
2型糖尿病对小鼠下颌骨骨再生及辅助性T细胞17、调节性T细胞相关因子表达的影响
2021-12-16 13:39王亚楠吴旋贾婷婷冯瑶刘世岳徐欣张东姣
华西口腔医学杂志 2021年6期
王亚楠 吴旋 贾婷婷 冯瑶 刘世岳 徐欣 张东姣
1.山东大学齐鲁医学院口腔医学院·口腔医院种植科 山东省口腔组织再生重点实验室山东省口腔生物材料与组织再生工程实验室,济南250012;2.宁波口腔医院月湖分院种植科,宁波315000;3.山东大学齐鲁医学院口腔医学院·口腔医院牙周科 山东省口腔组织再生重点实验室山东省口腔生物材料与组织再生工程实验室,济南250012
肿瘤、感染、创伤和发育畸形等常导致不同程度的颌面部骨组织缺损,如何实现高效地颌骨骨再生一直是口腔医生面临的难题。多种系统性因素和局部因素可以影响颌骨再生,其中2型糖尿病(type 2 diabetes mellitus,T2DM) 是影响颌骨骨再生的危险因素。研究[1-3]表明,T2DM 可导致骨质疏松、骨折风险的增加和骨折愈合时间的延迟。然而,T2DM对颌骨骨缺损愈合的影响及潜在的分子机制尚未被完全阐明。
T2DM引发骨代谢不良的原因主要包括胰岛素分泌不足,尿钙、磷、镁排出增多,促破骨细胞因子分泌增多,活性维生素D 减少,糖基化终产物增多等等。近年来,糖尿病及其相关并发症的免疫致病机制得到越来越多的关注。研究人员发现,CD4+T 细胞中存在一组具有相反调节功能的亚群,包括发挥促炎效应的辅助性T 细胞17 (T helper cell 17,Th17) 和发挥抑炎效应的调节性T细胞(regulatory T cell,Treg)。有研究[4]证实,患有T2DM 的人与正常人相比,Th17/Treg 细胞的外周血比率显著升高。维持Th17/Treg 细胞平衡可以很好地维持骨代谢稳态,相反,Th17/Treg 细胞的失衡则与骨组织破坏显著相关[5]。然而,Th17/Treg细胞平衡在T2DM 导致的下颌骨骨缺损愈合不良过程中的改变仍需进一步的研究。
蛋白酪氨酸磷酸酶非受体型2 (protein tyrosine phosphatase non-receptor type 2,PTPN2) 是蛋白酪氨酸磷酸酶家族的重要成员,又称作T细胞蛋白酪氨酸磷酸酶。研究表明,PTPN2 在调节糖代谢[6]、抑制系统性炎症[7]、调控骨代谢[8]方面发挥重要作用。PTPN2 可以抑制Th17 细胞向的分化,抑制白细胞介素(interleukin,IL)-17分泌,同时促进Treg 细胞向分化,促进IL-10 分泌[9],但PTPN2在调节T2DM 小鼠下颌骨骨缺损愈合过程中Th17/Treg 细胞平衡中的表达情况和调控作用机制尚未得到全面的解释。
因此,本文通过建立T2DM 小鼠下颌骨骨缺损模型,观察T2DM 对小鼠下颌骨骨缺损愈合的影响及骨缺损区域内Th17 细胞、Treg 细胞相关因子表达的改变,并对PTPN2 是否参与此过程的可能性做出初步的探索,以期为T2DM 导致颌骨再生不良的分子机制研究和颌面部骨修复治疗提供新的思路。
1 材料和方法
1.1 实验动物
