
甘薯IbWRKY44 的克隆与表达分析
2021-12-16 03:37赵彩良吕运韬董静静刘世芳贾小云
山西农业科学 2021年12期
赵彩良,常 璐,吕运韬,张 洁,董静静,刘世芳,贾小云
(1.山西农业大学生命科学学院,山西太谷 030801;2.山西农业大学农学院,山西太谷 030801;3.临沂市河东区汤泉高级中学,山东临沂 276027)
甘薯(Ipomoea batatas(L.)Lam)块根富含多种营养成分如维生素、蛋白质以及花青素、类胡萝卜素等,是重要的粮食作物。甘薯的种植具有操作简单、成本低、产量高等特点,其产量仅次于水稻、小麦和玉米[1]。
花青素属于类黄酮物质,是一种水溶性的可食用色素,存在于植物液泡中且在不同pH 条件下呈现粉色、红色、蓝色、紫色等颜色[2]。花青素不仅可以帮助植物抵抗不良的外界环境、吸引昆虫传播花粉,还在人类的医疗保健方面发挥着重要作用,如具有抗氧化、抗癌、预防心血管疾病等功能[3-5]。花青素的生物合成受到多种因素的调节,包括结构基因、调节基因、环境因子(光照、温度、糖、激素、pH、干旱、肥料、病害等)以及microRNA 等。结构基因是指编码花青素生物合成途径中的酶基因,包括CHS(查尔酮合成酶)、CHI(查尔酮异构酶)、F3H(黄烷酮-3-羟化酶)、DFR(二羟基黄酮醇还原酶)、ANS(花色素合成酶)和UFGT(类黄酮糖基转移酶)等。调节基因主要包括MYB、bHLH 和WD40 三类转录因子,它们通过形成MBW 复合体来参与调控花青素的生物合成。近年来也发现许多其他类型的转录因子参与了花青素生物合成的调控,包括NAC、WRKY、AP2、SPL等。杨慧珍[6]研究表明,IbANS 在紫色甘薯的块根、茎和叶中的表达量均显著高于白色甘薯,可能参与花青素的合成。CHU 等[7]研究表明,过表达IbMYB1 的转基因拟南芥中花青素的含量上升且花青素合成相关基因CHI、F3′H、DFR 的表达量均上调。DONG等[8]的研究发现,甘薯IbWD40 的表达量随花青素的积累而升高。目前有关甘薯花青素的研究主要集中在MBW 复合体的调控机制上,而关于甘薯WRKY 转录因子对花青素调控的研究鲜有报道。
WRKY 转录因子的N 端含有保守基序为WRKYGQK 的WRKY 保守结构域,C 端含有C2H2或C2HC 锌指结构[9-10]。WRKY 转录因子一般通过与基因启动子中的W-Box 作用元件特异结合而调控基因的转录,进而调控植物的生长发育过程[10]。根据结构域的数量和特征,WRKY 转录因子可分为GroupⅠ、GroupⅡ和GroupⅢ等3 类,Group I 有2 个结构域,锌指结构为C2H2;GroupⅡ和GroupⅢ均只有1 个结构域,锌指结构分别为C2H2和C2HC。根据WRKY 结构域外的其他保守结构基序,GroupⅢ成员可进一步分为IIa、IIb、IIc、IId 和IIe 等5 个亚类[9]。WRKY 转录因子在植物的生长发育、新陈代谢、胁迫响应及次生代谢中具有重要作用。拟南芥AtWRKY44 在毛状体形成、花青素积累、种子色素沉着、胚乳发育、种子大小和根部有无毛细胞分化等生长发育进程中均发挥作用[11-15]。HAN 等[16]研究发现,在干旱胁迫下,GI-miRNA172E-AtWRKY44调控拟南芥的糖信号通路从而加速植株开花。王熙然[17]在草莓中过表达FaWRKY44 后,果实中花青素含量显著上升,原花青素含量下降;而减低表达FaWRKY44 后,花青素含量显著下降,原花青素含量显著上升。