
生物炭、磷及AMF 对Cd 胁迫下柳枝稷生长及土壤性质的影响
2021-12-17 01:26:52孙红郑玉龙林炎丽陈超杨富裕
草业学报 2021年12期
孙红 ,郑玉龙 ,林炎丽 ,陈超 ,杨富裕 *
(1. 贵州大学动物科学学院,贵州贵阳550000;2. 中国农业大学草业科学与技术学院,北京100089)
镉(Cd)作为生物毒性最强的主要重金属污染物,在我国污染土地面积已达2×107hm2[1-2]。Cd 对植物具严重的毒害作用:即抑制根系伸长生长、破坏叶绿素结构、抑制光合作用酶活性、引发氧化损伤、干扰营养元素吸收和运转等[3-4]。Cd 通过食物链进入人体后,损害肝脏、肾脏等器官,已在全球范围内引起诸多食品安全和人类健康问题[5]。因此,如何修复Cd 污染土壤已成为一个亟待解决的关键科学问题。
生物炭是各种有机物(家畜粪便、作物秸秆等)在厌氧条件下通过热解、气化和碳化过程获得的副产物[6],具有多孔、低密度、吸收面积大等特点[7],其作为肥料添加到土壤中可提高土壤中碳(有机碳)、氮、磷含量,同时改善土壤的酸碱度、孔隙度和持水能力等理化特性[8-9]。在重金属污染土壤中添加生物炭,可降低重金属的生物有效性,减少植物对土壤中重金属的吸收和转运[10-11]。土壤中的磷(phosphorus,P)由于化学沉淀和吸附作用易被土壤固定,生物活性低;研究发现添加生物炭可提高土壤中P 的利用效率,减少农业生产中无机磷肥的施用量[12-13]。许多研究也将生物炭和无机磷作为土壤改良剂固定其中的重金属离子,减少对植物的毒害作用[14-17]。
AMF 是与陆地上大多数植物形成共生关系的一类真菌,通过多种方式缓解重金属对植物的毒害作用。一方面AMF 可提高植物养分吸收(特别是P 的吸收),促进其生长,稀释重金属在植物体内的浓度[18-19];另一方面AMF 可将重金属聚集在其丛枝结构、根外菌丝及孢子中[20-21],或以多聚磷酸盐的形式将重金属螯合在液泡内[22-23]。因此,AMF 常作为一种生物肥料应用于重金属污染土壤的修复中。但AMF 与植物在重金属胁迫下的共生协同作用仅适于一定重金属含量范围内,超过一定阈值,植物提供的碳源不能满足AMF 生长所需,菌根真菌开始与植物竞争光和产物,导致接种AMF 后对植物生长的促进作用不明显[19];前期研究也发现土壤中Cd 含量超过20 mg·kg-1时,接种 AMF 对能源草柳枝稷(Panicum virgatum)生物量无显著促进作用[24]。
磷和生物炭的添加会影响AMF 与植物的亲和性。低浓度磷有利于AMF 对植物根系的侵染,过高浓度则会抑制[25];适度添加生物炭也有利于AMF 对植物根系的侵染和孢子的产生[26]。在重金属污染土壤中,AMF 结合生物炭可改良土壤理化性质,降低土壤中重金属的生物有效性,影响土壤微生物活性及丰度,降低重金属对植物的直接伤害[27-28]。已有研究报道了生物炭、磷及AMF 互作对自然土壤、Cd 污染土壤中植物的生长、Cd 吸收特性及土壤性质的影响[15-16]。