36 只6 周龄C57BL/6J 雄性小鼠均购自山东大学实验动物中心,所有实验按照《实验动物护理与使用指南》 进行,并经山东大学口腔医学院动物伦理委员会批准(NO.20170302)。36 只小鼠随机平分为2 组:正常对照(NC 组,n=18) 和2 型糖尿病组(T2DM 组,n=18)。T2DM 组喂养高脂高糖饲料4 周后,腹腔注射链脲佐菌素(streptozotocin,STZ)(30 mg·Kg-1);NC 组喂养普通饲料4周后,腹腔注射同等剂量柠檬酸钠缓冲液。分别于小鼠下颌骨骨缺损建模当天(0 d) 和术后7、14、28 d 进行空腹血糖的检测,当小鼠空腹血糖高于11.1 mmol·L-1且稳定1 周认为T2DM 建模成功,可用于后续实验。18 只T2DM 组小鼠建模成功后,在小鼠全身麻醉状态下,使用慢速手机于小鼠下颌骨下缘咬肌嵴的远中制作贯穿颊舌向的1 mm×1.5 mm 的骨缺损,即构建小鼠下颌骨骨缺损模型。愈合7、14、28 d后取材,4%多聚甲醛固定,12%乙二胺四乙酸脱钙,梯度乙醇脱水,石蜡包埋后进行切片检测。
1.2 主要试剂
STZ 试剂(Sigma 公司,美国),Runt 相关转录因子2 (Runt-related transcription factor 2,RUNX2) 抗体、叉头框蛋白P3 (forkhead box protein P3,Foxp3) 抗体(Cell Signaling Technology公司,美国),碱性磷酸酶(alkaline phosphatase,ALP)抗体、维甲酸相关孤核受体(retinoic acid related orphan receptor,ROR) γt 抗体、蛋白酪氨酸磷酸酶非受体型2 (tyrosine-protein phosphatase non-receptor type 2,PTPN2) 抗体(Abcam公司,英国),苏木素-伊红(hematoxylin-eosin,HE) 染色试剂盒(北京索莱宝科技有限公司),SP-9000 免疫组织化学试剂盒、二氨基联苯胺(3,3’-diaminobenzidine,DAB) 显色试剂盒(北京中杉金桥生物技术有限公司)。
1.3 HE染色
NC 组和T2DM 组小鼠下颌骨缺损部位的石蜡切片经过二甲苯脱蜡、梯度乙醇水化后,苏木素染色2 min,流水冲洗7 min;伊红溶液染色4 min,流水冲洗3 min;梯度乙醇脱水,二甲苯透明后使用中性树胶封片。使用光学显微镜(Olympus 公司,日本) 于10 倍物镜下观察小鼠下颌骨缺损部位的新骨形成情况。
1.4 免疫组织化学染色
为了观察T2DM 小鼠下颌骨缺损愈合过程中Th17/Treg 细胞平衡的改变情况,并进一步地探讨可能的原因,本研究通过免疫组织化学染色的方法对NC 组和T2DM 组小鼠下颌骨缺损组织内Treg细胞分化依赖的特异性转录因子Foxp3、Th17 细胞分化所需的特异性转录因子RORγt 以及在Th17/Treg 细胞平衡调控过程中发挥重要作用的蛋白PTPN2的表达情况进行了检测。