CHENG 等[18]在红火龙果中研究发现,HpWRKY44 通过与HpCytP450-like1 启动子中的W-Box 元件结合而激活其表达,二者在果实着色过程中含量逐步增加。杨树等[19]研究发现,丹参SmTTG2(SmWRKY44)的表达量随着花和种子的发育逐渐升高,推测其参与丹参种子的发育及萌发过程。PENG等[20]研究发现,猕猴桃的AcWRKY44 和AcMYBC1在烟草叶片中瞬时过表达会促进烟草叶片积累花青素,但在猕猴桃愈伤组织中稳定过表达后会促使F3′5′H 和原花青素合成相关基因及原花青素含量升高,说明AcWRKY44 和AcMYBC1 在平衡花青素与原花青素的含量方面具有重要作用。烟草NtTTG2 通过调控NtARF8、NtARF17 和NtARF19进而影响烟草的生长发育[21]。目前尚未见到关于甘薯IbWRKY44 功能研究的相关报道。紫心甘薯块根富含花青素,是研究植物地下器官中花青素生物合成调控机制的理想材料。
本研究从紫心甘薯块根中克隆了IbWRKY44,通过生物信息学分析和表达特性分析,初步探究了IbWRKY44 在花青素生物合成中的作用,并且成功构建了融合表达载体,为深入解析甘薯IbWRKY44在甘薯块根中花青素合成调控机制的研究提供参考依据,对培育富含花青素的甘薯新品种具有重要的理论和指导意义。
1 材料和方法
1.1 试验材料
供试紫心甘薯有徐紫薯3 号(XZS-3)、徐紫薯8 号(XZS-8)和宁紫薯4 号(NZS-4),白心甘薯有徐薯18 号(XS-18)和薯绿1 号(SL-1),淡黄心品种高系14(GX-14),均由江苏省农业科学院徐州甘薯研究中心提供,种植于山西农业大学甘薯基地。本氏烟草(Nicotiana benthamiana Domin.)和Columbia型野生拟南芥(Arabidopsis thaliana)种子由山西农业大学生命科学学院分子生物学实验室保存,拟南芥T-DNA 插入突变体种子(SALK-206852C 和SALK-204777C)购自AraShare 网站(www.arashare.cn)。烟草和拟南芥植株种植于人工气候培养箱,培养条件为温度26 ℃,光照16 h(光照强度125 mmol/(m2·s)),黑暗8 h,湿度50%。
大肠杆菌菌株DH5α 和农杆菌菌株GV3101均由山西农业大学生命科学学院分子生物学实验室保存。
1.2 RNA 的提取与cDNA 的合成
取种植90 d 的XZS-3、XZS-8、NZS-4、XS-18、SL-1 和GX-14 的块根,液氮研磨后用RNAiso Plus试剂提取RNA,经琼脂糖凝胶电泳和核酸蛋白检测仪NanoDrop 2000C(Thermo Scientifc)检测RNA 的完整性和质量后,按照TAKARA 反转录试剂盒的说明书反转录生成cDNA。
1.3 IbWRKY44 基因的克隆
通过分析山西农业大学生命科学学院贾小云课题组白心甘薯和紫心甘薯块根的转录组数据,找到在紫心甘薯中高表达而在白心甘薯中低表达的序列(Tai6.25536.1),经序列同源性比对,与拟南芥AtWRKY44 的相似性最高,命名为IbWRKY44。设计含有KpnⅠ(GGTAC|C)和SalⅠ(G|TCGAC)酶切位点的克隆引物(IbWRKY44-PF-KpnⅠ:GGGGTACC TTGAAGATGGAGATCAAAGAG;IbWRKY44-PRSalⅠ:GCGTCGACCCATCTCGGTTTTTGTCTG-AT),以XZS-3 块根的cDNA 为模板进行PCR 扩增,扩增产物纯化回收后与克隆载体PMD19-T 相连,获得重组质粒PMD19-T-IbWRKY44,将其转入大肠杆菌DH5α,在含氨苄青霉素(Amp)的固体培养基上挑选克隆进行PCR 鉴定,提取阳性克隆的质粒DNA后送上海生物工程有限公司(中国,上海)测序。
