
珠江河口棘头梅童鱼生物学特征的年代际变化
2021-12-22 05:56熊朋莉陈作志邱永松范江涛徐姗楠
南方水产科学 2021年6期
熊朋莉 ,陈作志,侯 刚,张 帅,邱永松,范江涛,徐姗楠
(1. 中国水产科学研究院南海水产研究所/农业农村部外海渔业开发重点实验室,广东 广州 510300;2. 上海海洋大学水产与生命学院,上海 201306; 3. 广东海洋大学水产学院,广东 湛江 524088)
棘头梅童鱼 (Collichthys lucidus) 属鲈形目、石首鱼科、梅童鱼属,为中国河口、沿岸水域重要经济鱼类,其适温、适盐范围广,在中国沿海河口均有分布,是河口生态系统的重要组成部分[1-2]。近年来,在人类捕捞、栖息地碎片化及全球气候变化导致的升温等因素影响下,全国河口、沿岸水域棘头梅童鱼的生长发育和资源状况受到威胁,个体小型化严重[3-5]。种群生物学研究是渔业资源评估的重要基础,可为渔业资源管理提供数据支撑。最新研究发现,中国沿海棘头梅童鱼可分为南北2个地理群体,在渔业资源开发和管理上须区别对待[6],国内有关棘头梅童鱼渔业生物学特征研究主要集中于种群结构、生长、摄食习性、性腺发育等方面[3-4,7]。现有研究多采用现状调查数据来分析种群生物学基本特征,有关种群生物学特征长期变化的研究仍显不足,因此有必要加强该种群的基础生物学研究。
珠江河口位于咸淡水交汇处,生态环境优越,是南海区重要的鱼类种质资源库,具有重要的经济地位和生态保育价值。目前,有关珠江河口棘头梅童鱼的相关报道较少,主要开展了其资源量及开发状态评估和基于耳石日轮相关的研究[1-2,8-9]。本文通过对比珠江河口1986—1987年与2018—2019年的渔业生物学调查数据,对棘头梅童鱼群体组成、生长和繁殖等生物学特征进行初步评估,分析了其群体生物学特征的年际变化,以期为珠江河口棘头梅童鱼资源的合理利用及生态保护提供基础数据和支撑。
1 材料与方法
1.1 数据来源