NC 组和T2DM 组小鼠下颌骨缺损部位的石蜡切片经过二甲苯脱蜡、梯度乙醇水化,胰酶抗原修复后,使用山羊血清封闭剂封闭30 min,滴加ALP 抗体(1∶300)、 RUNX2 抗体(1∶250)、Foxp3 抗体(1∶200)、RORγt 抗体(1∶200) 或PTPN2 抗体(1∶300),4 ℃过夜。次日复温后,分别滴加生物素化二抗和酶结合物于37 ℃条件下孵育25 min,DAB 溶液显色20~30 s,苏木素溶液复染1 min,流水冲洗7 min。切片梯度乙醇脱水,二甲苯透明,中性树脂封片。ALP 和PTPN2 的免疫组织化学染色强度由单位面积内积分光密度值(integrated optical density/area, IOD/area) 衡量;RUNX2、Foxp3 和RORγt 的免疫组织化学染色强度由40倍物镜视野下的阳性细胞数衡量,使用Image-J软件软件进行定量分析。
1.5 统计分析
所有数据均以至少3 个独立实验的平均值±标准差表示,通过Shapiro-Wilk 检验评估数据是否符合正态分布,对于正态分布的数据,采用SPSS 20.0统计软件进行t检验比较两组之间的差异;对于非正态分布的数据,使用非参数检验,P值小于0.05被认为差异有统计学意义。
2 结果
2.1 空腹血糖、HE 染色、ALP 和RUNX2 的免疫组织化学染色检测结果
结果如表1 所示,T2DM 组小鼠在下颌骨骨缺损建模当天(0 d) 和术后7、14、28 d的空腹血糖平均值分别为21.6、 22.6、 23.1、 23.2 mmol·L-1,均明显高于对应时间点NC 组小鼠的空腹血糖和11.1 mmol·L-1的建模标准。

表1 各组小鼠空腹血糖水平Tab 1 The fasting blood glucose levels in each group mmol·L-1
HE 染色结果(图1) 显示:7 d 时,成纤维细胞在NC 组下颌骨缺损内排列整齐,骨缺损边缘出现少量骨基质沉积,而T2DM 组下颌骨缺损区的新生骨基质较少;14 d 时,NC 组缺损边缘出现连续条索状的新生骨组织,而T2DM 组缺损区内出现大量软骨细胞;28 d 时,NC 组缺损区新生骨组织成片状分布,而T2DM 组的缺损区仍存在大量结缔组织。

图1 NC组和T2DM组小鼠下颌骨缺损愈合7、14、28 d后的HE染色结果Fig 1 HE staining of the mandibular defect in NC group and T2DM group mice after 7 d, 14 d, 28 d
ALP 的免疫组织化学染色结果(图2) 显示:7 d 时,ALP 在NC 组新生骨基质周围呈阳性表达,在T2DM 组缺损区仅有少量的表达,且表达量明显低于NC 组;随着骨愈合时间的延长,NC 组新生骨组织周围仍显示出ALP 的阳性表达,其表达强度较7 d 时有所降低,但均明显强于T2DM 组。定量分析结果(图3) 显示:ALP 在T2DM 组与NC 组下颌骨缺损愈合7、14、28 d 后的表达明显降低,差异有统计学意义(P<0.05)。

图2 小鼠下颌骨缺损愈合7、14、28 d后ALP免疫组织化学镜下照片Fig 2 Pictures under microscope of ALP immunohistochemical staining in the mandibular defect of mice after 7 d, 14 d, 28 d

图3 小鼠下颌骨缺损愈合7、14、28 d 后ALP 免疫组织化学染色定量分析Fig 3 Quantitative analysis of ALP immunohistochemical staining in the mandibular defect of mice after 7 d, 14 d, 28 d