1.4 IbWRKY44 的生物信息学分析
通过在线软件对IbWRKY44的氨基酸序列进行生物信息学分析,所用网址如表1 所示。

表1 IbWRKY44 序列分析所用网址及功能
1.5 IbWRKY44 的系统进化分析
下载PlantTFDB 中注释明确的拟南芥的69 个WRKY 家族成员的氨基酸序列,与IbWRKY44 的氨基酸序列利用MEGA 软件构建系统进化树。同时,将NCBI 中与IbWRKY44 氨基酸序列相似度较高的牵牛花、番茄、大豆等12 个物种的WRKYs 序列和IbWRKY44 利用DNAMAN 和MEGA 软件分别进行多序列比对和系统进化树的构建。
1.6 IbWRKY44 启动子的顺式作用元件分析
在甘薯基因组数据库(https://ipomoea-genome.org/)中比对获得IbWRKY44 基因的DNA 序列。下载IbWRKY44 基因起始密码子ATG 上游1 600 bp的序列,通过PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/search_CARE.html)进行启动子顺式作用元件分析。
1.7 IbWRKY44 的表达模式分析
根据IbWRKY44 的序列设计定量引物:qRTPCR-IbWRKY44-PF:CACCGTGCTATCCCGTTACA;qRT-PCR-IbWRKY44-PR:GCCTTCTGCCTCCAAT CCTT。以XZS-3、XZS-8、NZ-4、XS-18、SL-1 和GX-14 块根的cDNA 为模板,运用BIO-RAD CFX 96 荧光定量仪对IbWRKY44 基因进行表达模式分析,IbActin 为内参基因,每个样品设置3 个生物学重复。基因相对表达量的计算采用2-ΔΔCt法,运用SPSS Statistics v23 软件对数据进行差异显著性分析(P<0.05)。
1.8 IbWRKY44 融合表达载体的构建
将测序得到的IbWRKY44 序列去除终止密码子后设计带有SalⅠ酶切位点的后引物IbWRKY44-2PR-Sal Ⅰ:GCGTCGACTGATTTTTCCTTTGTGGC。以PMD19-T-IbWRKY44 质粒DNA 为模板进行PCR 扩增,对扩增产物和pCAMBIA1300-GFP 表达载体分别用KpnⅠ和SalⅠ双酶切,回收目的片段,T4 DNA 连接酶连接后转至大肠杆菌DH5α,挑选在添加卡那霉素(Kan)的LB 固体培养基上生长的克隆进行PCR 鉴定。提取阳性克隆的质粒DNA,用KpnⅠ和SalⅠ对质粒进行双酶切验证,获得35S::IbWRKY44::GFP 融合表达载体,4 ℃保存待用。
1.9 IbWRKY44 的烟草亚细胞定位