目前,还未见相关报道涉及高浓度Cd 胁迫下,AMF 和生物炭、磷互作对柳枝稷生长,矿质元素吸收及土壤性质的影响。因此,本研究以前期试验中对柳枝稷生长抑制作用较强的土壤Cd 含量(20 mg·kg-1)为基础,研究AMF、生物炭、磷对柳枝稷生物量,P、Se、Cd 吸收以及土壤Cd 形态的作用,以期能够找到提高柳枝稷修复重度Cd 污染土壤的方法。
1 材料与方法
1.1 试验材料及生长基质
本试验所用土壤采自于中国农业大学上庄试验站,土壤类型为壤土,风干后过2 mm 筛。土壤化学性质为:pH:8.27,速效氮:50.56 mg·kg-1,速效磷:15 mg·kg-1,速效钾:118.96 mg·kg-1,土壤有机质:1.13%,总 Cd 含量:0.27 mg·kg-1。细沙直径小于0.5 mm,自来水清洗干净后,用去离子水清洗3 次。按比例将土和沙子均匀混合(2∶1,w/w),经伽马射线辐射灭菌(25 kGy,10 MeVelecton beam)。Cd 处理是在沙土混合物中加入 20 mg·kg-1的 Cd(以 CdCl2·2.5H2O 形式),稳定 50 d。本试验 AMF 原始菌种(Rhizophagus irregularis,BGC AH01)由中国科学院生态环境研究中心提供。经过扩繁后接种物包括孢子、菌丝和植物根系的混合物,1 g 混合菌剂包含200个孢子。
1.2 试验设计
设置 1 个 Cd 浓度水平(20 mg·kg-1);2 个接菌处理:未接种 AMF(non-inoculation AMF,NM),接种 AMF(inoculation AMF,AM);4 个外源处理:对照(CK)、生物炭(B)、磷(P60)、磷和生物炭混合(B+P60)。每个处理设置 3 个重复,共 24 盆。其中,CK 处理是添加 30 mg·kg-1磷(以 KH2PO4的形式);B 处理是添加 30 mg·kg-1磷和4.5%的生物炭(w%),生物炭是由竹粉和猪粪混合高温厌氧制成,由浙江省农业科学院土壤肥料所提供;磷处理是添加60 mg·kg-1的磷(P60),磷和生物炭混合处理是分别添加60 mg·kg-1磷和4.5%生物炭。各处理均是先将生物炭、磷添加到土壤中混合均匀,之后再进行接菌与植物移栽。将2.3 kg 土壤基质放入用次氯酸钠消毒的花盆中(直径19 cm×高17 cm),达到2/3 时,将25 g 菌剂均匀地铺在上面,覆上剩余土壤。未接菌处理加入的菌剂在121 ℃烘箱中灭菌4 h。柳枝稷(Alamo)种子由中国农业大学动物科学技术学院育种试验室提供。用10% H2O2将种子消毒20 min,自来水冲洗干净后用去离子水冲洗3 遍,放在干净培养皿中发芽至4 cm。将10 株幼苗移栽到土壤基质中,生长10 d 后间苗为5 株;为保持接菌与未接菌处理具有相同的土壤细菌,每个未接种处理中加入5 mL 过0.425 mm 筛网的菌剂滤液。每天用去离子水浇水,保持花盆土壤15%的含水量(55%的水保持能力)。该试验于2017 年4-6 月在中国农业大学转基因基地温室中进行,在自然光条件下,温室条件控制在16 h/30 ℃,8 h/22 ℃。