调查数据来自于中国水产科学研究院南海水产研究所于1986—1987年1—4月、2018年1—2月和2019年3—4月在珠江河口进行的渔业资源调查(图1)。其中1986—1987年样本的采样工具为定置网,2018—2019年样本的采样工具为刺网 (由于2个年代的采样工具不同,仅选取体长大于50 mm的个体作为本次研究对象)。棘头梅童鱼样品冷冻保存,带回实验室进行生物学测定,测定内容包括体长 (精度1 mm)、体质量 (1986—1987年和2018—2019年测定精度分别为0.1和0.01 g)、性别、性腺成熟度、摄食等级等。性腺成熟度划分为Ⅰ —Ⅵ期,摄食等级划分为0—4级[10-11]。共选取1986—1987年调查期间测定的棘头梅童鱼612尾,2018—2019年测定的440尾。
图1 珠江河口渔业资源调查范围Figure 1 Survey area of fishery resources in Pearl River Estuary
1.2 数据处理
1.2.1 体长、体质量分布 体长和体质量分别按照10 mm和10 g的组距进行分组,并将超过样本总尾数20%的体长组和体质量组设为本研究的优势体长组和优势体质量组。采用单因素方差分析其群体平均体长和平均体质量的性别差异,并利用非参数检验判断雌、雄个体以及不同年代际体长和体质量分布的显著性差异。应用独立样本t-检验分析平均体长和平均体质量在不同年份间的差异。体长和体质量关系拟合采用幂函数:

式中:W表示体质量 (g);L表示体长 (mm);a、b分别代表条件因子和异速生长参数[10,12-13]。采用协方差分析,判断雌、雄体长-体质量曲线有无差异。并将异速生长参数b与常数3进行比较,判断棘头梅童鱼群体在不同年代际的生长状态[3]。
1.2.2 性别比例 统计不同时期、不同体长组的雌、雄个体尾数,分析性别比例在不同时期、季节、体长组的变化。