将空载体pCAMBIA1300-GFP 和构建好的35S::IbWRKY44::GFP 融合表达载体分别通过热激法转入农杆菌菌株GV3101 中,挑取在含有50 μg/mLKan 和50 μg/mL 利福平(Rif)的固体LB培养基上的克隆,经PCR 鉴定为阳性克隆后(35SF:GACGCACAATCCCACTATCC 和 IbWRKY44-2PR-SalⅠ),于50 mL 相应液体培养基中过夜培养至OD600为0.6,4 ℃,12 000 r/min 离心10 min,弃上清收集菌体,用含有10 mmol/L MgCl2、10 mmol/L 2-吗啉乙磺酸(MES)和100 μmol/L 乙酰丁香酮(AS)的悬浮液重悬菌体至OD600为0.2,室温孵育4 h 后注射长势一致的28 d 烟草叶片,培养箱培养2~3 d后撕取注射部位叶片,经1 μg/mL 的4′,6-二脒基-2-苯基吲哚(DAPI)染色5 min 后在Leica 荧光正置显微镜488 nm 的波长下观察绿色荧光。亚细胞定位的预测根据表1 所列网址进行。
2 结果与分析
2.1 IbWRKY44 基因的克隆
以XZS-3 块根cDNA 为模板获得的PCR 产物经琼脂糖凝胶电泳检测后条带单一,大小约为1 400 bp,纯化回收后与PMD19-T 克隆载体连接,得到的菌落PCR 检测条带也单一,且大小一致,约1 400 bp(图1)。阳性菌落提取的质粒DNA 经测序后,得到长度为1 401 bp 的IbWRKY44 基因的ORF序列。

2.2 IbWRKY44 序列的生物信息学分析
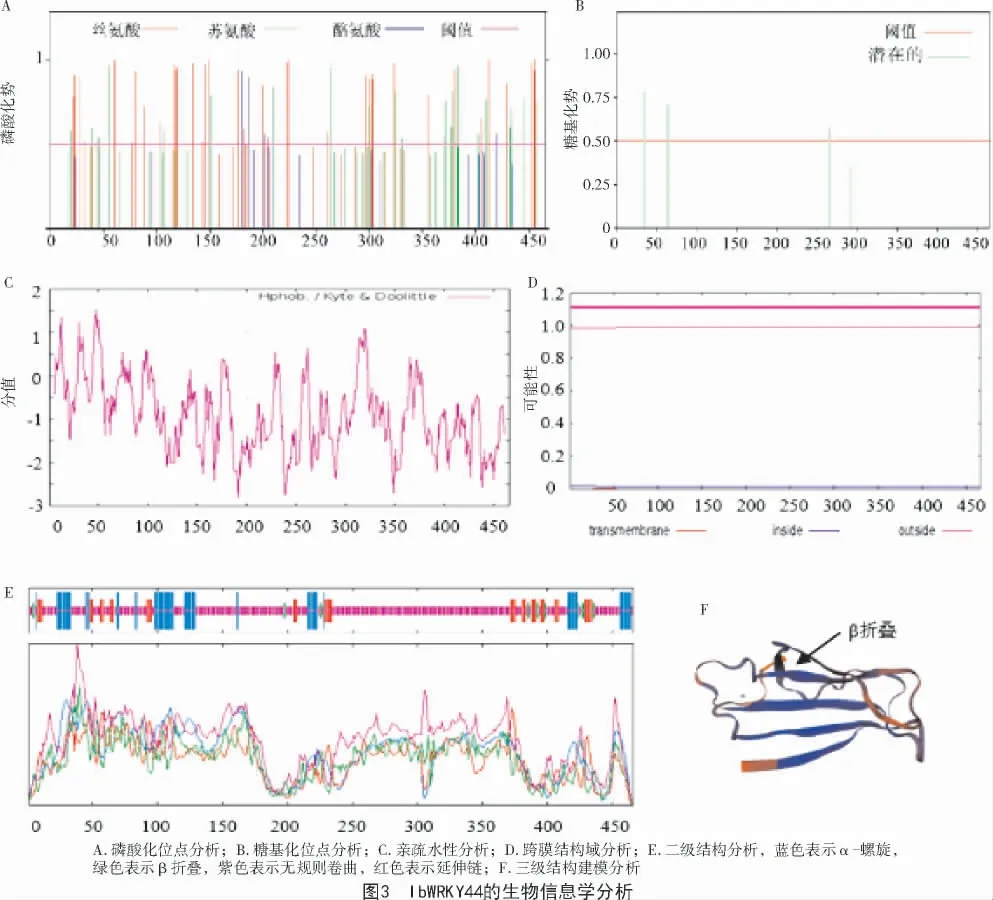
SMART 网站对保守结构域的分析显示,Ib-WRKY44 的氨基酸序列中含有2 个WRKY 保守结构域和2 个C2H2的锌指结构,共466 个氨基酸(图2)。ProtParam 分析显示,IbWRKY44 蛋白的预测分子质量为51 083.08 u,分子式为C2192H3518N654O717S18,总原子数为7099,等电点为9.31,脂溶性指数为53.99,不稳定系数为47.63,表明IbWRKY44 蛋白为碱性不稳定蛋白。NetPhos 分析显示,IbWRKY44存在67 个磷酸化修饰位点,其中,丝氨酸磷酸化位点有40 个,苏氨酸磷酸化位点有23 个,酪氨酸磷酸化位点有4 个(图3-A)。NetNGlyc 分析表明,IbWRKY44 在第36、64、266 个氨基酸位点的天冬酰胺可能存在3 个糖基化位点(图3-B)。ProtScale分析结果表明,IbWRKY44蛋白的氨基酸分值介于-2.822~1.511 之间,且疏水区域的面积明显大于亲水区域,表明IbWRKY44蛋白为疏水性蛋白(图3-C)。信号肽分析结果显示,IbWRKY44蛋白没有信号肽。TMHMM 分析结果表明,IbWRKY44 蛋白没有跨膜结构域(图3-D)。Wolf Psort Prediction 结果显示,IbWRKY44定位于细胞核内。二级结构预测显示,IbWRKY44蛋白主要由α-螺旋(15.24%)、β折叠(3.65%)、无规则卷曲(70.39%)和延伸链(10.73%)组成(图3-E)。三级结构预测发现,该蛋白具有典型的WRKY 转录因子的4β 折叠结构(图3-F)。

2.3 IbWRKY44 的系统进化分析与多序列比对



采用邻接法构建IbWRKY44 与69 个拟南芥WRKYs 蛋白的进化树,结果显示(图4),Ib-WRKY44 与拟南芥AtWRKY44 聚为一支。利用DNAMAN 软件对IbWRKY44 与12 个物种的WRKY44 氨基酸序列进行多序列比对,结果显示,序列相似度为61.38%,均含有2 个WRKY 结构域,且锌指结构为C2H2(图5);物种间系统进化关系显示,IbWRKY44 与同为旋花科的甘薯二倍体ItWRKY44 和牵牛InWRKY44 亲缘关系最近,与葡萄VvWRKY44 亲缘关系最远(图6)。