1.3 样品采集及测定方法
移栽1 周后,每周各处理随机选取3 株测定植株高度,共测定7 周。生长60 d 后用SPAD 502 叶绿素仪(SPAD-502PLUS,中国),自上往下取第3 片叶测定叶绿素相对含量(SPAD 值)。60 d 后将地上部和根系分开收获,根系先用自来水冲洗干净,再用去离子水冲洗3 次。新鲜根系分为两部分,一部分用于测定菌根侵染率,一部分烘干测定干重(在65 ℃下烘48 h)后粉碎,用于测定Cd,P,Se 含量。分别称取0.3 g 干物质添加HNO3和H2O2(3∶1),利用微波消解仪(MarsX CEM,美国)经3 个加热步骤消化制成消煮样品;之后利用电感耦合等离子体质谱法(ICP-MS)(model 7500,安捷伦科技有限公司,美国)测定消煮样品中Cd、P、Se 含量。
采用2∶1 水土比(m∶v)测定土壤pH,采用Olsen 法测定土壤速效磷含量[29],采用对硝基苯磷酸二钠比色法测定土壤酸性磷酸酶活性[30]。利用改进后的BCR 连续提取法提取土壤中不同形态Cd[31],用ICP-MS 测定Cd 含量。
1.4 数据处理
采用SPSS 17.0 软件的three-ways ANOVA 分析生物炭、磷、AMF 及各处理互作对柳枝稷根系侵染率、SPAD、生物量、Cd、P、Se 含量及土壤pH 值、速效磷、土壤酸性磷酸酶活性和土壤不同Cd 形态含量的影响,如果不同处理在0.05 水平影响显著,之后进行LSD 多重比较。
2 结果与分析
2.1 生长速率
接种AMF 两周后各处理植株高度均高于未接菌处理(图1)。4 个处理植株高度在接菌和未接菌处理下的变化趋势基本一致,未接种AMF 植株高度变化趋势为:CK>P60和 B+P60>B;接种 AMF 变化趋势为:CK>P60>B+P60和 B。虽然接种 AMF 能显著提高柳枝稷植株高度,但接种AMF 后柳枝稷高度在CK与B、P60及B+P60处理间无显著差异。
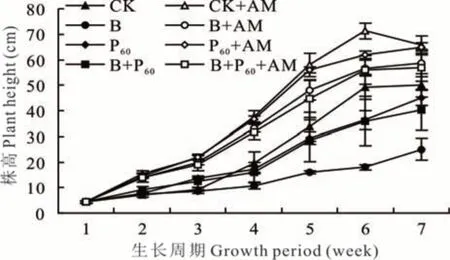
图1 不同处理下柳枝稷7 周内的植株高度Fig. 1 The plant height of switchgrass under different treatment for 7 weeks(mean±SD,n=3)
2.2 侵染率、SPAD 和生物量
