 不同封育年限下大针茅各构件数量特征Table 1 Quantitative traits of the modules in Stipa grandis clones under different grazing exclusion years
随封育年限增加,大针茅丛径变异系数呈降低趋势。放牧地大针茅生殖枝数、生殖枝高度、花序高度、地上总生物量的变异系数均超过80%,具有较强变异性,封育降低了大针茅生殖枝数、生殖枝高度、花序高度、地上总生物量、生殖枝生物量和花序生物量的变异系数。3 个样地中生殖枝生物量和花序生物量的变异系数普遍大于营养枝生物量。
2.2 不同封育年限下大针茅无性系构件的地上生物量分配变化
在放牧地,大针茅花序生物量、营养枝生物量和生殖枝生物量分别占地上总生物量的13.9%、34.5%和51.6%。封育10 年草地中,大针茅花序生物量、营养枝生物量和生殖枝生物量分别占地上总生物量的13.1%、45.7%和41.2%,封育20 年的草地则分别占10.8%、66.7%和22.5%。封育降低了生殖枝生物量比例而提高了营养枝生物量比例,在草地封育20 年时更为明显,但对花序生物量分配比例无明显影响(图1)。

图1 大针茅无性系不同构件的地上生物量分配Figure 1 Aboveground biomass allocation of differentmodule structures of Stipa grandis clones
2.3 不同封育年限下大针茅无性系不同构件单株生物量变化
封育10 年和20 年草地大针茅单株营养枝生物量显著高于放牧地(P< 0.05),但封育10 年和20 年草地间差异不显著(P> 0.05)。另外,3 个样地的单株生殖枝生物量和单株地上总生物量差异不显著(P> 0.05) (图2)。

图2 大针茅无性系不同构件平均单株生物量Figure 2 Average biomass per tiller of different module structures of Stipa grandis clones
2.4 大针茅无性系各构件数量特征关系
在9 个数量特征的相关性分析(表2)中,呈显著正相关关系的共有21 对(P< 0.05),呈显著负相关关系的共有2 对(P< 0.05)。其中大针茅丛径与地上总生物量、生殖枝数、生殖枝生物量、花序生物量极显著正相关(P< 0.001),与营养枝生物量极显著正相关(P< 0.01),与总枝数显著正相关(P< 0.05)。地上总生物量与生殖枝数、营养枝数、总枝数、营养枝生物量极显著正相关(P< 0.001),与分蘖芽数、生殖枝生物量、花序生物量极显著正相关(P< 0.01)。总枝数与地上总生物量、营养枝数、分蘖芽数和营养枝生物量极显著正相关(P< 0.001)。分蘖芽数与营养枝数、总枝数、营养枝生物量极显著正相关(P<0.001),与地上总生物量极显著正相关(P< 0.01)。总之,营养枝在大针茅生长阶段起到重要作用,是其占据空间及物质资源的关键。