2.4 IbWRKY44 启动子的顺式作用元件分析
通过在线软件PlantCARE 分析IbWRKY44 上游1 600 bp 启动子序列的顺式作用元件,结果显示,序列中除含有TATA-box 和CAAT-box 等启动子基础元件外,还含有G-Box、GT1-motif、MRE 等光响应元件,脱落酸响应元件ABRE,乙烯响应元件ERE 等元件,以及MYC、MYB、AP1 等转录因子的结合位点(表2)。

表2 IbWRKY44 启动子的顺式作用元件分析
2.5 IbWRKY44 的表达特性分析
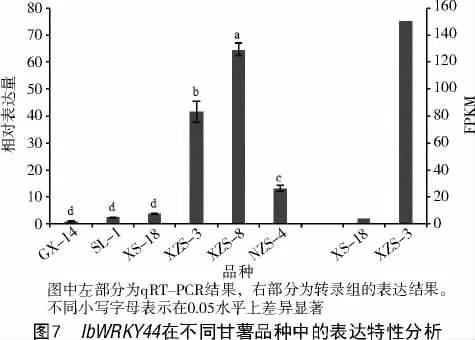
IbWRKY44 在3 个紫心甘薯XZS-3、XZS-8 和NZS-4 和3 个非紫心甘薯XS-18、SL-1 和GX-14的块根中的表达量有显著差异。从图7 可以看出,IbWRKY44 在不同甘薯品种中均表达,但在紫心甘薯中的表达量显著高于非紫心甘薯,与转录组数据一致。
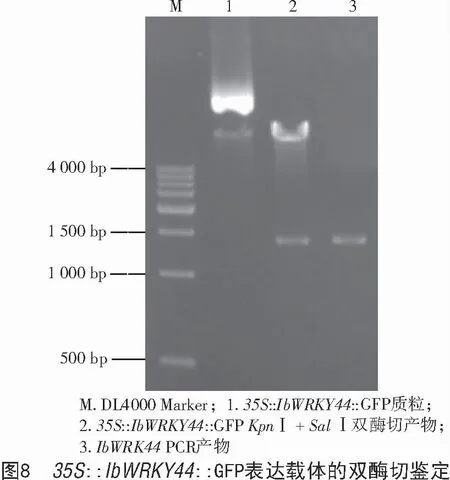
2.6 IbWRKY44 表达载体的构建
以PMD19-T-IbWRKY44 质粒DNA 为模板的扩增产物和pCAMBIA1300-GFP 表达载体经KpnⅠ和SalⅠ双酶切,连接后转至大肠杆菌DH5α,提取阳性克隆的质粒DNA,用KpnⅠ和SalⅠ对质粒进行双酶切验证(图8),酶切获得2 条预期条带,一条为pCAMBIA1300-GFP 载体片段,另一条目的条带与IbWRKY44 基因的PCR 产物大小一致,表明Ib-WRKY44 序列已连接到pCAMBIA1300-GFP 过表达载体,命名为35S::IbWRKY44::GFP 融合表达载体。
2.7 IbWRKY44 的亚细胞定位
亚细胞定位的预测结果显示,IbWRKY44 基因位于细胞核。为验证这一结果,本研究注射烟草叶片进行瞬时表达,观察GFP 荧光部位。将DAPI 染色后的烟草叶片撕取表皮细胞,在488 nm 波长下观察绿色荧光。由图9 可知,转化空载(pCAMBIA1300-GFP)的叶片细胞在细胞质、细胞质膜及细胞核中都存在绿色荧光,而转化融合表达载体(35S::IbWRKY44::GFP)的叶片表皮细胞只在细胞核中有绿色荧光,表明IbWRKY44 蛋白定位于细胞核内,与预测结果一致。

3 结论与讨论
WRKY 转录因子参与植物花青素及其他次生代谢物的合成,大麦HvWRKY23 可以与花青素合成通路基因HvPAL2、HvCHS1 的启动子结合进而促进花青素的合成[22]。YU 等[23]研究发现,丹参中SmWRKY1、SmWRKY7、SmWRKY19、SmWRKY29、SmWRKY45、SmWRKY52、SmWRKY56、SmWRKY58和SmWRKY68 可能参与酮和酚酸的合成。同时,WRKY 转录因子在胁迫响应方面也发挥着重要作用,WEI 等[24]研究发现,GmWRKY54 通过激活转基因大豆ABA 和Ca2+信号通路的基因从而提高植物的抗旱性。张鹏飞[25]在棉花中发现,枯萎病原菌和SA、ETH、JA 等激素的诱导能够激活GhWRKY75基因的表达,进而调控植物对枯萎病的抗性。赵曾强等[26]研究发现,过表达GhWRKY44 的烟草在枯萎病菌的侵染下能够激活抗病相关基因的表达。此外,大量的研究还表明,WRKY 转录因子参与调控植物的生长发育和衰老进程。ZHENG 等[27]在苹果中发现,MdWRKY9 通过抑制油菜素类固醇MdDWF4的转录,从而减少矮化调控因子油菜素类固醇的产生。拟南芥AtWRKY75 通过抑制下游基因的表达来调控根毛的发育[28]。