磷添加显著影响柳枝稷根系侵染率(P<0.05),生物炭及与磷交互作用对菌根侵染率无显著影响(P>0.05)。接种AMF 显著影响柳枝稷叶片SPAD 值、地上部和根系生物量(P<0.05,P<0.001),生物炭添加显著影响根系生物量(P<0.05),磷添加对上述指标无显著影响(P>0.05)(表1)。
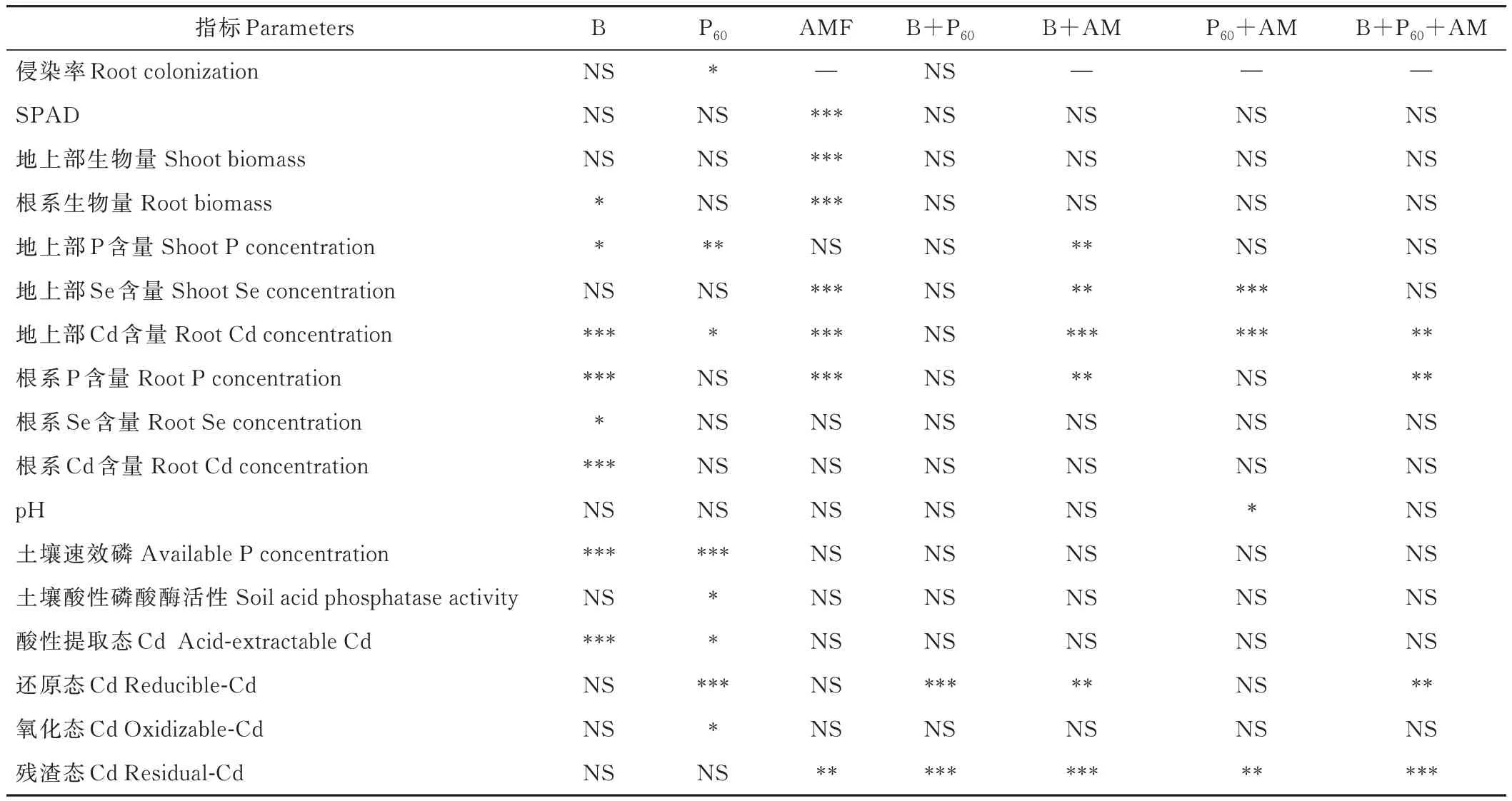
表1 生物炭、磷及AMF 处理对测定指标的影响Table 1 Effect of biochar,phosphorus addition and AMF inoculation on parameters tested
P60处理根系侵染率达到56.9%,显著高于CK,但与B 和B+P60处理差异不显著。与CK 相比,B、P60和B+P60处理叶片SPAD、地上部和根系生物量无显著变化,接种AMF 后各处理SPAD、地上部和根系生物量均显著升高。CK+AM、B+AM、P60+AM 和B+P60+AM 处理下SPAD 比对应未接菌处理分别提高了14%、24%、7%和15%,其中B+AM 处理SPAD 显著高于其他接菌处理;接种AMF 地上部生物量比对应未接菌处理分别提高了109%、1410%、214%和84%,根系生物量则分别提高了121%、608%、242%和129%,但接菌后各处理间生物量无显著差异(表2)。

表2 不同处理下柳枝稷侵染率、SPAD 值和生物量Table 2 The root colonization,SPAD value and biomass under biochar,phosphorus addition and AMF inoculation treatments(mean±SD,n=3)
2.3 P、Se 和 Cd 含量
生物炭、磷及生物炭与AMF 互作显著影响柳枝稷地上部P 含量(P<0.05,P<0.01);生物炭和磷显著影响地上部 Cd 含量(P<0.001,P<0.05),AMF 和 AMF 与磷、生物炭两两互作显著影响地上部 Se、Cd 含量(P<0.01,P<0.001)。生物炭添加显著影响根系P、Se、Cd 含量,AMF、AMF 与生物炭及与生物炭和磷三者互作也显著影响根系P 含量(P<0.01,P<0.001)(表1)。
与CK 相比,除了根系B+AM、地上部和根系B+P60+AM 的P 含量显著升高外,其他处理无显著变化。但与未接种处理相比,接种AMF 显著提高了B、P60处理地上部和根系P 含量。B、B+P60和P60+AM 地上部Se 含量与CK 无显著差异,接种AMF 后CK、B 和B+P60处理地上部Se 含量显著高于未接菌处理,分别提高了77%、280%和55%;与CK 相比,B+P60+AM 处理根系Se 含量显著升高,而其他处理无显著变化。地上部Cd 含量除了B 处理与CK 无显著差异外,其他处理均显著高于CK;接种AMF 后CK、B 和B+P60处理地上部Cd 含量高于未接菌处理,分别提高了116%、349%和13%。此外,B 和B+P60根系Cd 含量显著高于CK 和P60处理,但各处理根系Se 和Cd 含量在接菌与未接菌处理间无显著差异(图2)。