1.2.3 50%性成熟体长 将性腺发育达到Ⅳ期及以上的个体视为性成熟个体[14-15]。筛选出样本中的雌性性成熟个体,按照体长5 mm的组距进行分组。采用非线性回归拟合反正弦平方根形式的逻辑斯蒂模型得到50%性成熟体长的95%置信区间[16-17]。
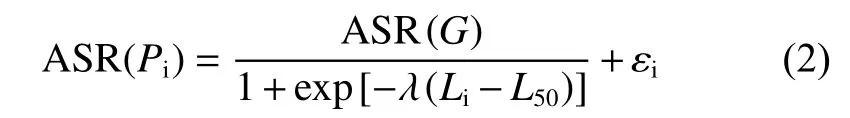
1.2.4 摄食等级 统计棘头梅童鱼在不同时期的摄食等级,利用独立样本t-检验判断摄食等级在不同季节和性别是否存在显著性差异以及不同年份间平均摄食等级之间的差异。
1.2.5 肥满度 根据测定样本计算棘头梅童鱼群体肥满度大小[18]。利用独立样本t-检验分析不同年份间平均肥满度的差异。

式中:K为肥满度指数;W为体质量 (g);L为体长 (cm)。
2 结果
2.1 体长-体质量分布
1986—1987年调查中,棘头梅童鱼平均体长为 (95±13) mm,优势体长组为 90~110 mm (54.41%),平均体质量为 (20.11±7.39) g ,优势体质量组为10~30 g (81.67%)。雌、雄体长分布、体质量分布、平均体长、平均体质量均有显著差异 (P<0.05);2018—2019年调查中,棘头梅童鱼平均体长为 (93±17) mm,优势体长组为90~110 mm (41.81%),平均体质量为 (22.19±12.43) g,优势体质量组为0~30 g (74.09%)。雌、雄平均体长有显著差异(P<0.05),而雌、雄平均体质量、体长分布和体质量分布均无显著差异 (P>0.05)。不同年份调查的体长分布、体质量分布特征见表1,不同年份调查的体长组、体质量组分布频率见图2。
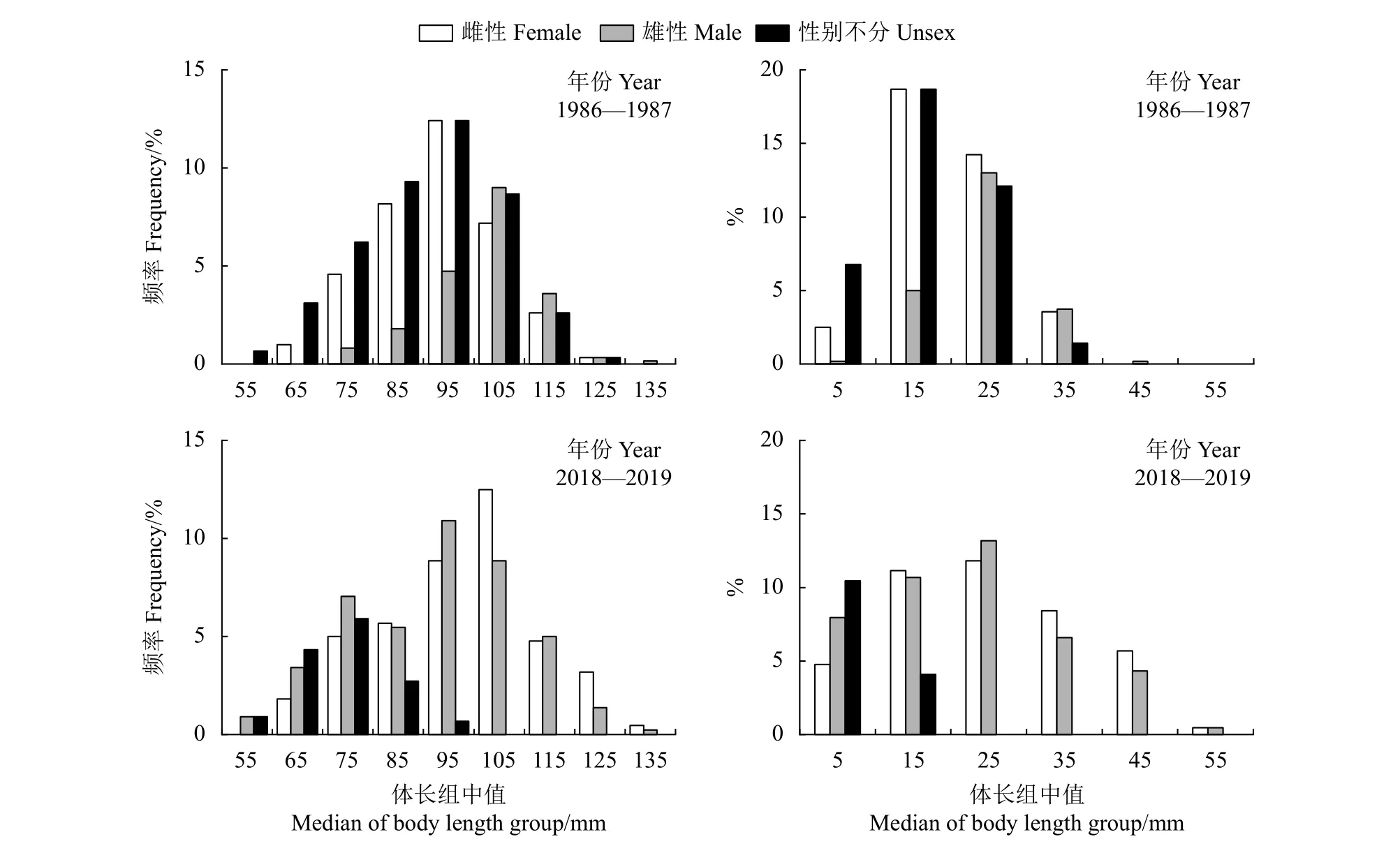
图2 珠江河口棘头梅童鱼体长、体质量分布Figure 2 Distribution of body length and body mass of C. lucidus in Pearl River Estuary
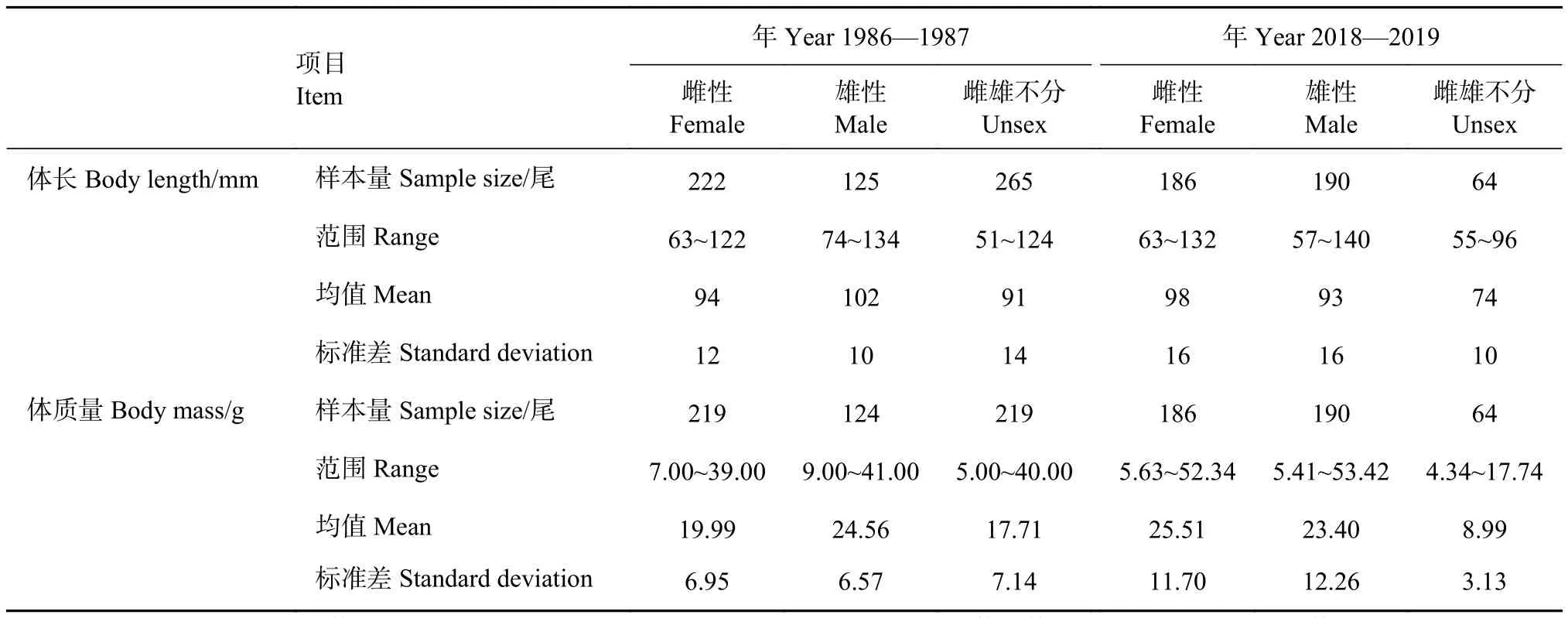