RUNX2 的免疫组织化学染色结果(图4) 显示:7 d 时,RUNX2 在NC 组新生骨周围呈现阳性表达,随着骨缺损愈合时间的延长,新生骨组织周围中RUNX2 的表达出现较7 d 时减弱的现象,RUNX2 在T2DM 组骨缺损区域的表达均明显低于NC 组。定量分析结果(图5) 显示:T2DM 组骨缺损区域的RUNX2阳性细胞数量明显低于NC组,差异有统计学意义(P<0.05)。
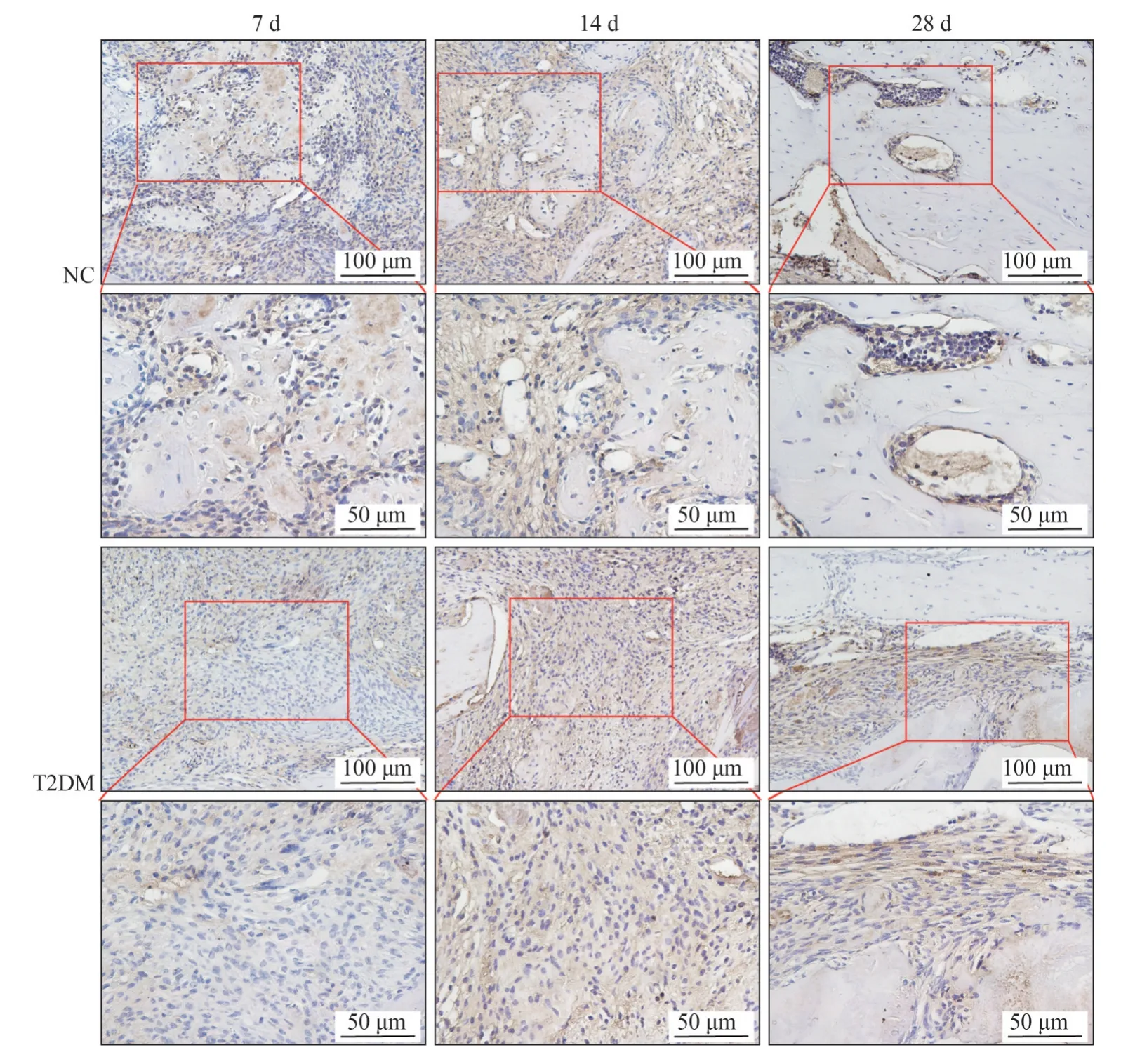
图4 小鼠下颌骨缺损愈合7、14、28 d后RUNX2免疫组织化学镜下照片Fig 4 Pictures under microscope of RUNX2 immunohistochemical staining in the mandibular defect of mice after 7 d, 14 d, 28 d

图5 小鼠下颌骨缺损愈合7、14、28 d 后RUNX2 免疫组织化学染色定量分析Fig 5 Quantitative analysis of RUNX2 immunohistochemical staining in the mandibular defect of mice after 7 d, 14 d,28 d
上述结果证实T2DM 显著抑制了小鼠下颌骨骨缺损的愈合过程。
2.2 Foxp3和RORγt的免疫组织化学染色检测结果
Foxp3的免疫组织化学染色结果(图6) 显示:7 d 时,Foxp3 在NC 组新生骨周围呈现阳性表达,随着骨缺损愈合时间的延长,新生骨组织周围中Foxp3的表达较7 d时减弱,Foxp3在T2DM组骨缺损区域的表达均明显低于NC组。定量分析结果(图7) 显示:T2DM组骨缺损区域的Foxp3阳性细胞数量明显低于NC组,差异有统计学意义(P<0.05)。

图7 小鼠下颌骨缺损愈合7、14、28 d 后Foxp3 免疫组织化学染色定量分析Fig 7 Quantitative analysis of Foxp3 immunohistochemical staining in the mandibular defect of mice after 7d, 14 d, 28 d
RORγt 的免疫组织化学染色结果(图8) 显示,7 d时,NC组新生骨基质周围存在少量RORγt阳性细胞,而T2DM 组缺损区的RORγt 阳性细胞明显增多;14 d 时,NC 组骨缺损区域内RORγt 阳性细胞数量较7 d 时有所降低,但明显少于T2DM组;28 d时,T2DM 组缺损区的RORγt阳性细胞数量较14 d时增多,且明显多于NC 组。定量分析结果(图9) 显示:T2DM 组骨缺损区域的RORγt 阳性细胞数量明显高于NC 组,差异有统计学意义(P<0.05)。

图8 小鼠下颌骨缺损愈合7、14、28 d后RORγt免疫组织化学镜下照片Fig 8 Pictures under microscope of RORγt immunohistochemical staining in the mandibular defect of mice after 7 d, 14 d, 28 d