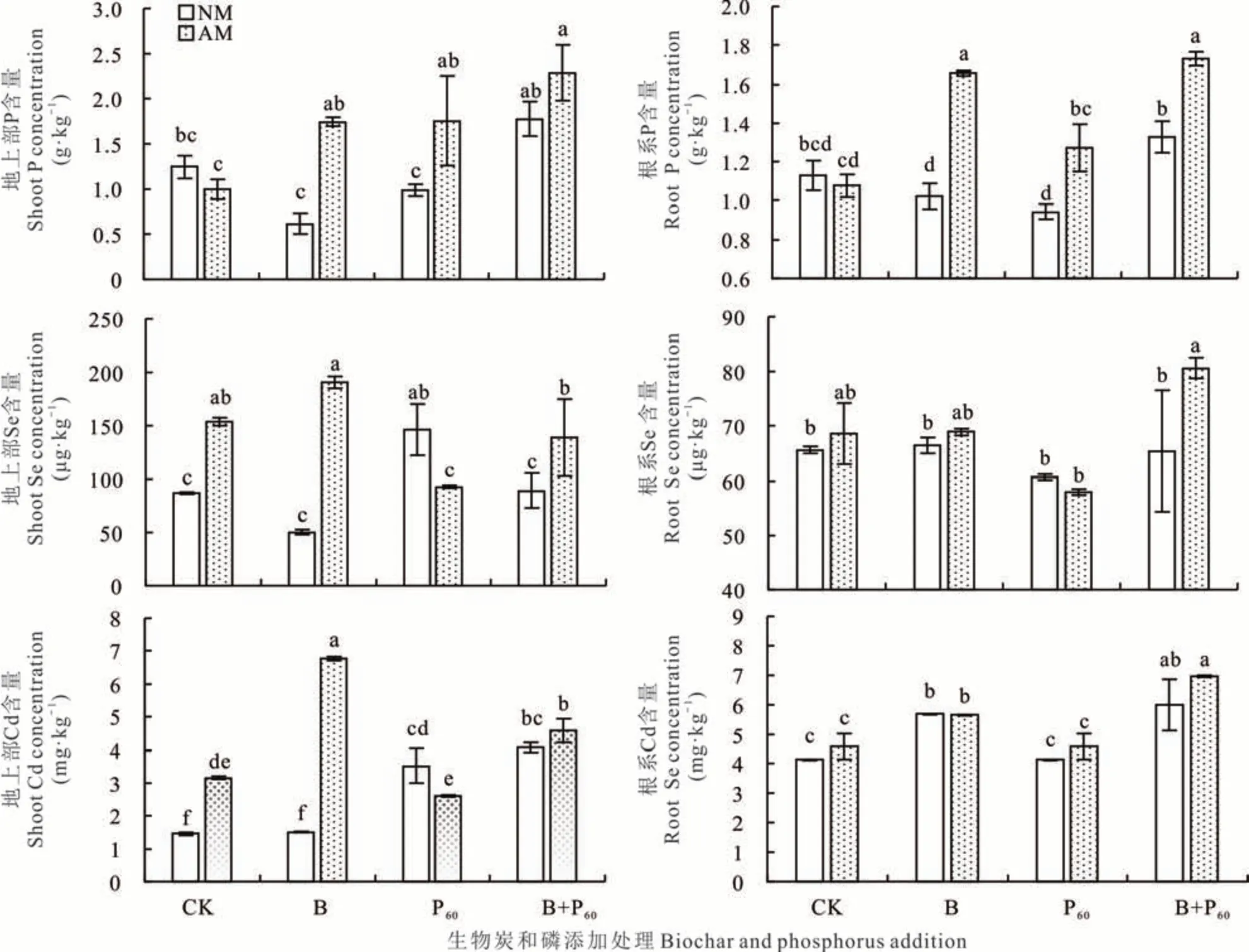
图2 不同处理下柳枝稷地上部和根系P,Se 和Cd 含量Fig. 2 P,Se and Cd concentration of switchgrass under biochar and phosphorus treatment,regardless of AMF inoculation(mean±SD,n=3)
2.4 土壤pH、速效磷和酸性磷酸酶活性
多因素方差分析表明:磷和AMF 交互显著影响土壤pH 值(P<0.05),生物炭添加显著影响土壤速效磷含量(P<0.001),磷添加显著影响土壤速效磷和磷酸酶活性(P<0.05)(表1)。无论是否接种AMF,各处理土壤pH值间无显著差异。不同处理下土壤速效P 含量趋势为:B+P60>P60>B>CK,除了AM+P60处理的速效P 含量显著低于未接菌处理(18%)外,其他处理在接菌与未接菌间无显著差异。与CK 相比,只有B+P60处理土壤酸性磷酸酶活性显著升高,其他处理无论是否接种AMF 均无显著变化。AMF+CK 处理下的土壤酸性磷酸酶活性比未接菌处理升高了110%,AM+B+P60处理比未接菌处理显著降低了51%(图3)。
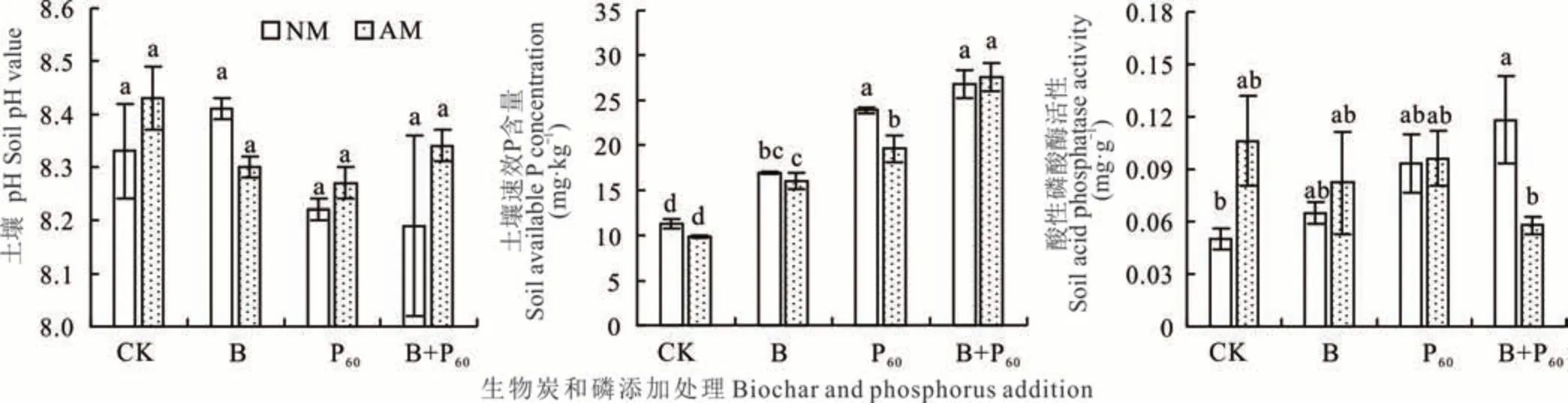
图3 不同处理下土壤pH,速效磷含量和酸性磷酸酶活性Fig. 3 The soil pH,available phosphorus concentration and acid phosphatase activity under different treatment,regardless of AMF inoculation(mean±SD,n=3)
2.5 土壤不同Cd 形态
生物炭和磷添加显著影响酸性提取态Cd 含量(P<0.001,P<0.05);生物炭和磷、AMF 两者交互及三者交互显著影响还原态Cd 含量(P<0.01,P<0.001);磷添加显著影响氧化态Cd 含量(P<0.05);除生物炭和磷单独作用外,AMF 及不同处理间的交互作用均显著影响残渣态Cd 含量(P<0.01,P<0.001)(表1)。无论是否接种AMF,各处理中酸性提取态和还原态Cd 含量变化趋势相似:其中酸性提取态Cd 含量变化趋势为:CK>P60>B和 B+P60处理;还原态 Cd 含量变化趋势为:P60、B 和 B+P60>CK;无论是否接菌,P60处理土壤中氧化态 Cd 含量保持在0.07 mg·kg-1,略高于其他处理。另外,接种AMF 显著提高了CK、B、B+P60处理下的土壤中残渣态Cd 含量,相比于未接菌处理分别升高了59%、53%、88%(表3)。