表1 棘头梅童鱼体长体质量分布特征Table 1 Characteristics of body length and mass distribution of C. lucidus
1986—1987年和2018—2019年的平均体长无显著差异 (P>0.05),平均体质量存在显著性差异(P<0.05)。2个时期的体长分布差异显著 (P<0.05),体质量分布无显著差异 (P>0.05)。
2.2 体长-体质量关系
珠江河口棘头梅童鱼雌、雄及全部群体的体长-体质量关系符合幂函数关系(图3)。协方差检验结果表明,不同年份体长-体质量关系中的雌、雄差异均不显著 (P>0.05)。1986—1987年,其雌、雄群体均呈负异速生长 (b<3),而2018—2019年其雌、雄群体均呈正异速生长 (b>3)。
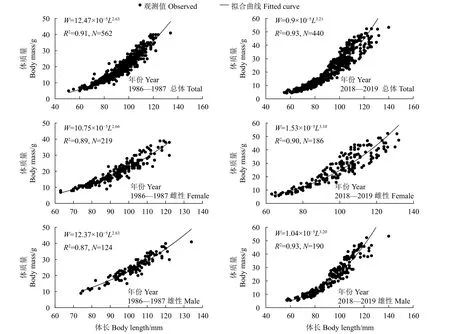
图3 珠江河口棘头梅童鱼体长-体质量关系Figure 3 Relationship between body length and body mass of C. lucidus in Pearl River Estuary
2.3 性别比例
1986—1987年棘头梅童鱼样本雌、雄性比为1.72∶1,不符合 1∶1 (P<0.05);2018—2019 年其性别比例符合1∶1 (P>0.05)。此外,1986—1987年雄性个体占比小于2018—2019年,2018—2019年雄性个体占比大于1986—1987年。
不同体长组中,棘头梅童鱼群体性比也会发生相应的变化 (图4)。体长介于60~90 mm,1986—1987年雌性个体占比随着体长组的增加而降低,2018—2019年样本中雌性占比随着体长组的增加而上升。棘头梅童鱼群体在不同的季节也呈现不同的性比。1986—1987年春季 (3—4月) 性比为1.51∶1,冬季 (1—2 月) 为 2.78∶1;2018—2019年春季性比为1∶1.13,冬季为2.03∶1。