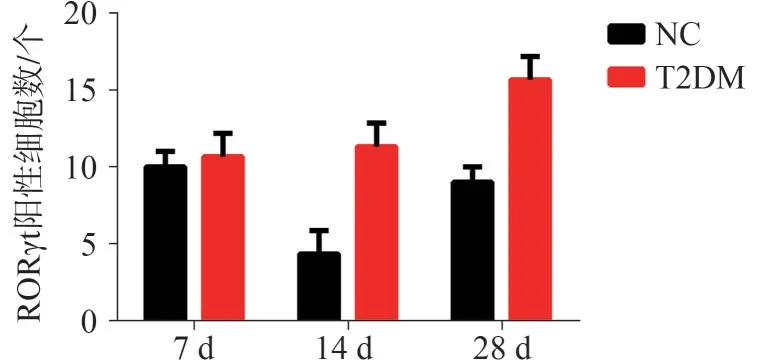
图9 小鼠下颌骨缺损愈合7、14、28 d 后RORγt 免疫组织化学染色定量分析Fig 9 Quantitative analysis of RORγt immunohistochemical staining in the mandibular defect of mice after 7 d, 14 d,28 d
上述结果证实T2DM 小鼠下颌骨缺损愈合过程中出现了Treg-Th17向偏移。
2.3 PTPN2的免疫组织化学染色检测结果
PTPN2 的免疫组织化学染色结果(图10) 显示:7 d 时,PTPN2 在NC 组新生骨周围呈现阳性表达,随着骨缺损愈合时间的延长,新生骨组织周围中PTPN2 的表达较7d 时减弱, PTPN2 在T2DM 组骨缺损区域的表达均明显低于NC 组。定量分析结果(图11) 显示:T2DM 组骨缺损区域内PTPN2表达量明显低于NC组,差异有统计学意义(P<0.05)。

图10 小鼠下颌骨缺损愈合7、14、28 d后PTPN2免疫组织化学镜下照片Fig 10 Pictures under microscope of PTPN2 immunohistochemical staining in the mandibular defect of mice after 7 d, 14 d, 28 d

图11 小鼠下颌骨缺损愈合7、14、28 d 后PTPN2 免疫组织化学染色定量分析Fig 11 Quantitative analysis of PTPN2 immunohistochemical staining in the mandibular defect of mice after 7 d, 14 d,28 d
上述结果发现,PTPN2 在T2DM 小鼠下颌骨缺损愈合过程中的表达明显降低,这提示PTPN2表达的降低可能是T2DM 小鼠下颌骨缺损愈合过程中出现Treg-Th17向偏移的原因。
3 讨论
T2DM是以高血糖为特征的代谢性疾病,具有很高的致残率和致死率,现已成为严重危害人类健康的全球性公共问题[10-11]。本研究联合应用高脂高糖饮食与低剂量STZ 注射的方法,成功构建T2DM 小鼠模型[12]。在整个实验过程中,T2DM 组小鼠的空腹血糖水平均稳定维持在11.1 mmol·L-1之上。HE 染色结果显示,T2DM 组小鼠的下颌骨骨缺损范围内新生骨的数量明显少于NC 组;免疫组织化学染色结果显示,成骨相关蛋白ALP 和RUNX2 的表达在T2DM 组的骨缺损内明显降低;上述结果证实了T2DM 可以显著地抑制下颌骨的骨再生过程。
近年来,Th17/Treg 免疫平衡机制在糖尿病及其并发症、炎症性骨代谢等多种生物学过程中的作用受到了越来越多的关注[5]。研究[13]表明,在T2DM 肾病患者中,尿白蛋白与肌酐的比值与Th1和Th17 细胞的比例呈正相关,与Treg 细胞的比例呈负相关。Samuel 等[14]研究证实,Th17 细胞在糖尿病大鼠根尖周炎的发生发展过程中发挥主导作用。 Th17 细胞分化所需的特异性转录因子是RORγt,Treg细胞的分化则依赖于特异性转录因子Foxp3[15], 因此Foxp3 与RORγt 的表达对“Treg-Th17 向偏移” 具有重要的调控作用。免疫组织化学染色的结果显示,T2DM小鼠下颌骨骨缺损范围内的RORγt阳性细胞数量增加,Foxp3阳性细胞数量减少,这提示T2DM 小鼠下颌骨骨缺损范围内可能出现了 “Treg-Th17向偏移”。