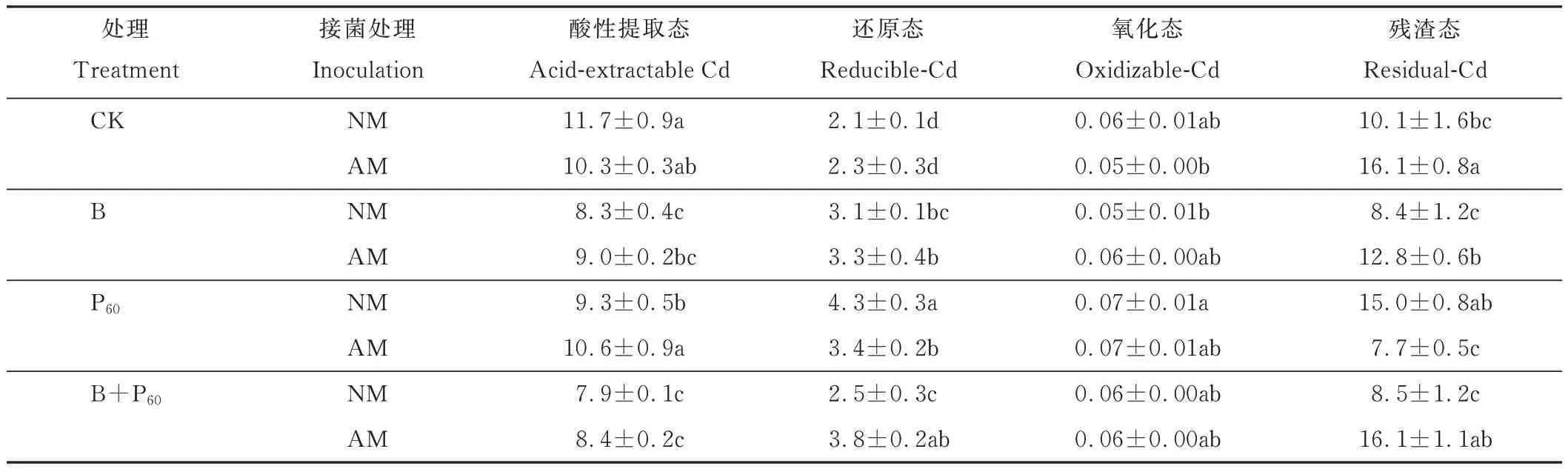
表3 不同处理下土壤酸性提取态、还原态、氧化态和残渣态Cd 含量Table 3 The concentrations of acid-extractable Cd,reducible-Cd,oxidizable-Cd and residual-Cd in the soil under different treatments(mg·kg-1,mean±SD,n= 3)
3 讨论
3.1 柳枝稷生长及养分吸收
生物炭是由有机物通过高温裂解而制成,保留了原料中的大量和微量元素,因此,施加生物炭能够提高土壤肥力而促进植物生长。Liu 等[15]和Xiao 等[16]研究发现单独添加生物炭或磷显著提高了Cd 胁迫下白三叶(Trifolium repens)、紫花苜蓿(Medicago sativa)的生物量,但本研究发现添加生物炭或磷并没有促进柳枝稷的生长(表 2),可能是由于本研究中土壤 Cd 浓度为 20 mg·kg-1,远高于之前报道的 Cd 浓度(1 mg·kg-1),从而导致生物炭和磷对柳枝稷的生长促进作用并不显著。此外,本研究结果发现接种AMF 后无论是否添加生物炭或磷,柳枝稷地上部和根系生物量均显著高于未接菌处理(表2)。大量研究已经证实AMF 与植物共生后一方面通过螯合物将重金属螯合在细胞质或将其区隔在液泡、丛枝、菌丝结构中,降低重金属对植物的直接伤害[20-22],另一方面可提高植物根系对养分的吸收效率(特别是磷),促进植物生长,通过“稀释效应”降低植物体内重金属浓度[23-24]。本试验也发现接种AMF 后所有处理(除了CK)的P 含量均显著高于未接菌处理(图2)。
Se 是植物必需的矿质元素之一,也是植物抗氧化系统中谷胱甘肽过氧化物酶(glutathione peroxidase,GSH)和其他硒蛋白的基本组成成分,在逆境条件下添加Se 可以促进植物体内形成GSH,提高植物抗氧化能力[25]。本试验结果发现添加生物炭可显著影响地上部Se 含量,表明生物炭中可能含有较高浓度的Se。此外,接种AMF后CK、B 和B+P60处理柳枝稷地上部Se 含量显著高于未接菌处理,AM+P60表现出相反趋势;同时各处理地上部Cd 含量变化与Se 相似,可能是因为接种AMF 后Se2+和Cd2+利用相同的离子载体进入植物体内,Se2+随Cd2+的吸收和转移可能也促进相应部位GSH 的合成,提高柳枝稷抗氧化能力。Durán 等[26]发现在干旱胁迫下莴苣(Lactuca sativa)接种AMF 后Se 含量与未接菌处理无显著差异,可能与植物受胁迫程度及AMF 种类有关。Cd胁迫下,生物炭、AMF 与植物体内Se 吸收间的关系,是否会直接影响柳枝稷体内GSH 的活性,还有待于进一步研究。
3.2 Cd 含量、土壤特性及土壤Cd 形态