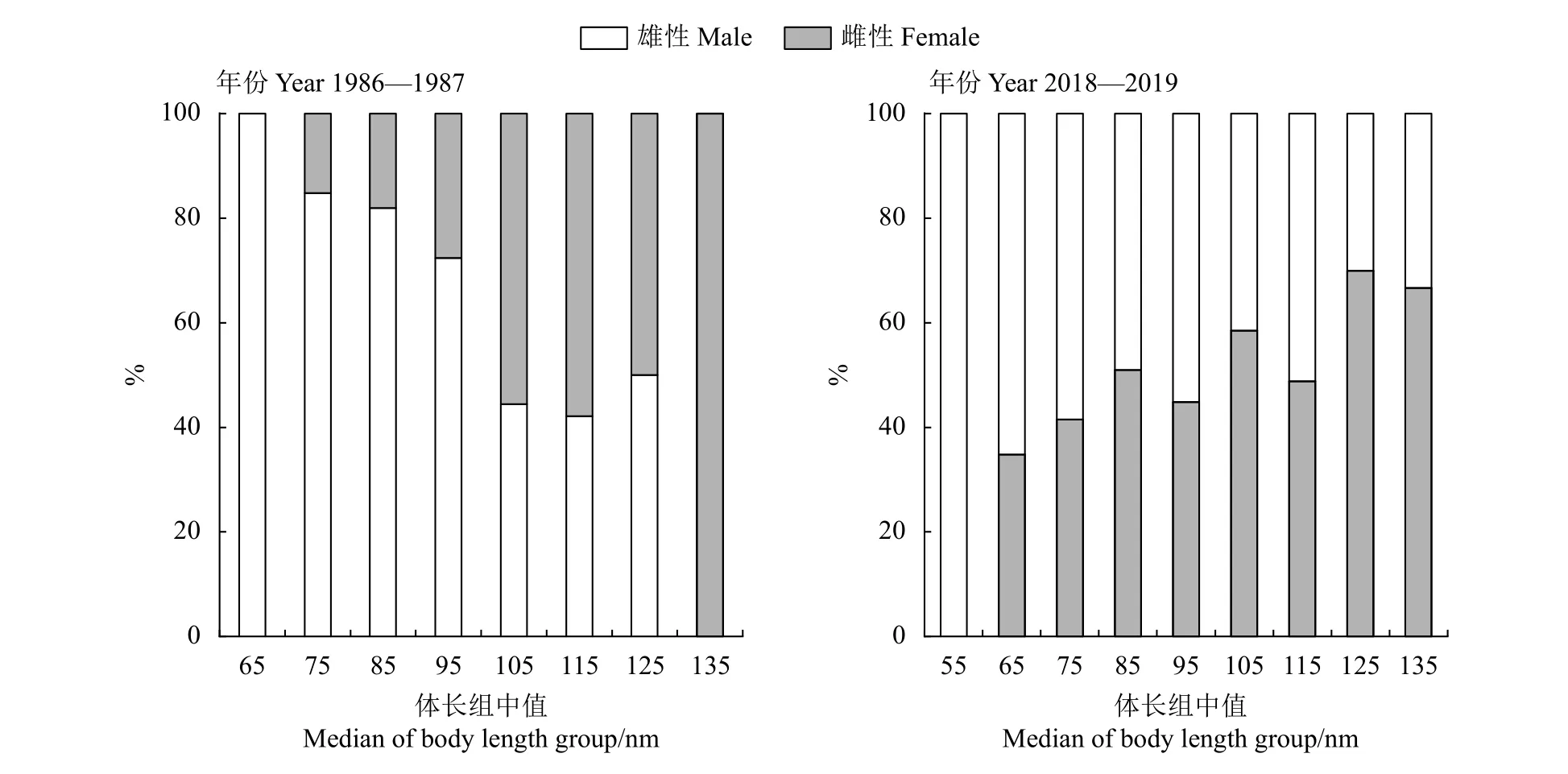
图4 珠江河口棘头梅童鱼各体长组性别比例Figure 4 Sex ratio of C. lucidus of different body length groups in Pearl River Estuary
2.4 50%性成熟体长
1986—1987年和2018—2019年棘头梅童鱼50%性成熟体长分别为112 mm (105~118 mm为95% 置信区间,后同) 和 88 mm (78~97 mm) (图 5)。
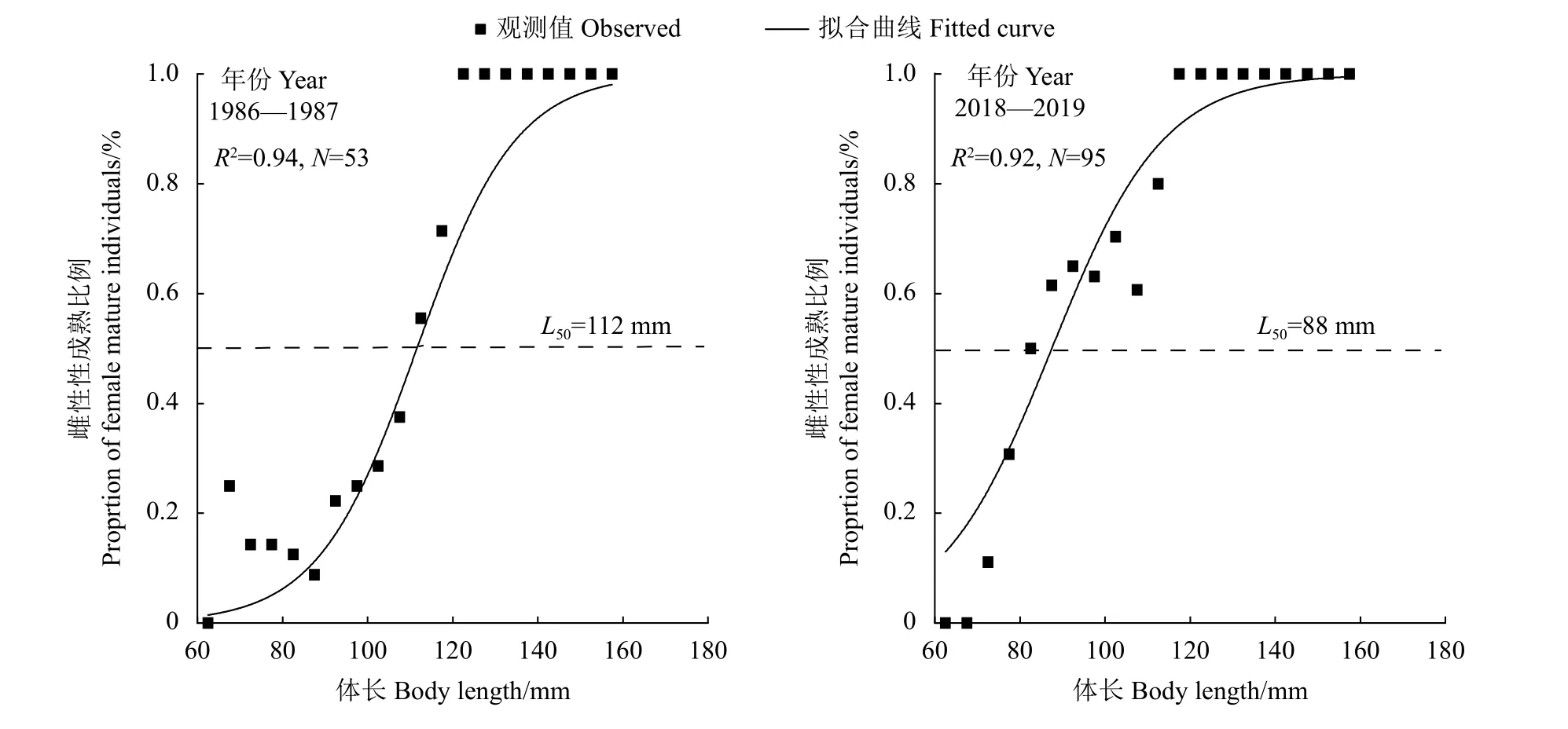
图5 珠江河口棘头梅童鱼雌性个体性成熟比例观测值与ASR模型拟合的逻辑斯蒂曲线Figure 5 Observed proportion of mature individuals and logisitic curve fitted by ASR models of C. lucidus in Pearl River Estuary
2.5 摄食等级和肥满度
珠江河口棘头梅童鱼摄食等级以1和2级为主,与1986—1987年相比,2018—2019年的摄食等级中2和3级占比上升 (图6)。1986—1987年和2018—2019年空胃率分别为8.52%和17.39%。1986—1987年和2018—2019年雌、雄群体平均摄食等级差异均显著 (P<0.05)。1986—1987年雌、雄群体平均肥满度无显著性别差异 (P>0.05)。2018—2019年雌、雄群体平均肥满度差异显著(P<0.05)。1986—1987年棘头梅童鱼群体在春、冬季平均摄食等级无显著差异 (P>0.05)。而2018—2019年春、冬季平均摄食等级差异显著 (P<0.05),且冬季摄食强度高于春季 (图7)。
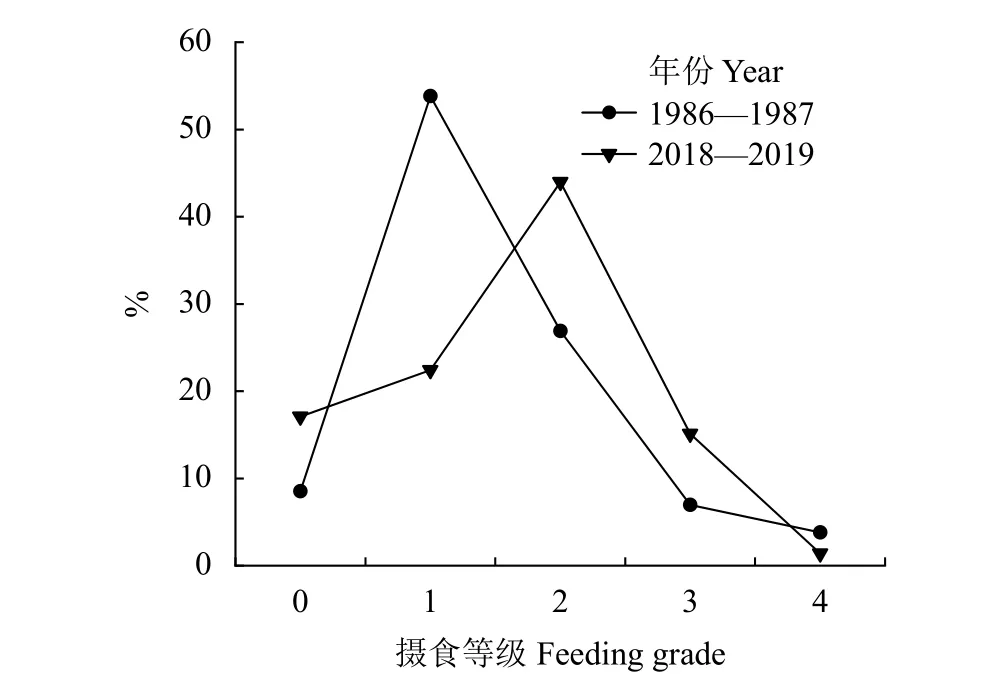