表2 大针茅无性系构件数量特征间的相关性分析Table 2 Correlation coefficient of different module forms of Stipa grandis clones
3 讨论
封育措施是退化草地恢复与重建的良好驱动力。封育后土壤理化性质改善和养分积累对草地植物组成和竞争格局会产生一定影响[21]。封育和放牧可对植物种群进行年龄结构、种间关系、空间格局和表型可塑性等多方位的调控。本研究主要探讨了封育和放牧对大针茅无性系构件结构和生物量分配的影响。数量特征的显著改变均可在构件层次上反映出植物对资源的利用能力和对生境的适应能力[22]。研究结果表明,封育措施提高了大针茅的丛径、生殖枝高度和花序高度,特别是封育20 年显著提高了大针茅丛径、总枝数、生殖枝数、营养枝数、分蘖芽数、生殖枝高度、花序高度、地上总生物量、营养枝生物量和花序生物量。在封育草地,植物未被家畜践踏、采食,大针茅可供食用部分和用于繁殖的构件生物量逐渐增高,植株高度和丛径明显增高,繁殖能力也随之增强,逐步发育成草地群落优势种,草地可利用价值和更新能力得到提升。在高寒草地,长期放牧导致优良牧草高度下降,生殖枝减少[23];在草甸草原,长期放牧导致优良牧草株高和个体地上生物量分别降低了76.8%和89.8%[24];上述研究结果与本研究结果一致。另外在不同封育年限下,大针茅根系生物量差异不显著,这可能是由于封育时间过长,枯落物层较厚,草地群落趋于稳定,大针茅根系生物量变化不显著,但放牧草地较低可能是因为植物地上部分补偿性生长[17]。
变异系数通常用来评定数据的离散程度,反映植物表型可塑性[8]。较高的表型可塑性是植物适应复杂环境,占据生境的主要原因[25]。放牧地中大针茅生殖枝数、生殖枝高度、花序高度、地上总生物量的变异系数均超过80%,而封育措施降低了这些指标的变异系数,说明封育措施降低了大针茅的表型可塑性,使大针茅株高和丛径变大,均匀一致。但李西良等[24]指出羊草对表型矮小化具有保守性,其茎、叶性状对3 年封育措施无明显响应,与本研究结果不一致。这可能是由于封育年限长短不同引起的,短期封育还未能明显改变植物的表型可塑性,但长期封育引起了植物表型可塑性的改变。本研究中大针茅对表型矮小化不具有保守性。在放牧地,大针茅采取地上生物量和高度降低的适应策略,降低了被家畜采食的机率。汪诗平等[26]也指出植物随着放牧强度的增加往往采取降低高度和生物量的避食机制。植物的矮化型变是草地群落结构和功能改变的重要触发机制。放牧干扰下植物矮化型变可能受环境、生理和基因方面的影响,但仍需进一步深入研究。另一方面,封育和放牧措施对生殖枝数、生殖枝高度和生殖枝生物量的变异系数影响较大,说明大针茅有性繁殖的可塑性较高。据报道,在放牧地来源于种子的实生苗生长为成年植株的比例较高,而封育草地中植物每年可产生大量种子形成种子库,但因其光照和空间资源竞争能力较弱,来源于种子的实生苗生长为成年植株的可能性较小,导致草地植物的繁殖更新以无性繁殖为主,以有性繁殖为辅[27]。
分蘖是克隆植物大针茅进行无性繁殖的主要方式[28],分蘖芽数是评定其营养繁殖力的重要指标[29],是无性系扩展和持续更新的基础。分蘖芽数和变异程度可预示下一个生长季种群的兴衰[30-31]。本研究中封育20 年草地的植株分蘖芽数显著高于其他草地,且变异系数有所增加,可推测出在下一个生长季,各样地中大针茅无性系营养繁殖仍具较大差异。退化草地封育20 年明显保护了大针茅繁殖更新的基础,有利于草地恢复。
生物量是评定草地生态系统结构和功能优劣的直接表现,反映了植物适应环境能力的强弱。植物构件生物量的改变是该植物和周围环境及不同草地管理措施共同作用的结果。本研究中随封育年限增加,大针茅营养枝生物量比例显著增加,生殖枝生物量比例降低,花序生物量比例基本稳定。这说明封育减少了家畜对大针茅地上部分的采食量,同时其无性繁殖能力显著增强,营养枝生物量比例随之增高。这与高寒草甸封育后相似,放牧压力的消失,使植物对光照的竞争增加,从而对光合器官的生物量投入增加,减少了对生殖器官的投入[32]。此外,封育后大针茅生殖株单株生物量明显较高,这在大针茅生长发育、繁殖扩张、维持优势种地位中具有一定作用[8]。但以无性繁殖为主的大针茅,营养枝总生物量占主体,是草地生产力的主要来源。这进一步说明,植物各构件生物量在不同封育年限下的资源分配存在着明显的竞争关系,通过生长发育和繁殖更新,逐步趋于稳定,形成适应当地环境的理想分配模式。
丛径是大针茅无性系大小的主要测定指标,本研究中,随丛径增加,大针茅无性系构件数量及生物量均增加,无性系各构件数量特征间存在极显著关系。有研究发现,短花针茅(Stipa breviflora)的基丛径与地上生物量呈显著的正相关关系[33];小花碱茅(Puccinellia tenuiflora)的生殖株数量和生物量与丛径也呈正相关关系[34];松嫩平原上的羽茅[8]、硬质早熟禾(Poa sphondylodes)[35]和野大麦(Hordeum brevisubulatum)[36]等禾草无性系构件的相关研究结果也是如此。而大针茅无性系地上总生物量与营养枝数、营养枝生物量、总枝数、生殖枝数极显著正相关,表明了地上总生物量主要与营养枝数和营养枝生物量有关,这主要是因为大针茅以无性繁殖为主,营养枝枝数及生物量比例较高。总之,大针茅无性系构件间的相关性变化在生长繁殖和资源分配中遵循着某种规律,良好地响应环境变化,在促进群落更新和提高草地生产力方面发挥着重要作用。
4 结论
随封育年限增加,大针茅无性系构件的生殖枝数及单株生物量的变异系数大于营养枝,说明其表型可塑性较高,有性繁殖变异度高,无性繁殖稳定。营养枝生物量比例随封育年限增加而显著增加,生殖枝生物量比例降低,花序生物量比例基本稳定。但营养枝枝数及生物量均占优势,其在群落中占据空间及物质资源的能力远大于生殖枝,这为大针茅及无性系植物的进一步研究奠定了一定的理论基础。短期封育提高了大针茅的生殖分配,使大针茅迅速成为群落优势种,而长期封育使大针茅更依靠无性繁殖进行种群更新。
猜你喜欢
草地学报(2022年9期)2022-09-29 11:22:48
天津农林科技(2022年2期)2022-04-19 10:48:14
河南畜牧兽医(2022年3期)2022-04-13 09:12:58
——植物学教材质疑(六)
生命科学研究(2022年1期)2022-03-14 07:50:40
绿色科技(2021年10期)2021-06-23 03:13:06
草业学报(2019年2期)2019-02-25 01:57:16
山东农业科学(2018年7期)2018-10-22 07:01:10
南风(2017年34期)2018-01-16 07:29:09
科学种养(2017年6期)2017-06-13 08:42:04
科学种养(2017年5期)2017-05-17 10:18:50