生物炭具有吸附表面积大和离子交换能力强和显碱性的特点。之前研究发现添加生物炭能够提高土壤pH,降低土壤Cd 活性[15-16,27]。但本研究发现添加生物炭对土壤pH 无显著影响(图3),可能是因为本试验采用的土壤基质pH 为8.27,而之前报道的土壤pH 值低于7.5,表明生物炭对偏碱性土壤pH 值的影响可能弱于酸性或中性土壤。此外,本研究还发现添加生物炭后柳枝稷根系Cd 含量显著高于CK(图2),而之前报道发现添加生物炭可以降低植物地上部和根系Cd 含量[15-16,27],一方面可能是因为本试验中设置的Cd 浓度为20 mg·kg-1,远高于其他报道的Cd 浓度(1、5、10 mg·kg-1),虽然生物炭对重金属有一定的富集能力,降低了土壤中酸性提取态、还原态和残渣态Cd 含量(表3),但在生物炭添加质量分数类似(3.0%、4.5%、5.0%)的条件下,其对Cd2+的吸附力有限,可能会导致土壤中剩余较多的Cd 进入柳枝稷根系蓄积,另一方面,制作生物炭采用的热解温度不同会影响有机官能团和CO32-比例,导致生物炭对重金属吸收量产生差异[28]。
土壤中施加磷可以将土壤中活性态的Cd 转化为磷酸盐形态,降低Cd 的生物活性,进而减少植物中Cd 含量[32];本试验发现P60处理下地上部Cd 含量高于CK,根系Cd 含量与CK 无显著差异;土壤速效磷和残渣态Cd 含量显著升高,表明施加磷可降低Cd2+的生物活性。接种AMF 也可以改变重金属在植物组织中的吸收:接种AMF 后玉米(Zea mays)、柳枝稷地上部 Cd 含量降低[24,27],而百脉根(Lotus corniculatus)、龙葵(Solanum nigrum)地上部和根系Cd 含量升高[19,33],本试验发现接种AMF 对不同处理地上部Cd 含量产生了不同效应:AMF+P60处理柳枝稷地上部Cd 含量低于未接菌处理,其他接菌处理均高于未接菌处理,可能是因为磷添加处理显著影响了AMF 的侵染率,进而影响了菌根结构对 Cd 的螯合固定作用[20,34-35]。
土壤中施加生物炭可以增加酸性和中性土壤中的速效磷含量,提高土壤磷的可利用性[15-16,36],本研究发现无论是否接种AMF,生物炭均可提高偏碱性土壤中速效磷含量(图3),可能是因为生物炭自身磷含量及活性较高。土壤磷酸酶活性能够在一定程度上表示土壤中P 的转化效率及其对植物的有效性,缺磷环境下植物根系可直接分泌产生。接种AMF 后CK 处理的土壤酸性磷酸酶活性略高于未接菌处理,这与前期的研究结果相似[37-38]。AMF 自身也能产生一定量的酸性磷酸酶,提高土壤中酸性磷酸酶活性,促进植物对P 的吸收[39]。土壤磷酸酶活性只有在土壤P 含量不满足植物生长时才会升高,如果P 含量过高会抑制植物丝氨酸蛋白磷酸酶基因的表达,进而降低土壤中磷酸酶活性[40]。磷酸酶活性与土壤磷含量紧密相关。AMF 结合B+P60处理的酸性磷酸酶活性反而比未接菌处理降低50%,其他处理下接种AMF 对土壤磷酸酶活性未产生显著影响,推测可能是该处理下土壤P 含量和速效P 含量高于其他处理(图3)。因此,在接种AMF 后应该根据土壤磷含量适度添加生物炭或磷,以进一步提高土壤P 的利用率。