图6 珠江河口棘头梅童鱼摄食等级分布Figure 6 Percentage of feeding grades of C. lucidus in Pearl River Estuary
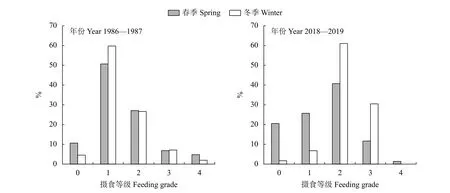
图7 珠江河口棘头梅童鱼摄食等级的季节变化Figure 7 Seasonal variation of feeding grades of C. lucidus in Pearl River Estuary
棘头梅童鱼雌、雄和全部群体的平均肥满度和平均摄食等级均呈增加趋势 (表2)。2018—2019年平均肥满度和摄食等级均显著大于1986—1987年 (P<0.01)。
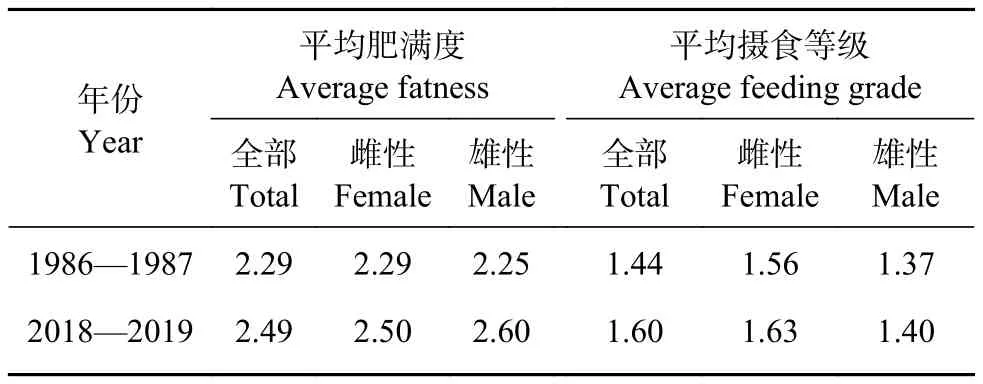
表2 棘头梅童鱼平均肥满度和其平均摄食等级Table 2 Average condition factor and feeding grade of C. lucidus
3 讨论
鱼类生物学特征是反映其栖息地环境变化的重要指标。研究表明,与20世纪80年代相比,当前珠江河口棘头梅童鱼L50呈降低趋势。基于目前的研究数据,初步推断珠江河口棘头梅童鱼出现了性早熟现象。水温是造成性腺发育提前的重要因素。水温升高,新陈代谢加快,促进性腺发育提前。棘头梅童鱼L50下降的过程中,其平均摄食等级上升。因此,本研究认为珠江河口棘头梅童鱼性腺发育提前可能是由于该水域水温升高且饵料充足所致。此外,对南海多个近海经济种类的研究表明,过度捕捞也会诱导种群性腺发育提前[19-22]。
珠江河口棘头梅童鱼群体平均肥满度上升。肥满度可以密切反映鱼类的营养状况,一般而言,鱼类营养条件越好,其肥满度越高[12,18,23]。林坤等[24]、袁梦等[25]研究表明,珠江河口近岸海域生态网络密度下降、群落结构趋于弱化。笔者认为,珠江河口区域在高强度的捕捞胁迫下导致鱼群的资源密度下降,捕食、栖息地等种内竞争或种间竞争压力减少,饵料相对充足[21,26-27],鱼群的稀疏效应对棘头梅童鱼肥满度的上升起到促进作用。