本试验从XZS-3 的块根中克隆到IbWRKY44,其ORF 序列长度为1 401 bp,编码的蛋白属于GroupⅠ家族,锌指结构为C2H2,含有2 个WRKY结构域。IbWRKY44序列中含有67 个磷酸化位点和3 个糖基化位点,表明IbWRKY44 蛋白可能在蛋白激酶或糖基化作用的调控下,在信号转导、细胞周期等方面发挥作用。本试验预测的IbWRKY44蛋白的三级结构中包括4 个β 折叠,与WRKY 转录因子的空间结构相符。IbWRKY44的烟草亚细胞定位表明,该基因编码的蛋白位于细胞核,在细胞核内发挥功能。IbWRKY44与拟南芥WRKY 家族的AtWRKY44 亲缘关系最近,在物种间的进化关系上与同属旋花科的甘薯二倍体三叶裂薯ItWRKY44和牵牛InWRKY44 的亲缘关系最近,表明IbWRKY44可能具有与AtWRKY44、ItWRKY44 和InWRKY44相似的功能。多序列比对结果表明,IbWRKY44与其他物种的WRKY44 均含有2 个WRKY 结构域,属于Ⅰ类转录因子。
IbWRKY44 的启动子序列中包含脱落酸、乙烯、厌氧和光响应诱导元件,表明脱落酸、乙烯、厌氧和光诱导响应机制中的相关因子可被IbWRKY44 的启动子激活或抑制转录调控过程,从而在植物的生长发育或在对环境的适应机制中发挥作用;包含调节胚乳发育的元件,推测IbWRKY44 参与调控植物胚乳的发育,且调控机制可能与拟南芥AtWRKY44 类似;还含有AP1、MYB 和MYC 转录因子的结合位点,说明IbWRKY44 还可能受AP1、MYB 和MYC 转录因子的调控。
基因表达特性分析结果显示,IbWRKY44 在紫心甘薯块根中的表达量显著高于非紫心甘薯,表明IbWRKY44 可能正调控甘薯块根中花青素的合成。该结果为解析甘薯IbWRKY44 的功能提供了理论基础,丰富了甘薯花青素合成的潜在调节网络,同时为紫心甘薯的分子育种提供了新的研究思路。为了深入研究IbWRKY44 在花青素合成中的功能及挖掘其他功能,后期将对构建好的35S::IbWRKY44::GFP 过表达载体进行拟南芥的转化,筛选得到T3纯合且IbWRKY44 高表达的植株,并测定转基因植株中花青素的含量与花青素合成相关基因的表达量;构建CRISPR-IbWRKY44 敲除表达载体,将其与35S::IbWRKY44::GFP 过表达载体分别进行甘薯悬浮细胞的转化,对得到的转基因甘薯苗进行基因层面的鉴定及表型观察,移至试验田中获得薯块,对薯块进行表型观察及测序分析。同时,筛选与IbWRKY44互作的蛋白,研究其作用机制。
猜你喜欢
农民致富之友(2019年23期)2019-08-16
热带作物学报(2018年12期)2018-05-14
食品界(2017年12期)2018-01-20
现代家庭·生活版(2017年10期)2017-10-20
课堂内外(小学版)(2017年1期)2017-03-08
世界热带农业信息(2016年3期)2016-04-05
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
恋爱婚姻家庭·养生版(2011年8期)2011-05-14
小学生导刊(中年级)(2009年6期)2009-11-10