重金属的生物毒性受土壤重金属总量和金属种类的调控,重金属种类影响金属的生物可利用性。本试验土壤中Cd 的主要存在形式为酸性提取态和残渣态,与Chen 等[41]报道的结果相似。外源物质添加也会影响土壤中Cd 的形态及生物活性,本研究中无论是否接菌,磷、生物炭及二者混合处理下酸性提取态Cd 含量均显著低于对照,而相应还原态Cd 含量略高于对照,表明生物炭和磷的添加有助于土壤中酸性提取态Cd 向还原态转化。同时,AMF 也显著影响土壤中残渣态Cd 含量(表1),接种AMF 后CK、B 和B+P60处理下残渣态Cd 含量显著高于未接菌处理,表明AMF 能够降低土壤中重金属的生物活性。前期研究发现AMF 显著影响土壤酸性提取态和还原态Cd 含量[24],而本试验结果发现AMF 显著影响残渣态Cd 含量,对其他形式的Cd 无显著影响,可能是由于土壤Cd 浓度、土壤质地及柳枝稷品种的不同影响了其与AMF 的亲和性,导致重金属形态转化产生差异。
4 结论
当土壤Cd 浓度为20 mg·kg-1时,单独添加生物炭和磷对柳枝稷生物量无显著影响,但接种AMF 后各处理生物量均显著提高。单独添加P60和B+P60及接种AM 时地上部Cd 含量均显著升高,B 和B+P60处理根系Cd 含量显著升高。生物炭、磷添加可显著提高土壤速效磷含量。生物炭添加可显著降低土壤酸性提取态Cd 含量,磷添加可显著提高土壤还原性Cd 含量,接种AMF 显著提高CK、B 和B+P60土壤残渣态Cd 含量,但降低了P60残渣态Cd 含量。综上,AMF 结合生物炭和磷显著提高柳枝稷生物量、Cd 含量及土壤残渣态Cd 含量,促进了柳枝稷对Cd 的提取及土壤对Cd 的固定,可以在重度污染的Cd 土壤中应用。
猜你喜欢
作文大王·低年级(2023年2期)2023-02-10 13:20:59
小学生必读(低年级版)(2021年4期)2021-07-28 06:14:34
今日农业(2020年19期)2020-12-14 14:16:52
文苑(2020年2期)2020-11-22 01:45:32
文苑·经典美文(2020年2期)2020-04-19 10:00:34
中国组织化学与细胞化学杂志(2017年1期)2017-06-15 20:27:45
广西林业科学(2016年1期)2016-03-20 05:33:00
中国科技信息(2015年2期)2015-11-16 08:18:32
医学研究杂志(2015年8期)2015-06-22 14:00:57
植物营养与肥料学报(2014年1期)2014-03-11 20:24:38