相关研究表明,杭州湾北部、闽江口等区域的棘头梅童鱼已出现小型化现象[3,28]。一般而言,大部分鱼类在经过较大的捕捞强度以及环境因子的胁迫后,其种群的表现性状会产生适应性响应[29-30],其中体长会表现出较为明显的表化[20,29]。本研究中棘头梅童鱼优势体长组占比降低,但2个年代际的平均体长并无显著差异。因此,珠江河口棘头梅童鱼是否出现小型化现象还需进一步扩大样本量研究其生长参数。
本研究发现,2018—2019年珠江河口棘头梅童鱼产卵群体中,雄性多于雌性,雌雄比为1∶1.42,且在性别比例中,雄性个体数占比也高于雌性。当前该海域棘头梅童鱼性比模式与1986—1987年的调查结果以及杭州湾北部湾棘头梅童鱼的调查结果相反[3]。但国内一些关于鱼类生物学的研究中,性别比例也有呈现出雄性个体较高的现象,如大鳞裂峡鲃 (Hampala macrolepidota)、南海北部陆坡海域中瓦氏眶灯鱼 (Diaphus watasei)等[31-32]。鱼类的性别比例会因栖息环境、个体大小以及生活阶段的不同而发生相应变化[11]。产卵期间雄性多于雌性的性比模式有利于提高卵子的受精率[11]。研究表明a与鱼类群体的生活环境关系密切[33]。本研究评估的1986—1987年和2018—2019年a有较大差异,表明这2个时期间珠江河口的环境发生了较大变化。与何宝权等[1]的研究结果相比,本研究1986—1987年a略有上升,b略有下降,其原因可能是样本的选取范围和样本量不同。何宝全等选取300个1986年样品的测量数据进行体长-体质量关系拟合,而本研究选取的是1986—1987年1—4月的全部样本数据。但与本研究中2018—2019年棘头梅童鱼体长-体质量关系的研究结果相比,a、b呈相同的变化趋势。因此,本研究认为棘头梅童鱼性比模式变化较大的原因可能是近年来珠江河口海域海表温度升高、水环境变化迅速等导致其为适应该海域栖息环境的变化而进行的种群自我调节[34-35]。