PTPN2 是蛋白酪氨酸磷酸酶家族的重要成员,包括TC45 和TC48 两种形式[16]。其中当细胞受到刺激后,TC45 游离出细胞核,去磷酸化细胞质内的某些关键蛋白质(如蛋白酪氨酸激酶、信号转导与转录激活因子等),从而抑制下游信号通路的转导,在调节糖代谢、抑制系统性炎症、调控骨代谢方面发挥重要作用[17]。在本实验中,免疫组织化学染色的结果显示T2DM 小鼠下颌骨骨缺损范围内的PTPN2 表达减少,这提示PTPN2 可能参与到T2DM 小鼠颌骨愈合 “Treg-Th17 向偏移” 的过程中,这与先前的研究发现较为一致。有报道证实,高血糖状态会显著降低全身组织或器官中PTPN2 的表达,T2DM 患者的血清中PTPN2 水平显著低于健康成年人。Li 等[18]通过免疫组织化学染色和蛋白质印迹法这两种实验手段检测到与正常组相比,糖尿病组小鼠附睾白色脂肪组织中PTPN2 蛋白水平显着降低。本课题组的前期实验研究[8]发现,T2DM 大鼠股骨种植体周围PTPN2 的表达明显降低;另外,经高糖培养基培养的大鼠骨髓间充质干细胞中PTPN2 的表达也与体内实验呈现相同的趋势。
PTPN2 是调控糖代谢的重要分子,已有研究表明,肝脏特异性PTPN2 敲除小鼠的肝脏,脂肪变性、肥胖和胰岛素抵抗情况明显加重;胰腺特异性PTPN2 敲除小鼠经长期高脂肪饮食喂养后,表现出葡萄糖耐量降低和葡萄糖刺激的胰岛素分泌减少。另外,PTPN2 可以调节机体的固有免疫和适应性免疫,有效地抑制系统性炎症。Li 等[18]证实,过表达PTPN2 可以逆转糖尿病小鼠附睾白色脂肪组织中的高Th17/Treg 比例。有研究[9]发现,在结肠炎小鼠动物模型中进行Ptpn2-/-CD4+T 细胞再输注,使得Th1细胞数量增加了近3倍,Th17细胞数量增加了2 倍,Treg 细胞数量减少了3 倍。除此之外,PTPN2 在调控骨代谢、降低骨缺损的局部破骨细胞密度等方面具有一定的积极作用。最近的研究表明,PTPN2 缺乏导致小鼠自发性滑膜炎和软骨下的骨吸收,股骨中骨量减少以及破骨细胞密度增加。因此,通过靶向调控PTPN2,实现对糖尿病性骨代谢不良的治疗,具有极大的应用前景,这也是后续进一步的研究方向。
综上所述,本研究结果表明T2DM 可以显著地抑制下颌骨骨再生过程,PTPN2 的下降以及Treg-Th17向偏移可能为其潜在的分子机制。
利益冲突声明:作者声明本文无利益冲突。
猜你喜欢
中国现代医生(2022年23期)2022-09-21
江苏农业科学(2021年22期)2021-12-08
影像诊断与介入放射学(2021年5期)2021-11-29
口腔颌面外科杂志(2021年4期)2021-09-04
口腔颌面外科杂志(2021年3期)2021-07-15
昆明医科大学学报(2021年1期)2021-02-07
口腔医学(2020年8期)2020-09-08
野生动物学报(2020年1期)2020-02-21
浙江医学(2019年9期)2019-05-20
安徽农业科学(2015年7期)2015-07-13