珠江河口棘头梅童鱼摄食等级具有显著的季节变化。通过研究2018—2019年调查数据发现,棘头梅童鱼冬季的平均摄食等级和饱食率 (摄食等级达到3级和4级的个体数占比) 均高于春季,且春季的空胃率高于冬季。该结果与棘头梅童鱼群体春季摄食强度高于冬季的已有研究结果存在差异[4,7,36],原因可能是水域环境不同导致鱼类群体摄食强度的季节变化出现差异。
此外,珠江河口棘头梅童鱼性成熟个体的平均摄食等级要低于群体平均摄食等级,群体摄食强度冬季高于春季,推测可能是春季为其主要繁殖期,个体达到性成熟的比例要高于冬季,性成熟个体由于性腺发育占据体腔空间,导致春季平均摄食强度低于冬季[3]。
棘头梅童鱼是珠江河口关键物种,其群体性早熟现象一定程度反映出整个珠江河口渔业资源的衰退。珠江河口虽设立了经济鱼类繁育保护区、中华白海豚国家级自然保护区等多个自然保护区,渔业管理部门对自然保护区采取了禁渔区、禁渔期、生态修复等渔业资源保护措施[37-38],但由于管理不善,珠江河口渔业资源仍面临严重威胁。因此,今后渔业管理部门应进一步完善渔业管理措施,严格控制渔业资源捕捞压力,加大执法力度,以便能更好地恢复珠江河口经济鱼类的资源量。
猜你喜欢
四川蚕业(2022年2期)2022-11-19
天天爱科学(2022年9期)2022-09-15
昆明医科大学学报(2022年1期)2022-02-28
特种经济动植物(2022年2期)2022-02-15
当代水产(2021年1期)2021-03-19
四川蚕业(2021年1期)2021-02-12
电子技术与软件工程(2016年20期)2016-12-21
少儿科学周刊·少年版(2015年11期)2015-12-17
少儿科学周刊·少年版(2015年11期)2015-12-17
少儿科学周刊·少年版(2015年11期)2015-12-17
